欢迎访问作物学报,今天是
作物学报 ›› 2019, Vol. 45 ›› Issue (10): 1586-1594.doi: 10.3724/SP.J.1006.2019.84177
薛晓梦1,李建国1,白冬梅2,晏立英1,万丽云1,康彦平1,淮东欣1,*( ),雷永1,廖伯寿1,*()
),雷永1,廖伯寿1,*()
XUE Xiao-Meng1,LI Jian-Guo1,BAI Dong-Mei2,YAN Li-Ying1,WAN Li-Yun1,KANG Yan-Ping1,HUAI Dong-Xin1,*(),LEI Yong1,LIAO Bo-Shou1,*()
摘要:
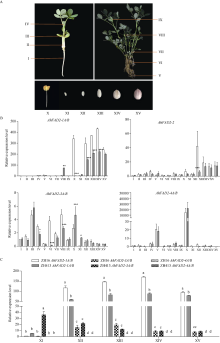
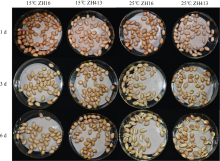
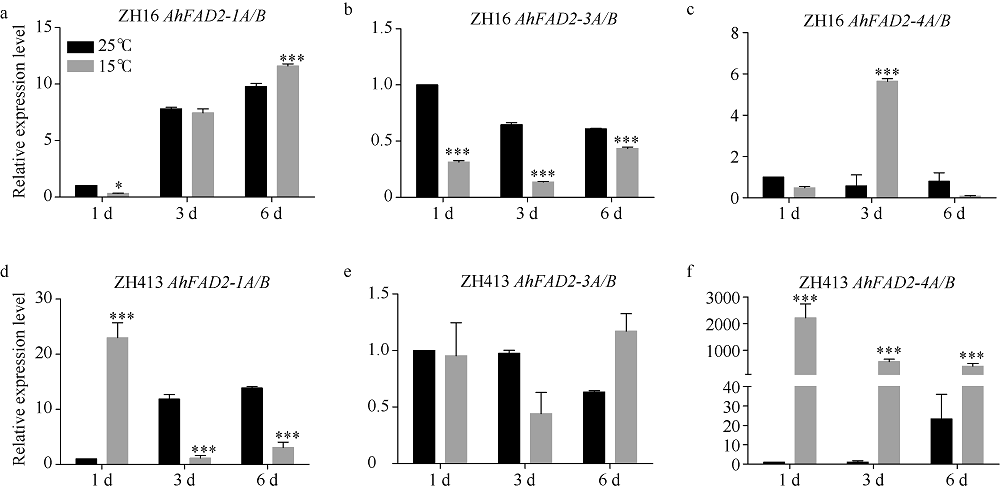
为了探究FAD2在花生低温响应中的作用, 本研究从普通油酸花生中花16 (ZH16)和高油酸花生中花413 (ZH413)中克隆得到花生AhFAD2家族的全部基因, 共7个。通过分析这些基因的表达模式发现, 在ZH16和ZH413中各FAD2基因表达模式相似, AhFAD2-1A/B主要在花和发育的种子中表达, AhFAD2-3A/B主要在营养组织中表达, AhFAD2-4A/B主要在根和花中表达, 表明AhFAD2基因在花生不同发育阶段和不同组织中发挥各自的生物学功能。在15℃下发芽6 d发现, ZH413的发芽率未显著下降, 而ZH16的发芽率显著下降。种子萌发过程中, AhFAD2-1A/B和AhFAD2-4A/B均受低温诱导表达。在ZH16中AhFAD2-1A/B在低温诱导第6天开始显著上调表达, 而在ZH413中第1天显著上调表达; 在ZH16中AhFAD2-4A/B在低温诱导第3天出现显著上调表达, 之后表达量下降, 但在ZH413中第1天就显著上调表达, 且始终维持在高水平表达。基于以上研究结果推测, 高油酸花生在受到低温胁迫时, AhFAD2-1A/B编码蛋白失活, 但AhFAD2-4A/B的高量表达在一定程度上弥补了这部分功能。同时也说明AhFAD2-1A/B功能的缺失并不是决定花生耐寒性的主要因素。本研究的开展为培育抗寒的高油酸花生品种奠定了理论基础, 为高油酸花生在高纬度、高海拔地区推广提供了理论支持。
| [1] | Chi X Y, Zhang Z M, Chen N, Zhang X W, Chen M G, Wang T, Pan L J, Chen J, Yang Z, Guan X Y, Yu S L . Isolation and characterization of fatty acid desaturase genes from peanut ( Arachis hypogaea L.). Plant Cell Rep, 2011,30:1393-1404. |
| [2] | 门爱军, 庞兴国, 胡东青, 陆福军, 李少骞 . 中国花生出口面临的困境及应对措施安徽农业科学, 2016,44:257-258. |
| Men A J, Pang X G, Hu D Q, Lu F J, Li S Q . Difficulties and countermeasures for China’s peanut export. J Anhui Agric Sci, 2016,44:257-258 (in Chinese without English abstract). | |
| [3] | 刘娟, 汤丰收, 张俊, 臧秀旺, 董文召, 易明林, 郝西 . 国内花生生产技术现状及发展趋势研究. 中国农学通报, 2017,33(22):13-18. |
| Liu J, Tang F S, Zhang J, Zang X W, Dong W Z, Yi M L, Hao X . Current status and development trends of peanut production technology in China. Chin Agric Sci Bull, 2017,33(22):13-18 (in Chinese with English abstract). | |
| [4] | Moore K M, Knauft D A . The inheritance of high oleic acid in peanut. Heredity, 1989,80:8-10. |
| [5] | 王传堂, 张建成, 唐月异, 于树涛, 王强, 刘峰, 李秋 . 中国高油酸花生育种现状与展望. 山东农业科学, 2018,50(6):171-176. |
| Wang C T, Zhang J C, Tang Y Y, Yu S T, Wang Q, Liu F, Li Q . Current situation and future directions of high oleic peanut breeding in China. Shandong Agric Sci, 2018,50(6):171-176 (in Chinese with English abstract). | |
