欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (7): 1205-1214.doi: 10.3724/SP.J.1006.2021.01074
• 作物遗传育种·种质资源·分子遗传学 • 下一篇
耿腊1, 黄业昌2, 李梦迪1, 谢尚耿1, 叶玲珍1,3,*( ), 张国平1
), 张国平1
GENG La1, HUANG Ye-Chang2, LI Meng-Di1, XIE Shang-Geng1, YE Ling-Zhen1,3,*(), ZHANG Guo-Ping1
摘要:
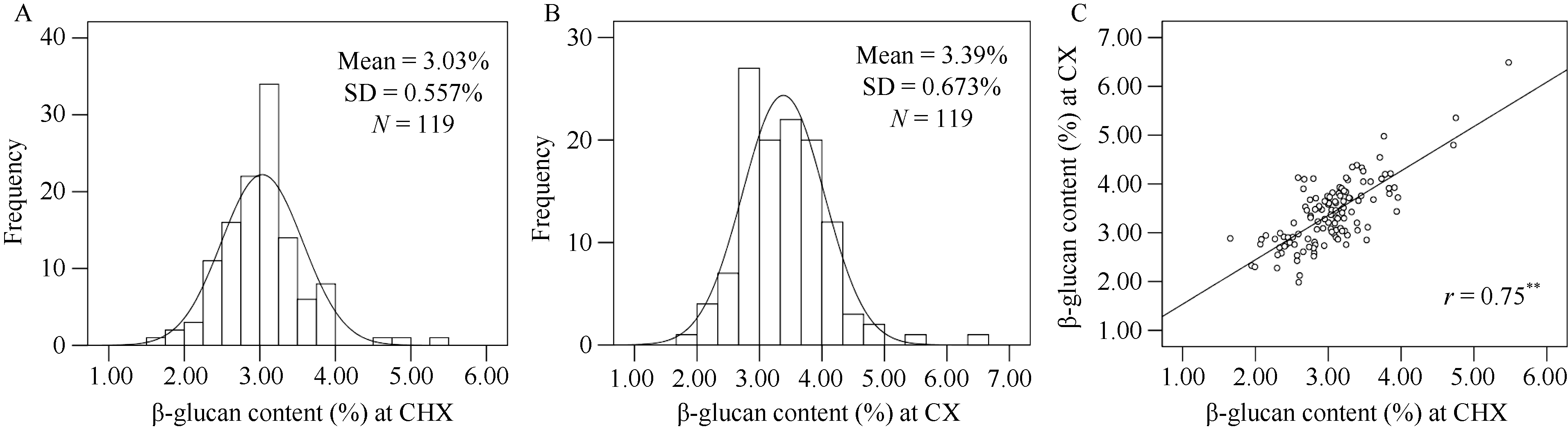
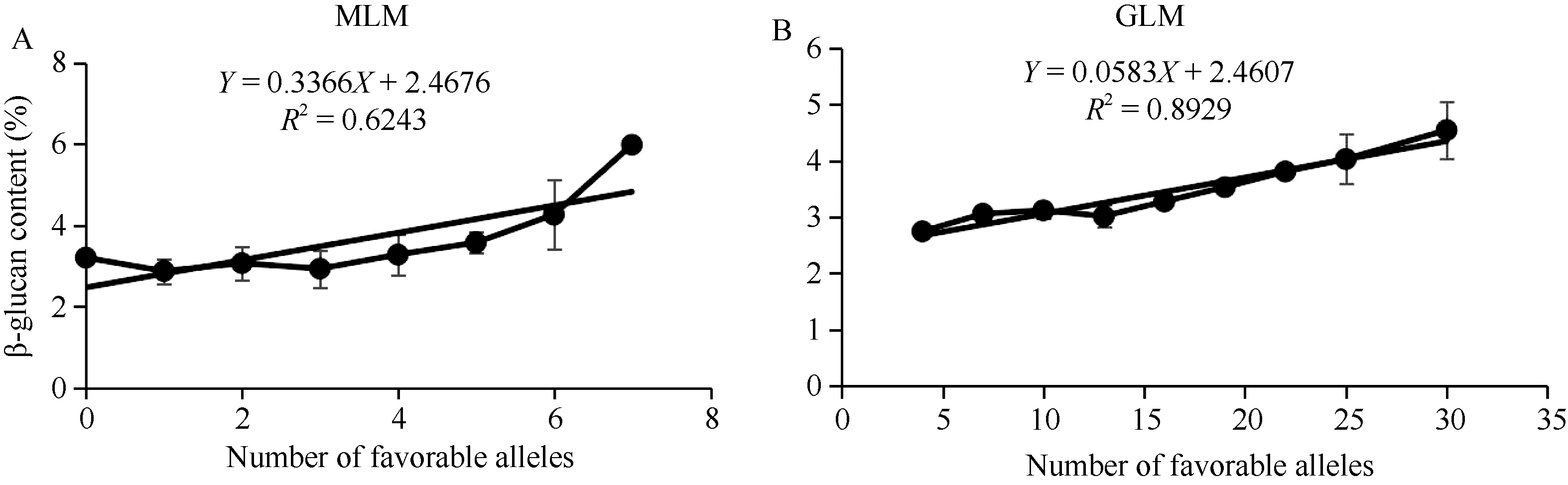
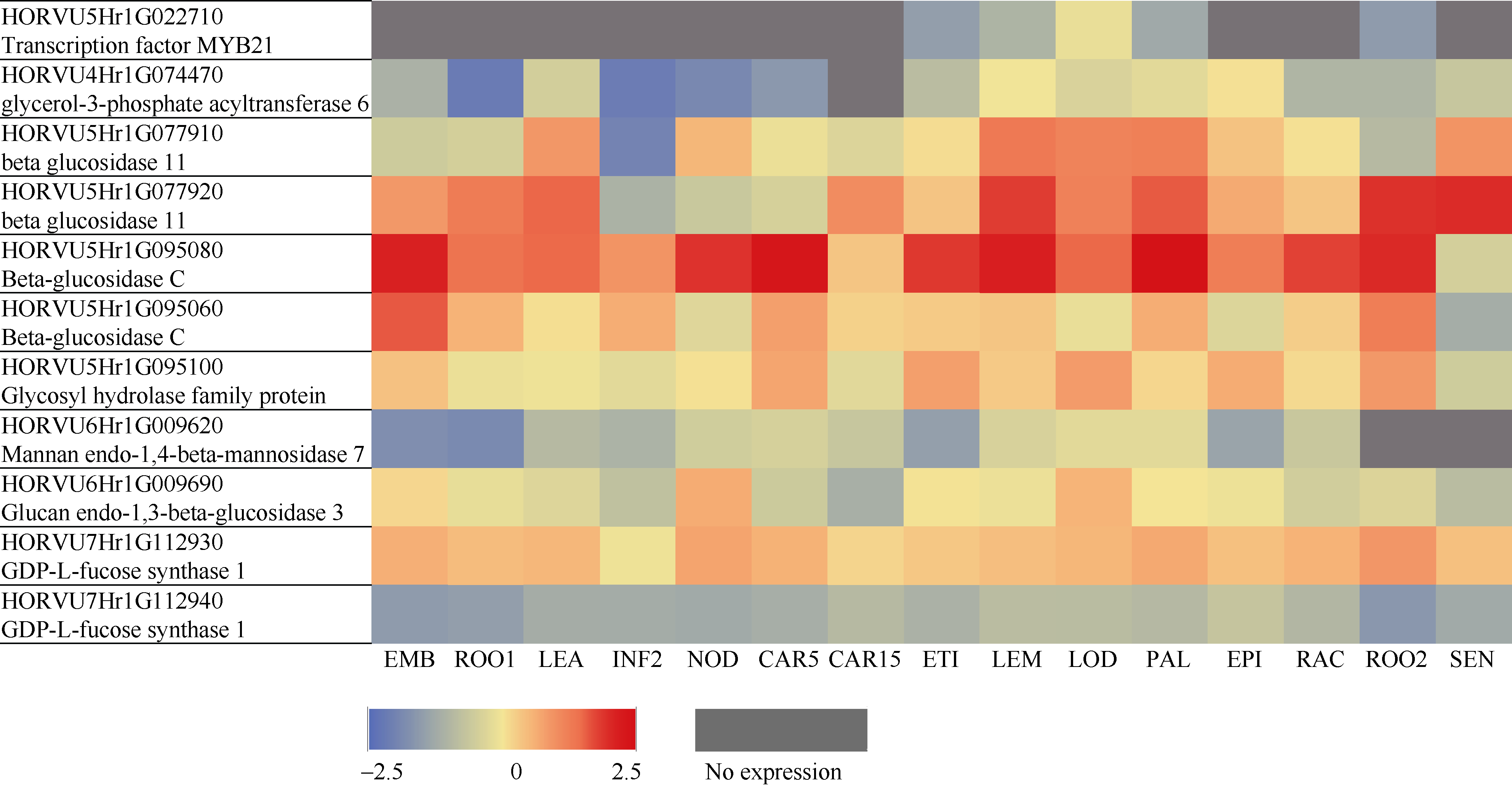
β-葡聚糖是大麦籽粒的一个重要品质性状, 其含量高低影响大麦啤用、饲用和食用品质。虽然有关大麦β-葡聚糖合成的相关基因已有报道, 但关于大麦籽粒β-葡聚糖积累的遗传调控机制仍不十分清楚。本研究以前期收集的全球119份大麦基因型为材料, 种植在土壤与气候条件有一定差异的两试点, 利用混合线性模型(MLM)和一般线性模型(GLM)对不同大麦材料籽粒β-葡聚糖含量进行GWAS分析。结果表明, 大麦籽粒β-葡聚糖含量的基因型差异显著, 其在2个环境下的广义遗传力为73.9%。利用MLM和GLM模型分别检测到8个和40个显著位点, 合并2个模型重叠位点后共得到44个显著位点, 其中HORVU5Hr1G022710基因在2个模型、2个地点均被鉴定到, 故被认为是与β-葡聚糖含量显著相关的候选基因。