欢迎访问作物学报,今天是
作物学报 ›› 2019, Vol. 45 ›› Issue (11): 1649-1655.doi: 10.3724/SP.J.1006.2019.93009
王晓娟1,潘振远2,刘敏2,刘忠祥1,周玉乾1,何海军1,邱法展2,*( )
)
WANG Xiao-Juan1,PAN Zhen-Yuan2,LIU Min2,LIU Zhong-Xiang1,ZHOU Yu-Qian1,HE Hai-Jun1,QIU Fa-Zhan2,*()
摘要:
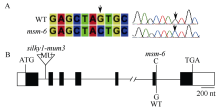
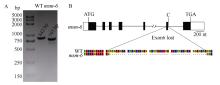
前期试验发现一个玉米雄性不育突变体(male sterile mutant), 命名为msm-6。经过连续多代杂合单株自交发现该突变体性状能稳定遗传, 遗传模式分析表明该突变体为单个隐性 1核基因控制。以msm-6 与自交系B73 杂交构建F2 遗传定位群体, 利用BSA (Bulk Segregant Analysis)方法, 筛选到与msm-6位点连锁的4个SSR标记, 即C6-24、C6-30、C6-34和C6-40。进一步采用444个F2单株对这4个连锁标记验证, 将msm-6定位于标记C6-24与C6-34之间, 即玉米第6染色体68.5~98.1 Mb之间。通过基因组序列信息分析发现, 在此定位区间内存在一个已报道的Silky1基因。Silky1基因编码MADS-BOX蛋白, 是参与花器官建成ABCD模型的B功能基因, 该基因的突变会造成雄花不育、雌花花丝增多等表型。利用杂合体+/silky1-mum3与纯合突变体msm-6/msm-6杂交进行等位测验, 杂交后代中正常植株与突变植株分离比例符合1∶1。基因组序列以及转录本序列分析发现, msm-6突变体中第6个内含子5'端供体位点AG/GTAAG (外显子/内含子交接处)的序列+1位G突变为C, 导致第6外显子被错误剪切掉, 产生了第6个外显子缺失的Silky1异常转录本。以上实验证据表明, msm-6与所报道的由mutator转座子插入造成的Silky1突变体silky1-mum2、silky1-mum3和silky1-mum4的突变方式不同, 是一个新的Silky1基因等位突变体。msm-6的发现与鉴定为进一步深入解析玉米穗花器官决定的遗传机制提供丰富的实验材料, 同时也为RNA加工过程中剪接位点保守性提供重要证据。
| [1] | Coen E S, Meyerowitz E M . The war of the whorls: genetic interactions controlling flower development. Nature, 1991,353:31-37. |
| [2] | Pelaz S, Ditta G S, Baumann E, Wisman E, Yanofsky M F . B and C floral organ identity functions require SEPALLATA MADS-box genes. Nature, 2000,405:200-203. |
| [3] | Bowman J L, Smyth D R, Meyerowitz E M . Genetic interactions among floral homeotic genes of Arabidopsis. Development, 1991,112:1-20. |
| [4] | Honma T, Goto K . Complexes of MADS-box proteins are sufficient to convert leaves into floral organs. Nature, 2001 409:525-529. |
| [5] | Schmidt R J, Ambrose B A . The blooming of grass flower development. Curr Opin Plant Biol, 1998,1:60-67. |
| [6] | Ambrose B A, Lerner D R, Ciceri P, Padilla C M, Yanofsky M F, Schmidt R J . Molecular and genetic analyses of the silky1 gene reveal conservation in floral organ specification between eudicots and monocots. Mol Cell, 2000,5:569-579. |
| [7] | Moore M J, Sharp P A . Evidence for two active sites in the spliceosome provided by stereochemistry of pre-mRNA splicing. Nature, 1993,365:364-368. |
| [8] | Simpson G F W . Splicing of precursors to mRNA in higher plants: Mechanism, regulation and sub-nuclear organisation of the spliceosomal machinery. Plant Mol Biol, 1996,32:1-41. |
