欢迎访问作物学报,今天是
作物学报 ›› 2021, Vol. 47 ›› Issue (11): 2091-2098.doi: 10.3724/SP.J.1006.2021.01083
张福彦1( ), 程仲杰1, 陈晓杰1, 王嘉欢1, 陈锋2, 范家霖1, 张建伟1,*(), 杨保安1,*()
), 程仲杰1, 陈晓杰1, 王嘉欢1, 陈锋2, 范家霖1, 张建伟1,*(), 杨保安1,*()
ZHANG Fu-Yan1(), CHENG Zhong-Jie1, CHEN Xiao-Jie1, WANG Jia-Huan1, CHEN Feng2, FAN Jia-Lin1, ZHANG Jian-Wei1,*(), YANG Bao-An1,*()
摘要:
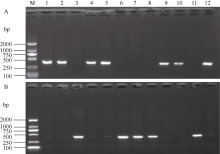
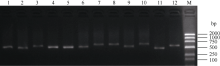
采用特异性引物PCR扩增方法对183份黄淮海麦区的小麦品种(系)的粒重基因TaCwi-A1、TaGw8-B1和TaGS-D1的等位变异进行分子鉴定, 并结合2016—2017和2017—2018年度的千粒重表型数据, 分析不同等位变异类型对小麦粒重的影响, 从而找出优势基因型组合。结果表明, 不同年份间参试品种(系)的千粒重差异达到极显著水平(P<0.01); TaCwi-A1位点上发现TaCwi-A1a和TaCwi-A1b两种等位变异, 其分布频率分别为66.7%和33.3%; TaGw8-B1位点上TaGw8-B1a等位变异分布频率较高, 为94.5%, 而TaGw8-B1b等位变异分布频率极低, 仅为5.5%; TaGS-D1位点上发现TaGS-D1a和TaGS-D1b两种等位变异, 其分布频率分别为79.8%和20.2%。不同等位变异组合的品种千粒重存在显著差异(P<0.05), 其中具有3个高千粒重等位变异组合TaCwi-A1a/TaGS-D1a/TaGw8-B1a品种的平均千粒重最高, 与具有TaCwi-A1b/TaGS-D1a/TaGw8-B1a品种的平均千粒重差异不显著, 但是显著高于其他组合(P<0.05)。TaCwi-A1a/TaGS-D1a/TaGw8-B1b基因型组合小麦品种(系)的平均千粒重最低。TaCwi-A1、TaGw8-B1和TaGS-D1位点上的不同等位变异均会导致小麦千粒重的显著变化, 其中TaGw8-B1和TaGS-D1位点上的等位变异对小麦粒重的影响更为重要。在参试材料中没有发现具有3个低千粒重等位变异组合TaCwi-A1b/TaGS-D1b/TaGw8-B1b的品种, 在7种不同等位变异组合中, 具有3个高千粒重等位变异组合TaCwi-A1a/TaGS-D1a/TaGw8-B1a品种的平均千粒重最高, 是优势基因型组合。
| [1] | 刘志勇, 王道文, 张爱民, 梁翰文, 吕慧颖, 邓向东, 葛毅强, 魏珣, 杨维才. 小麦育种行业创新现状与发展趋势. 植物遗传资源学报, 2018, 19: 430-434. |
| Liu Z Y, Wang D W, Zhang A M, Liang H W, Lyu H Y, Deng X D, Ge Y Q, Wei X, Yang W C. Current status and perspective of wheat genomics, genetics, and breeding. J Plant Genet Resour, 2018, 19: 430-434 (in Chinese with English abstract). | |
| [2] | Zanke C D, Ling J, Plieske J, Kollers S, Ebmeyer E, Korzun V, Argillier O, Stiewe G, Hinze M, Neumann F, Eichhorn A, Polley A, Jaenecke C, Ganal M W, Röder M S. Analysis of main effect QTL for thousand grain weight in European winter wheat (Triticum aestivum L.) by genome-wide association mapping. Front Plant Sci, 2015, 6: 644. |
| [3] | 宋健民, 戴双, 李豪圣, 程敦公, 刘爱峰, 曹新有, 刘建军, 赵振东. 山东省近年来审定小麦品种农艺和品质性状演变分析. 中国农业科学, 2013, 46: 1114-1126. |
| Song J M, Dai S, Li H S, Cheng D G, Liu A F, Cao X Y, Liu J J, Zhao Z D. Evolution of agronomic and quality traits of wheat cultivars released in Shandong province recently. Sci Agric Sin, 2013, 46: 1114-1126 (in Chinese with English abstract). | |
| [4] | 曹廷杰, 赵虹, 王西成, 崔党群, 詹克慧. 河南省半冬性小麦品种主要农艺性状的演变规律. 麦类作物学报, 2010, 30: 439-442. |
| Cao T J, Zhao H, Wang X C, Cui D Q, Zhan K H. Evolution of main agronomic traits for semi winter wheat varieties in Henan province. J Triticeae Crops, 2010, 30: 439-442 (in Chinese with English abstract). | |
| [5] |
Ma D Y, Yan J, He Z H, Wu L, Xia X C. Characterization of a cell wall invertase gene TaCwi-A1 on common wheat chromosome 2A and development of functional markers. Mol Breed, 2012, 29: 43-52.