| [6] | 迟晓元, 郝翠翠, 陈明娜, 潘丽娟, 陈娜, 王通, 王冕, 杨珍, 梁成伟, 禹山林 . 花生AhFAD2-1基因与油酸/亚油酸比值的关系. 花生学报, 2016,45(4):20-24. |
| Chi X Y, Hao C C, Chen M N, Pan L J, Chen N, Wang T, Wang M, Yang Z, Liang C W, Yu S L . Correlation between AhFAD2-1 and oleic acid/linoleic acid ratio in different peanut varieties. J Peanut Sci, 2016,45(4):20-24 (in Chinese with English abstract). | |
| [7] | Matos A R, Hourton-Cabassa C, Cicek D, Arrabaca J D, Zachowski A, Moreau F . Alternative oxidase involvement in cold stress response of Arabidopsis thaliana fad2 and fad3+ cell suspensions altered in membrane lipid composition. Plant Cell Physiol, 2007,48:856-865. |
| [8] | Miquel M, James D, Dooner H, Browse A J . Arabidopsis requires polyunsaturated lipids for low-temperature survival. Proc Natl Acad Sci USA, 1993,90:6208-6212. |
| [9] | Watanabe K, Oura T, Sakai H, Kajiwara S . Yeast Δ12 fatty acid desaturase: gene cloning, expression, and function. Biosci Biotechnol Biochem, 2004,68:721-727. |
| [10] | Kargiotidou A, Deli D, Galanopoulou D, Tsaftaris A, Farmaki T . Low temperature and light regulate delta 12 fatty acid desaturases (FAD2) at a transcriptional level in cotton ( Gossypium hirsutum). J Exp Bot, 2008,59:2043-2056. |
| [11] | 阮建, 单雷, 李新国, 郭峰, 孟静静, 万书波, 彭振英 . 花生FAD基因家族的全基因组鉴定与表达模式分析. 山东农业科学, 2018,50(6):1-9. |
| Ruan J, Shan L, Li X G, Guo F, Meng J J, Wan S B, Peng Z Y . Genome-wide identification and expression pattern analysis of peanut FAD gene family. Shandong Agric Sci, 2018,50(6):1-9 (in Chinese with English abstract). | |
| [12] | Singh S C, Sinha R P, HaÈder D P . Role of lipids and fatty acids in stress tolerance in cyanobacteria. Acta Protozoo, 2002,41:297-308. |
| [13] | 张建军, 闫世江, 王浩, 司龙亭, 马志国, 杨佳明 . 黄瓜种子脂肪酸含量与耐低温性关系的研究. 安徽农业科学, 2008,36:4859-4861. |
| Zhang J J, Yan S J, Wang H, Si L T, Ma Z G, Yang J M . Study on the relationship between the contents of fatty acids and low temperature tolerance in cucumber seeds. J Anhui Agric Sci, 2008,36:4859-4861 (in Chinese with English abstract). | |