| [9] | Robberson B L, Cote G J, Berget S M . Exon definition may facilitate splice site selection in RNAs with multiple exons. Mol Cell Biol, 1990,10:84. |
| [10] | Collins L, Penny D . Proceedings of the SMBE Tri-National Young Investigators’ Workshop 2005. Investigating the intron recognition mechanism in eukaryotes. Mol Biol Evol, 2006,23:901-910. |
| [11] | Nakai K, Sakamoto H . Construction of a novel database containing aberrant splicing mutations of mammalian genes, Gene, 1994,141:171-177. |
| [12] | Alvarez C J, Wise J A . Activation of a cryptic 5 splice site by U1 snRNA. RNA, 2001,7:342-350. |
| [13] | Carmel I, Tal S, Vig I, Ast G . Comparative analysis detects dependencies among the 5 splice-site positions. RNA, 2004,10:828-840. |
| [14] | Michelmore R W, Paran I, Kesseli R V . Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991,88:9828-9832. |
| [15] | Suzuki M, Settles A M, Tseung C W, Li Q B, Latshaw S, Wu S, Porch T G, Schmelz E A, James M G, McCarty D R . The maize viviparous15 locus encodes the molybdopterin synthase small subunit. Plant J, 2006,45:264-274. |
| [16] | 王关林, 方宏筠 . 植物基因工程(第2版). 北京: 科学出版社, 2002. pp 742-744. |
| Wang G L, Fang H J . Plant Genetic Engineering, 2nd edn. Beijing: Science Press, 2002. pp 742-744(in Chinese) | |
| [17] | Takagi H1, Abe A, Yoshida K, Kosugi S, Natsume S, Mitsuoka C, Uemura A, Utsushi H, Tamiru M, Takuno S, Innan H, Cano L M, Kamoun S, Terauchi R . QTL-seq: rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J, 2013,74:174-183. |
| [18] | Wenger W J, Schwartz K, Sherlock G . Bulk segregant analysis by high-throughput sequencing reveals a novel xylose utilization gene from Saccharomyces cerevisiae. PLoS Genet, 2010,6:e1000942. |
| [19] | Cai M J, Li S Z, Sun F, Sun Q, Zhao H L, Ren X M, Zhao Y X, Tan B C, Zhang Z X, Qiu F Z . Emp10 encodes a mitochondrial PPR protein that affects the cis-splicing of nad2 intron 1 and seed development in maize. Plant J, 2017,91:132-144. |
| [20] | Zhao Y, Zhang Y, Wang L, Wang X, Xu W, Gao X, Liu B . Mapping and functional analysis of a maize silkless mutant sk-A7110. Front Plant Sci, 2018,9:1227. |
| [21] | Strable J, Wallace J G . Maize YABBY genes drooping leaf1 and drooping leaf2 regulate plant architecture. Plant Cell, 2017,29:1622-1641. |
| [22] | Thi Tran H T, Takeshima Y, Surono A, Yagi M, Wada H, Matsuo M . A G- to -A transition at the fifth position of intron-32 of the dystrophin gene inactivates a splice-donor site both in vivo and in vitro. Mol Genet Metab, 2005,85:213-219. |