doi: 10.1007/s11032-010-9524-z |
| [6] |
Jiang Y M, Jiang Q Y, Hao C Y, Hou J, Wang L F, Zhang H N, Zhang S N, Chen X H, Zhang X Y. A yield-associated gene TaCWI, in wheat: its function, selection and evolution in global breeding revealed by haplotype analysis. Theor Appl Genet, 2015, 128: 131-143.
doi: 10.1007/s00122-014-2417-5 |
| [7] | Zhang Y J, Liu J D, Xia X C, He Z H. TaGS-D1, an ortholog of rice OsGS3, is associated with grain weight and grain length in common wheat. Mol Breed, 2014, 34: 1097-1107. |
| [8] | Wang S S, Zhang X F, Chen F, Cui D Q. A single-nucleotide polymorphism of TaGS5 gene revealed its association with kernel weight in Chinese bread wheat. Front Plant Sci, 2015, 6: 1166. |
| [9] | Wang S S, Yan X F, Wang Y Y, Liu H M, Cui D Q, Chen F. Haplotypes of the TaGS5-A1 gene are its association with high thousand-kernel weight in Chinese bread wheat. Front Plant Sci, 2016, 7: 783. |
| [10] | Hanif M, Gao F M, Liu J D, Wen W E, Zhang Y J, Rasheed A, Xia X C, He Z H, Cao S H. TaTGW6-A1, an ortholog of rice TGW6, is associated with grain weight and yield in bread wheat. Mol Breed, 2016, 36: 1‒8. |
| [11] |
Hu M J, Zhang H P, Cao J J, Zhu X P, Wang S X, Jiang H, Wu Z Y, Lu J, Chang C, Sun G L, Ma C X. Characterization of an IAA-glucose hydrolase gene TaTGW6 associated with grain weight in common wheat (Triticum aestivum L.). Mol Breed, 2016, 36: 1-11.
doi: 10.1007/s11032-015-0425-z |
| [12] |
Yang J, Zhou Y J, Wu Q H, Chen Y X, Zhang P P, Zhang Y, Hu W G, Wang X C, Zhao H, Dong L L, Han J, Liu Z Y, Cao T J. Molecular characterization of a novel TaGL3-5A allele and its association with grain length in wheat (Triticum aestivum L.). Theor Appl Genet, 2019, 132: 1799-1814.
doi: 10.1007/s00122-019-03316-1 |
| [13] |
Ma M, Zhao H X, Li Z J, Hu S W, Song W N, Liu X L. TaCYP78A5 regulates seed size in wheat (Triticum aestivum). J Exp Bot, 2015, 67: 1397-1410.
doi: 10.1093/jxb/erv542 |
| [14] | 司文洁, 吴林楠, 郭利建, 周梦蝶, 刘香利, 马猛, 赵惠贤. 小麦粒重相关基因TaCYP78A5功能标记开发及验证. 作物学报, 2019, 45: 1905-1911. |
| Si W J, Wu L N, Guo L J, Zhou M D, Liu X L, Ma M, Zhao H X. Development and validation of the functional marker of grain weight-related gene TaCYP78A5 in wheat (Triticum aestivum L.). Acta Agron Sin, 2019, 45: 1905-1911 (in Chinese with English abstract). | |
| [15] |
Yan X F, Zhao L, Ren Y, Dong Z D, Cui D Q, Chen F. Genome-wide association study revealed that the TaGW8 gene was associated with kernel size in Chinese bread wheat. Sci Rep, 2019, 9: 2702.