| [14] | Ishizaki-nishizawa O, Fujii T, Ohtani T, Toguri T . Low- temperature resistance of higher plants is significantly enhanced by a nonspecific cyanobacterial desaturase. Nat Biotechnol, 1996,14:1003-1006. |
| [15] | 黄锐之, 刘智宏, 郎春秀, 胡张华, 陈锦清 . 植物中多不饱和脂肪酸生物合成的基因工程. 植物生理学通讯, 2001,37:547-550. |
| Huang R Z, Liu Z H, Lang C X, Hu Z H, Chen J Q . Genetic engineering of polyunsaturated fatty acid biosynthesis in plants. Plant Physiol Commun, 2001,37:547-550 (in Chinese with English abstract). | |
| [16] | 陈儒钢, 巩振辉, 逯明辉, 李大伟, 黄炜 . 植物抗寒基因工程研究进展. 西北植物学报, 2008,28:1274-1280. |
| Chen R G, Gong Z H, Lu M H, Li D W, Huang W . Research advance in the cold-resistance genetic engineering in plants. Acta Bot Boreali-Occident Sin, 2008,28:1274-1280 (in Chinese with English abstract). | |
| [17] | 王洪春 . 植物抗逆性与生物膜结构功能研究的进展. 植物生理学通讯, 1985, ( 1):60-66. |
| Wang H C . Advances in studies on plant stress resistance and biofilm structure. Plant Physiol Commun, 1985, ( 1):60-66 (in Chinese with English abstract). | |
| [18] | 杨玉珍, 罗青, 彭方仁 . 不同种源香椿叶片脂肪酸组分含量与抗寒性关系. 林业科技开发, 2011,25(5):21-25. |
| Yang Y Z, Luo Q, Peng F R . A relationship between cold resistance and content of fatty acid composition in leaves of Toona sinensis from different provenances. China For Sci Technol, 2011,25(5):22-25 (in Chinese with English abstract). | |
| [19] | 戴晓峰, 肖玲, 武玉花, 吴刚, 卢长明 . 植物脂肪酸去饱和酶及其编码基因研究进展. 植物学通报, 2007,24:105-113. |
| Dai X F, Xiao L, Wu Y H, Wu G, Lu C M . An overview of plant fatty acid desaturases and the coding genes. Chin Bull Bot, 2007,24:105-113 (in Chinese with English abstract). | |
| [20] | Yang Q, Fan C, Guo Z, Qin J, Wu J, Li Q, Fu T, Zhou Y . Identification of FAD2 and FAD3 genes in Brassica napus genome and development of allele-specific markers for high oleic and low linolenic acid contents. Theor Appl Genet, 2012,125:715-729. |
| [21] | Cao S J, Zhou X R, Wood C C, Green G A, Singh S P, Liu L X, Liu Q . A large and functionally diverse family of Fad2 genes in safflower( Carthamus tinctorius L.). BMC Plant Biol, 2013,13:5. doi: 10.1186/1471-2229-13-5. |
| [22] | 张照华, 王志慧, 淮东欣, 谭家壮, 陈剑洪, 晏立英, 王晓军, 万丽云, 陈傲, 康彦平, 姜慧芳, 雷永, 廖伯寿 . 利用回交和标记辅助选择快速培育高油酸花生品种及其评价. 中国农业科学, 2018,51:1641-1652. |