| [23] | Jap T S, Wu Y C, Tso Y C, Chiu C Y . A novel mutation in the intron 1 splice donor site of the cholesterol ester transfer protein (CETP) gene as a cause of hyperalphalipoproteinemia. Metabolism, 2002,51:394-397. |
| [24] | Hirano H Y, Eiguchi M, Sano Y . A single base change altered the regulation of the Waxy gene at the posttranscriptional level during the domestication of rice. Mol Biol Evol, 1998,15:978-987. |
| [25] | Lal S, Choi J H, Shaw J R, Hannah L C . A splice site mutant of maize activates cryptic splice sites, elicits intron inclusion and exon exclusion, and permits branch point elucidation. Plant Physiol, 1999,121:411-418. |
| [26] | Kramer E M, Dorit, R L, Irish V F . Molecular evolution of genes controlling petal and stamen development: divergence within the APETALA3 and PISTILLATA MADS box gene lineages. Genetics, 1998,149:765-783. |
| [1] | 肖颖妮, 于永涛, 谢利华, 祁喜涛, 李春艳, 文天祥, 李高科, 胡建广. 基于SNP标记揭示中国鲜食玉米品种的遗传多样性[J]. 作物学报, 2022, 48(6): 1301-1311. |
| [2] | 崔连花, 詹为民, 杨陆浩, 王少瓷, 马文奇, 姜良良, 张艳培, 杨建平, 杨青华. 2个玉米ZmCOP1基因的克隆及其转录丰度对不同光质处理的响应[J]. 作物学报, 2022, 48(6): 1312-1324. |
| [3] | 郑崇珂, 周冠华, 牛淑琳, 和亚男, 孙伟, 谢先芝. 水稻早衰突变体esl-H5的表型鉴定与基因定位[J]. 作物学报, 2022, 48(6): 1389-1400. |
| [4] | 王丹, 周宝元, 马玮, 葛均筑, 丁在松, 李从锋, 赵明. 长江中游双季玉米种植模式周年气候资源分配与利用特征[J]. 作物学报, 2022, 48(6): 1437-1450. |
| [5] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [6] | 陈静, 任佰朝, 赵斌, 刘鹏, 张吉旺. 叶面喷施甜菜碱对不同播期夏玉米产量形成及抗氧化能力的调控[J]. 作物学报, 2022, 48(6): 1502-1515. |
| [7] | 徐田军, 张勇, 赵久然, 王荣焕, 吕天放, 刘月娥, 蔡万涛, 刘宏伟, 陈传永, 王元东. 宜机收籽粒玉米品种冠层结构、光合及灌浆脱水特性[J]. 作物学报, 2022, 48(6): 1526-1536. |
| [8] | 单露英, 李俊, 李亮, 张丽, 王颢潜, 高佳琪, 吴刚, 武玉花, 张秀杰. 转基因玉米NK603基体标准物质研制[J]. 作物学报, 2022, 48(5): 1059-1070. |
| [9] | 王好让, 张勇, 于春淼, 董全中, 李微微, 胡凯凤, 张明明, 薛红, 杨梦平, 宋继玲, 王磊, 杨兴勇, 邱丽娟. 大豆突变体ygl2黄绿叶基因的精细定位[J]. 作物学报, 2022, 48(4): 791-800. |
| [10] | 许静, 高景阳, 李程成, 宋云霞, 董朝沛, 王昭, 李云梦, 栾一凡, 陈甲法, 周子键, 吴建宇. 过表达ZmCIPKHT基因增强植物耐热性[J]. 作物学报, 2022, 48(4): 851-859. |
| [11] | 刘磊, 詹为民, 丁武思, 刘通, 崔连花, 姜良良, 张艳培, 杨建平. 玉米矮化突变体gad39的遗传分析与分子鉴定[J]. 作物学报, 2022, 48(4): 886-895. |
| [12] | 闫宇婷, 宋秋来, 闫超, 刘爽, 张宇辉, 田静芬, 邓钰璇, 马春梅. 连作秸秆还田下玉米氮素积累与氮肥替代效应研究[J]. 作物学报, 2022, 48(4): 962-974. |
| [13] | 徐宁坤, 李冰, 陈晓艳, 魏亚康, 刘子龙, 薛永康, 陈洪宇, 王桂凤. 一个新的玉米Bt2基因突变体的遗传分析和分子鉴定[J]. 作物学报, 2022, 48(3): 572-579. |
| [14] | 宋仕勤, 杨清龙, 王丹, 吕艳杰, 徐文华, 魏雯雯, 刘小丹, 姚凡云, 曹玉军, 王永军, 王立春. 东北主推玉米品种种子形态及贮藏物质与萌发期耐冷性的关系[J]. 作物学报, 2022, 48(3): 726-738. |
| [15] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
|
||