2个模型中最佳等位基因数与β-葡聚糖含量均呈显著正相关。此外, 基于基因注释, 共鉴定到10个与糖合成、转运及分解相关的酶类基因, 这些基因可能与籽粒中β-葡聚糖的合成、积累和分解紧密相关。本研究结果为阐明β-葡聚糖的遗传调控机制提供了新的视角, 亦为大麦籽粒β-葡聚糖的遗传改良奠定了一定的理论基础。
| [1] | Havrlentová M, Kraic J. Content of β-D-glucan in cereal grains. J Food Nutr Res-Slov, 2006,45:97-103. |
| [2] | Zhang G, Chen J, Wang J, Ding S. Cultivar and environmental effects on (1→3, 1→4) β-D-glucan and protein content in malting barley. J Cereal Sci, 2001,34:295-301. |
| [3] | Jeroch H, Dänicke S. Barley in poultry feeding: a review. Worlds Poultry Ence J, 1995,51:271-291. |
| [4] | Mcnab J M. Barley β-glucan: an antinutritional factor in poultry feeding. Nutr Res Rev, 1992,5:45-60. |
| [5] |
Kerckhoffs D A J M, Hornstra G, Mensink R P. Cholesterol-lower ing effect of beta-glucan from oat bran in mildly hypercholesterolemic subjects may decrease when beta-glucan is incorporated into bread and cookies. Am J Clin Nutr, 2003,78:221-227.
doi: 10.1093/ajcn/78.2.221 pmid: 12885701 |
| [6] |
Pear J R, Kawagoe Y, Schreckengost W E, Delmer D P, Stalker D M. Higher plants contain homologs of the bacterial celA genes encoding the catalytic subunit of cellulose synthase. Proc Natl Acad Sci USA, 1996,93:12637-12642.
doi: 10.1073/pnas.93.22.12637 pmid: 8901635 |
| [7] |
Doblin M S, Pettolmo F A, Wilson S M, Campbell R, Burton R A, Fincher G B, Newbigin E, Bacic A. A barley cellulose synthase-like CSLH gene mediates (1,3;1,4)-β-D-glucan synthesis in transgenicarabidopsis. Proc Natl Acad Sci USA, 2009,106:5996-6001.