| Zhang Z H, Wang Z H, Huai D X, Tan J Z, Chen J H, Yan L Y, Wang X J, Wan L Y, Chen A, Kang Y P, Jiang H F, Lei Y, Liao B S . Fast development of high oleate peanut cultivars by using maker-assisted backcrossing and their evaluation. Sci Agric Sin, 2018,51:1641-1652 (in Chinese with English abstract). | |
| [23] | Jung S, Swift D, Sengoku E, Patel M, Teule F, Powell G, Moore K, Abbott A . The high oleate trait in the cultivated peanut ( Arachis hypogaea L.). Isolation and characterization of two genes encoding microsomal oleoyl-PC desaturases. Mol Gen Genet, 2000,263:796-805. |
| [24] | Wang Y, Zhang X G, Zhao Y L, Prakash C S, He G H, Yin D M. Insights into the novel members of the FAD2 gene family involved in high-oleate fluxes in peanut. NRC Res Press, 2015,58:1-9. |
| 周丽侠, 唐桂英, 陈高, 毕玉平, 单雷 . 花生AhFAD2基因的多态性及其与籽粒油酸/亚油酸比值间的相关性. 作物学报, 2011,37:415-423. | |
| Zhou L Y, Tang G Y, Chen G, Bi Y P, Shan L . Correlation between AhFAD2 polymorphism and oleic acid/linoleic acid ratio in peanut seeds. Acta Agron Sin, 2011,37:415-423 (in Chinese with English abstract). | |
| [25] | D’Angeli S, Falasca G, Matteucci M M, Altamura M M . Cold perception and gene expression differ in Olea europaea seed coat and embryo during drupe cold acclimation. New Phytol, 2013,197:123-138. |
| [26] | 唐桂英, 王芳, 徐平丽, 单雷 . 花生AhFAD2-2基因的克隆与表达分析. 山东农业科学, 2018,50(6):27-34. |
| Tang G Y, Wang F, Xu P L, Shan L . Cloning and expression analysis of AhFAD2-2 gene in peanut. Shandong Agric Sci, 2018,50(6):27-34 (in Chinese with English abstract). | |
| [27] | Heppard E P, Kinney A J, Stecca K L, Miao G H . Developmental and growth temperature regulation of two different microsomal ω-6 desaturase genes in soybeans. Plant Physiol, 1996,110:311-319. |
| [28] | Jung S, Powell G, Moore K, Abbott A . The high oleate trait in the cultivated peanut ( Arachis hypogaea L.): II. Molecular basis and genetics of the trait. Mol Gen Genet, 2000,263:806-811. |
| [29] | Okuley J, Lightner J, Feldmann K, Yadav N, Lark E, Browse J . Arabidopsis FAD2 gene encodes the enzyme that is essential for polyunsaturated lipid synthesis. Plant Cell, 1994,6:147-158. |