doi: 10.1038/s41598-019-38570-2 |
| [16] | 寇程, 高欣, 李立群, 李扬, 王中华, 李学军. 小麦粒重基因TaGW2-6A等位变异的组成分析及育种选择. 作物学报, 2015, 41: 1640-1647. |
| Kou C, Gao X, Li L Q, Li Y, Wang Z H, Li X J. Composition and selection of TaGW2-6A alleles for wheat kernel weight. Acta Agron Sin, 2015, 41: 1640-1647 (in Chinese with English abstract). | |
| [17] | 时佳, 白璐, 任毅, 穆培源, 梁晓东, 玛依拉, 耿洪伟. 新疆小麦TaGW2-6A、TaCwi-A1、TaSus2-2B等位变异对粒重的影响及应用. 分子植物育种, 2018, 16: 848-858. |
| Shi J, Bai L, Ren Y, Mu P Y, Liang X D, Ma Y L, Geng H W. Effects and application of allelic variation of TaGW2-6A, TaCwi-A1 and TaSus2-2B on grain weight of Xinjiang wheat. Mol Plant Breed, 2018, 16: 848-858 (in Chinese with English abstract). | |
| [18] |
Chen F, Zhang F Y, Xia X C, Dong Z D, Cui D Q. Distribution of puroindoline alleles in bread wheat cultivars of the Yellow and Huai valley of China and discovery of a novel puroindoline a allele without PINA protein. Mol Breed, 2012, 29: 371-378.
doi: 10.1007/s11032-011-9553-2 |
| [19] |
Cui F, Ding A M, Li J, Zhao C H, Li X F, Feng D S, Wang X Q, Wang L, Gao J R, Wang H G. Wheat kernel dimensions: how do they contribute to kernel weight at an individual QTL level. J Genet, 2011, 90: 409-425.
pmid: 22227928 |
| [20] |
Wu W, Li C J, Ma B L, Shah F, Liu Y, Liao Y C. Genetic progress in wheat yield and associated traits in China since 1945 and future prospects. Euphytica, 2014, 196: 155-168.
doi: 10.1007/s10681-013-1033-9 |
| [21] |
Rasheed A, Xia X C, Ogbonnaya F, Mahmood T, Zhang Z W, Mujeeb-Kazi A, He Z H. Genome-wide association for grain morphology in synthetic hexaploid wheats using digital imaging analysis. BMC Plant Biol, 2014, 14: 128.
doi: 10.1186/1471-2229-14-128 pmid: 24884376 |
| [22] |
Gupta P K, Rustgi S, Kumar N. Genetic and molecular basis of grain size and grain number and its relevance to grain productivity in higher plants. Genome, 2006, 49: 565-571
doi: 10.1139/g06-063 |
| [23] | 相吉山, 穆培源, 桑伟, 聂迎彬, 徐红军, 庄丽, 崔凤娟, 韩新年, 邹波. 小麦粒重基因 TaCwi-A1功能标记CWI22、CWI21的验证及应用. 中国农业科学, 2014, 47: 2671-2679. |
| Xiang J S, Mu P Y, Sang W, Nie Y B, Xu H J, Zhuang L, Cui F J, Han X N, Zou B. Validation and application of function markers CWI22 and CWI21 of TaCwi-A1 gene related to wheat kernel weight. Sci Agric Sin, 2014, 47: 2671-2679 (in Chinese with English abstract). | |
| [24] | 刘永伟, 周硕, 王雪征, 孙果忠, 朱金永, 韩秋芬, 李春杰, 赵和, 王海波. 粒重基因TaCwi-A1等位变异在黄淮麦区小麦品种(系)中的分布及功能分析. 华北农学报, 2017, 32: 131-137. |
| Liu Y W, Zhou S, Wang X Z, Sun G Z, Zhu J Y, Han Q F, Li C J, Zhao H, Wang H B. Functional analysis and distribution of allelic variations of TaCwi-A1 gene related to kernel weight in Yellow and Huai River Valleys facultative wheat zone. Acta Agric Boreali-Sin, 2017, 32: 131-137 (in Chinese with English abstract). | |
| [25] |
Lu J, Chang C, Zhang H P, Wang S X, Sun G L, Xiao S H, Ma C X. Identification of a novel allele of TaCKX6a02 associated with grain size, filling rate and weight of common wheat. PLoS One, 2015, 10: e0144765.