doi: 10.1073/pnas.0902019106 pmid: 19321749 |
| [8] |
Burton A R. Cellulose synthase-like cslf genes mediate the synthesis of cell wall (1,3;1,4)-β-D-glucans. Science, 2006,311:1940-1942.
doi: 10.1126/science.1122975 pmid: 16574868 |
| [9] |
Schreiber M, Wright F, Mackenzie K, Hedley P E, Schwerdt J G, Little A, Burton R A, Fincher G B, Marshall D, Waugh R, Halpin C. The barley genome sequence assembly reveals three additional members of the CslF( 1,3;1,4)-beta-glucan synthase gene family. PLoS One, 2014,9:e90888.
pmid: 24595438 |
| [10] |
Burton R A, Collins H M, Kibble N A J, Smith J A, Shirley N J, Jobling S A, Henderson M, Singh R R, Pettolino F, Wilson S M, Bird A R, Topping D L, Bacic A, Fincher G B. Over-expression of specific HvCslF cellulose synthase-like genes in transgenic barley increases the levels of cell wall (1,3;1,4)-β-D-glucans and alters their fine structure. Plant Biotechnol J, 2011,9:117-135.
doi: 10.1111/j.1467-7652.2010.00532.x pmid: 20497371 |
| [11] |
Nemeth C, Freeman J, Jones H D, Sparks C, Pellny T K, Wilkinson M D, Dunwell J, Andersson A A M, åman P, Guillon F, Saulnier L, Mitchell R A C, Shewry P R. Down-regulation of the CSLF6 gene results in decreased (1,3;1,4)-β-D-glucan in endosperm of wheat. Plant Physiol, 2010,152:1209-1218.
doi: 10.1104/pp.109.151712 pmid: 20089768 |
| [12] |
Zhang C, Zhou Z, Yong H, Zhang X, Hao Z, Zhang F, Li M, Zhang D, Li X, Wang Z. Analysis of the genetic architecture of maize ear and grain morphological traits by combined linkage and association mapping. Theor Appl Genet, 2017,130:1011-1029.
doi: 10.1007/s00122-017-2867-7 pmid: 28215025 |
| [13] | Tadesse W, Ogbonnaya F C, Jighly A, Sanchez-Garcia M, Sohail Q, Rajaram S, Baum M. Genome-wide association mapping of yield and grain quality traits in winter wheat genotypes. PLoS One, 2015,10:e141339. |
| [14] | Huang X, Wei X, Sang T, Zhao Q, Feng Q, Zhao Y, Li C, Zhu C, Lu T, Zhang Z. Genome-wide association studies of 14 agronomic traits in rice landraces. Nat Genet, 2010,42:961. |
| [15] |
Houston K, McKim S M, Comadran J, Bonar N, Druka I, Uzrek N, Cirillo E, Guzy-Wrobelska J, Collins N C, Halpin C, Hansson M, Dockter C, Druka A, Waugh R. Variation in the interaction between alleles of HvAPETALA2 and microRNA172 determines the density of grains on the barley inflorescence. Proc Natl Acad Sci USA, 2013,110:16675-16680.
doi: 10.1073/pnas.1311681110 pmid: 24065816 |
| [16] |
Comadran J, Kilian B, Russell J, Ramsay L, Stein N, Ganal M, Shaw P, Bayer M, Thomas W, Marshall D, Hedley P, Tondelli A, Pecchioni N, Francia E, Korzun V, Walther A, Waugh R. Natural variation in a homolog of Antirrhinum CENTRORADIALIS contributed to spring growth habit and environmental adaptation in cultivated barley. Nat Genet, 2012,44:1388-1392.
doi: 10.1038/ng.2447 pmid: 23160098 |
| [17] |
Ramsay L, Comadran J, Druka A, Marshall D F, Waugh R. INTERMEDIUM-C, a modifier of lateral spikelet fertility in barley, is an ortholog of the maize domestication gene TEOSINTE BRANCHED 1. Nat Genet, 2011,43:169-172.