| [30] | 李金金, 张晶晶, 年洪娟 . Δ 12-脂肪酸去饱和酶FAD2的基本特性及其在胁迫中的功能 . 生命科学研究, 2013,17:174-178. |
| Li J J, Zhang J J, Nian H J . Characteristics of Δ 12-fatty acid desaturase FAD2 and its functions under stress. Life Sci Res, 2013,17:174-178 (in Chinese with English abstract). | |
| [31] | 年洪娟, 陈丽梅 . 不饱和脂肪酸在逆境胁迫中的作用. 中国微生态学杂志, 2012,24:760-762. |
| Nian H J, Chen L M . The role of unsaturated fatty acid in various environmental stresses. Chin J Microecol, 2012,24:760-762 (in Chinese with English abstract). | |
| [32] | Orvar B L, Sangwan V, Omann F, Dhindsa R S . Early steps in cold sensing by plant cells: the role of actin cytoskeleton and membrane fluidity. Plant J, 2000,23:785-794. |
| [1] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [2] | 李海芬, 魏浩, 温世杰, 鲁清, 刘浩, 李少雄, 洪彦彬, 陈小平, 梁炫强. 花生电压依赖性阴离子通道基因(AhVDAC)的克隆及在果针向地性反应中表达分析[J]. 作物学报, 2022, 48(6): 1558-1565. |
| [3] | 姚晓华, 王越, 姚有华, 安立昆, 王燕, 吴昆仑. 青稞新基因HvMEL1 AGO的克隆和条纹病胁迫下的表达[J]. 作物学报, 2022, 48(5): 1181-1190. |
| [4] | 周慧文, 丘立杭, 黄杏, 李强, 陈荣发, 范业赓, 罗含敏, 闫海锋, 翁梦苓, 周忠凤, 吴建明. 甘蔗赤霉素氧化酶基因ScGA20ox1的克隆及功能分析[J]. 作物学报, 2022, 48(4): 1017-1026. |
| [5] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [6] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
| [7] | 陈新宜, 宋宇航, 张孟寒, 李小艳, 李华, 汪月霞, 齐学礼. 干旱对不同品种小麦幼苗的生理生化胁迫以及外源5-氨基乙酰丙酸的缓解作用[J]. 作物学报, 2022, 48(2): 478-487. |
| [8] | 谢琴琴, 左同鸿, 胡燈科, 刘倩莹, 张以忠, 张贺翠, 曾文艺, 袁崇墨, 朱利泉. 甘蓝自交不亲和相关基因BoPUB9的克隆及表达分析[J]. 作物学报, 2022, 48(1): 108-120. |
| [9] | 薛晓梦, 吴洁, 王欣, 白冬梅, 胡美玲, 晏立英, 陈玉宁, 康彦平, 王志慧, 淮东欣, 雷永, 廖伯寿. 低温胁迫对普通和高油酸花生种子萌发的影响[J]. 作物学报, 2021, 47(9): 1768-1778. |
| [10] | 王艳朋, 凌磊, 张文睿, 王丹, 郭长虹. 小麦B-box基因家族全基因组鉴定与表达分析[J]. 作物学报, 2021, 47(8): 1437-1449. |
| [11] | 宋天晓, 刘意, 饶莉萍, Soviguidi Deka Reine Judesse, 朱国鹏, 杨新笋. 甘薯细胞壁蔗糖转化酶基因IbCWIN家族成员鉴定及表达分析[J]. 作物学报, 2021, 47(7): 1297-1308. |
| [12] | 唐锐敏, 贾小云, 朱文娇, 印敬明, 杨清. 马铃薯热激转录因子HsfA3基因的克隆及其耐热性功能分析[J]. 作物学报, 2021, 47(4): 672-683. |
| [13] | 岳洁茹, 白建芳, 张风廷, 郭丽萍, 苑少华, 李艳梅, 张胜全, 赵昌平, 张立平. 杂交小麦抗坏血酸过氧化物酶基因克隆及其在种子老化中的潜在功能分析[J]. 作物学报, 2021, 47(3): 405-415. |
| [14] | 解盼, 刘蔚, 康郁, 华玮, 钱论文, 官春云, 何昕. 甘蓝型油菜CBF基因家族的鉴定和表达分析[J]. 作物学报, 2021, 47(12): 2394-2406. |
| [15] | 李鹏, 刘彻, 宋皓, 姚盼盼, 苏沛霖, 魏跃伟, 杨永霞, 李青常. 烟草非特异性脂质转移蛋白基因家族的鉴定与分析[J]. 作物学报, 2021, 47(11): 2184-2198. |
|
||