doi: 10.1371/journal.pone.0144765 |
| [26] | 简大为, 周阳, 刘宏伟, 杨丽, 买春艳, 于立强, 韩新年, 张宏军, 李洪杰. 利用功能标记揭示新疆小麦改良品种与地方品种的遗传变异. 作物学报, 2018, 44: 657-671. |
| Jian D W, Zhou Y, Liu H W, Yang L, Mai C Y, Yu L Q, Han X N, Zhang H J, Li H J. Functional markers reveal genetic variations in wheat improved cultivars and landraces from Xinjiang. Acta Agron Sin, 2018, 44: 657-671 (in Chinese with English abstract). |
| [1] | 张钰坤, 陆赢, 崔看, 夏石头, 刘忠松. 芥菜种子颜色调控基因TT8的等位变异及其地理分布分析[J]. 作物学报, 2022, 48(6): 1325-1332. |
| [2] | 胡文静, 李东升, 裔新, 张春梅, 张勇. 小麦穗部性状和株高的QTL定位及育种标记开发和验证[J]. 作物学报, 2022, 48(6): 1346-1356. |
| [3] | 郭星宇, 刘朋召, 王瑞, 王小利, 李军. 旱地冬小麦产量、氮肥利用率及土壤氮素平衡对降水年型与施氮量的响应[J]. 作物学报, 2022, 48(5): 1262-1272. |
| [4] | 付美玉, 熊宏春, 周春云, 郭会君, 谢永盾, 赵林姝, 古佳玉, 赵世荣, 丁玉萍, 徐延浩, 刘录祥. 小麦矮秆突变体je0098的遗传分析与其矮秆基因定位[J]. 作物学报, 2022, 48(3): 580-589. |
| [5] | 冯健超, 许倍铭, 江薛丽, 胡海洲, 马英, 王晨阳, 王永华, 马冬云. 小麦籽粒不同层次酚类物质与抗氧化活性差异及氮肥调控效应[J]. 作物学报, 2022, 48(3): 704-715. |
| [6] | 刘运景, 郑飞娜, 张秀, 初金鹏, 于海涛, 代兴龙, 贺明荣. 宽幅播种对强筋小麦籽粒产量、品质和氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 716-725. |
| [7] | 马红勃, 刘东涛, 冯国华, 王静, 朱雪成, 张会云, 刘静, 刘立伟, 易媛. 黄淮麦区Fhb1基因的育种应用[J]. 作物学报, 2022, 48(3): 747-758. |
| [8] | 徐龙龙, 殷文, 胡发龙, 范虹, 樊志龙, 赵财, 于爱忠, 柴强. 水氮减量对地膜玉米免耕轮作小麦主要光合生理参数的影响[J]. 作物学报, 2022, 48(2): 437-447. |
| [9] | 王洋洋, 贺利, 任德超, 段剑钊, 胡新, 刘万代, 郭天财, 王永华, 冯伟. 基于主成分-聚类分析的不同水分冬小麦晚霜冻害评价[J]. 作物学报, 2022, 48(2): 448-462. |
| [10] | 陈新宜, 宋宇航, 张孟寒, 李小艳, 李华, 汪月霞, 齐学礼. 干旱对不同品种小麦幼苗的生理生化胁迫以及外源5-氨基乙酰丙酸的缓解作用[J]. 作物学报, 2022, 48(2): 478-487. |
| [11] | 马博闻, 李庆, 蔡剑, 周琴, 黄梅, 戴廷波, 王笑, 姜东. 花前渍水锻炼调控花后小麦耐渍性的生理机制研究[J]. 作物学报, 2022, 48(1): 151-164. |
| [12] | 孟颖, 邢蕾蕾, 曹晓红, 郭光艳, 柴建芳, 秘彩莉. 小麦Ta4CL1基因的克隆及其在促进转基因拟南芥生长和木质素沉积中的功能[J]. 作物学报, 2022, 48(1): 63-75. |
| [13] | 韦一昊, 于美琴, 张晓娇, 王露露, 张志勇, 马新明, 李会强, 王小纯. 小麦谷氨酰胺合成酶基因可变剪接分析[J]. 作物学报, 2022, 48(1): 40-47. |
| [14] | 李玲红, 张哲, 陈永明, 尤明山, 倪中福, 邢界文. 普通小麦颖壳蜡质缺失突变体glossy1的转录组分析[J]. 作物学报, 2022, 48(1): 48-62. |
| [15] | 罗江陶, 郑建敏, 蒲宗君, 范超兰, 刘登才, 郝明. 四倍体小麦与六倍体小麦杂种的染色体遗传特性[J]. 作物学报, 2021, 47(8): 1427-1436. |
|
||