doi: 10.1038/ng.745 pmid: 21217754 |
| [18] |
Houston K, Russell J, Schreiber M, Halpin C, Oakey H, Washington J M, Booth A, Shirley N, Burton R A, Fincher G B. A genome wide association scan for (1,3;1,4)-β-glucan content in the grain of contemporary 2-row Spring and Winter barleys. BMC Genomics, 2014,15:907.
doi: 10.1186/1471-2164-15-907 pmid: 25326272 |
| [19] | Narasimhalu P, Kong D, Choo T M, Ho K M, Ferguson T, Therrien M C, May K W, Jui P. Effects of environment and cultivar on total mixed-linkage β-glucan content in eastern and western Canadian barleys (Hordeum vulgare L.). Can J Plant Sci, 1995,75:371-376. |
| [20] | Yalçin E, çelik S, Akar T, Sayim I, Köksel H. Effects of genotype and environment on β-glucan and dietary fiber contents of hull-less barleys grown in Turkey. Food Chem, 2007,101:171-176. |
| [21] |
Houston K, Russell J, Schreiber M, Halpin C, Oakey H, Washington J M, Booth A, Shirley N, Burton R A, Fincher G B, Waugh R. A genome wide association scan for (1,3;1,4)-beta-glucan content in the grain of contemporary 2-row Spring and Winter barleys. BMC Genomics, 2014,15:907.
doi: 10.1186/1471-2164-15-907 pmid: 25326272 |
| [22] | Mohammadi M, Endelman J B, Nair S, Chao S, Jones S S, Muehlbauer G J, Ullrich S E, Baik B, Wise M L, Smith K P. Association mapping of grain hardness, polyphenol oxidase, total phenolics, amylose content, and β-glucan in US barley breeding germplasm. Mol Breed, 2014,34:1229-1243. |
| [23] |
Little A, Schwerdt J G, Shirley N J, Khor S F, Neumann K, O Donovan L A, Lahnstein J, Collins H M, Henderson M, Fincher G B. Revised phylogeny of the cellulose synthase gene superfamily: insights into cell wall evolution. Plant Physiol, 2018,177:1124-1141.
doi: 10.1104/pp.17.01718 pmid: 29780036 |
| [24] | Garcia-Gimenez G, Russell J, Aubert M K, Fincher G B, Houston K. Barley grain (1,3;1,4)-β-glucan content: effects of transcript and sequence variation in genes encoding the corresponding synthase and endohydrolase enzymes. Sci Rep(UK), 2019,9:17250. |
| [25] | Oziel A, Hayes P M, Chen F Q, Jones B. Application of quantitative trait locus mapping to the development of winter-habit malting barley. Plant Breed, 2010,115:43-51. |
| [26] | Islamovic E, Obert D E, Oliver R E, Harrison S A, Ibrahim A, Marshall J M, Miclaus K J, Hu G, Jackson E W. Genetic dissection of grain beta-glucan and amylose content in barley (Hordeum vulgare L.). Mol Breed, 2013,31:15-25. |
| [27] | Panozzo J F, Eckermann P J, Mather D E, Moody D B, Black C K, Collins H M, Barr A R, Lim P, Cullis B R. QTL analysis of malting quality traits in two barley populations. Aust J Agric Res, 2007,58:858-866. |
| [28] | Hrmova M, Farkas V, Lahnstein J, Fincher G B. A barley xyloglucan xyloglucosyl transferase covalently links xyloglucan, cellulosic substrates, and (1,3;1,4)-β-D-glucans. J Biol Chem, 2007,282:1295112962. |
| [29] |
Hrmova M, Banik M, Harvey A J, Garrett T P J, Fincher G B. Polysaccharide hydrolases in germinated barley and their role in the depolymerization of plant and fungal cell walls. Int J Biol Macromol, 1997,21:67-72.
doi: 10.1016/s0141-8130(97)00043-3 pmid: 9283018 |
| [30] | Chen S C, Luchsinger W W. The mechanism of action of malt β-glucanases. VI. Hydrolysis of barley β-D-glucan by endo-β-glucanases from germinated barley. Arch Biochem Biophys, 1964,106:71-77. |
| [31] | Hrmova M, Fincher G B. Dissecting the catalytic mechanism of a plant β-D-glucan glucohydrolase through structural biology using inhibitors and substrate analogues. Carbohyd Res, 2007,342:1613-1623. |
| [1] | 陈玲玲, 李战, 刘亭萱, 谷勇哲, 宋健, 王俊, 邱丽娟. 基于783份大豆种质资源的叶柄夹角全基因组关联分析[J]. 作物学报, 2022, 48(6): 1333-1345. |
| [2] | 孙思敏, 韩贝, 陈林, 孙伟男, 张献龙, 杨细燕. 棉花苗期根系分型及根系性状的关联分析[J]. 作物学报, 2022, 48(5): 1081-1090. |
| [3] | 渠建洲, 冯文豪, 张兴华, 徐淑兔, 薛吉全. 基于全基因组关联分析解析玉米籽粒大小的遗传结构[J]. 作物学报, 2022, 48(2): 304-319. |
| [4] | 赵海涵, 练旺民, 占小登, 徐海明, 张迎信, 程式华, 楼向阳, 曹立勇, 洪永波. 水稻协优9308重组自交系群体白叶枯病抗性的全基因组关联分析[J]. 作物学报, 2022, 48(1): 121-137. |
| [5] | 贺军与, 钟伟, 陈云琼, 王卫斌, 熊静蕾, 蒋亚丽, 施辉蒙, 陈升位. 大麦籽粒发育进程中7种黄酮类化合物的积累特性分析[J]. 作物学报, 2021, 47(8): 1624-1630. |
| [6] | 马娟, 曹言勇, 李会勇. 玉米穗轴粗全基因组关联分析[J]. 作物学报, 2021, 47(7): 1228-1238. |
| [7] | 陈灿, 农保选, 夏秀忠, 张宗琼, 曾宇, 冯锐, 郭辉, 邓国富, 李丹婷, 杨行海. 广西水稻地方品种核心种质稻瘟病抗性位点全基因组关联分析[J]. 作物学报, 2021, 47(6): 1114-1123. |
| [8] | 靳义荣, 刘金栋, 刘彩云, 贾德新, 刘鹏, 王雅美. 普通小麦氮素利用效率相关性状全基因组关联分析[J]. 作物学报, 2021, 47(3): 394-404. |
| [9] | 魏丽娟, 申树林, 黄小虎, 马国强, 王曦彤, 杨怡玲, 李洹东, 王书贤, 朱美晨, 唐章林, 卢坤, 李加纳, 曲存民. 锌胁迫下甘蓝型油菜发芽期下胚轴长的全基因组关联分析[J]. 作物学报, 2021, 47(2): 262-274. |
| [10] | 蒋伟, 潘哲超, 包丽仙, 周福仙, 李燕山, 隋启君, 李先平. 马铃薯资源晚疫病抗性的全基因组关联分析[J]. 作物学报, 2021, 47(2): 245-261. |
| [11] | 张帆, 杨茜. 大麦-双季稻轮作体系有机物料与化肥配施对大麦资源利用效率及产量的影响[J]. 作物学报, 2021, 47(12): 2522-2531. |
| [12] | 雷维, 王瑞莉, 王刘艳, 袁芳, 孟丽姣, 邢明礼, 徐璐, 唐章林, 李加纳, 崔翠, 周清元. 甘蓝型油菜容重及其相关性状的全基因组关联分析[J]. 作物学报, 2021, 47(11): 2099-2110. |
| [13] | 谢磊, 任毅, 张新忠, 王继庆, 张志辉, 石书兵, 耿洪伟. 小麦穗发芽性状的全基因组关联分析[J]. 作物学报, 2021, 47(10): 1891-1902. |
| [14] | 徐婷婷, 汪巧玲, 邹淑琼, 狄佳春, 杨欣, 朱银, 赵涵, 颜伟. 基于高通量测序的大麦InDel标记开发及应用[J]. 作物学报, 2020, 46(9): 1340-1350. |
| [15] | 陶爱芬,游梓翊,徐建堂,林荔辉,张立武,祁建民,方平平. 基于黄麻转录组序列SNP位点的CAPS标记开发与验证[J]. 作物学报, 2020, 46(7): 987-996. |
|
||