





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
高密度种植条件下去叶对不同株型夏玉米群体及单叶光合性能的调控
刘铁宁 , 徐彩龙, 谷利敏, 董树亭
, 徐彩龙, 谷利敏, 董树亭*
, 徐彩龙, 谷利敏, 董树亭
|
|


* 通讯作者(Corresponding author): 董树亭, E-mail:stdong@sdau.edu.cn, Tel: 0538-8245838
第一作者联系方式: E-mail:liutiening0319@163.com
当前玉米产量的提高部分归因于种植密度的增加, 但过高的种植密度使冠层中下部叶片光照条件变差, 致使单株生长速率降低。因此, 如何缓解该条件下群体光合与单株光合性能的矛盾成为当前玉米高产栽培中急需解决的问题。为此, 在种植密度为105 000株 hm-2的大田试验条件下, 研究了紧凑型玉米品种郑单958及半紧凑型玉米品种金海5号的群体光合速率(CAP)、叶面积指数(LAI)、穗位叶净光合速率(
The improved grain yield of modern maize hybrids is mainly due to the increased plant density. However, the solar radiation intercepted by ear leaves was reduced, and the growth rate of individual plant was suppressed under high plant density. Consequently, how to coordinate the relationship between canopy apparent photosynthesis (CAP) and individual leaf photosynthesis capacity under high plant density is a vital question we are facing. A field experiment was conducted using summer maize (
现代玉米单产提升部分归因于种植密度的提高[ 1]。增加种植密度, 提高光温资源利用率, 依靠群体发挥增产潜力是获得高产的重要措施之一[ 2]。但过高的种植密度使玉米群体下部叶片光照条件变差, 叶片早衰, 群体光合速率降低[ 3], 同时也会加剧植株间对水肥资源的竞争, 致使单株生长速率降低, 增加减产风险[ 4]。随着玉米生产条件的改善以及种植密度的再提高, 有报道认为产量的进一步提高有赖于源质量性能的改善[ 5]。
研究表明, 冠层中部叶片是玉米籽粒产量的主要来源, 其光合强度与产量密切相关[ 6]。高密度条件下, 群体通风透光条件较差, 穗位叶层光照强度仅为自然光强的18%~55%[ 7], 同时CO2浓度也较低, 穗位叶片光合能力受到限制。另有研究发现, 密度的增大势必造成资源的重新分配, 即更多的资源用于个体竞争能力的增强, 如根系、叶面积的相对增大造成生长冗余, 会过度消耗能量和营养物质造成减产[ 8]。因此, 去除部分过度生长的营养器官(叶片或根系)可使植物对获取的有限物质和能量进行合理的分配和利用, 提高物质和能量的利用率[ 9, 10, 11]。郝梦波等[ 12]发现去除1/2叶片可改善群体冠层内部的光照状况并最终提高穗位叶 Pn, 叶片Chl含量及气孔导度也显著增加[ 13]。针对去叶对植株干物质积累及运转问题, 前人亦做了大量研究[ 14, 15, 16, 17]。Zhu等[ 18]研究表明, 去叶使茎秆氮素积累量显著降低。Tollenaar等[ 19]发现去叶增加了茎秆和叶片花前储存干物质向籽粒的运转, 弥补了因源不足造成的同化物供应的减少, 维持了籽粒灌浆进程的正常进行。去叶也可显著调控植物的衰老进程, 影响植物体内的抗氧化代谢[ 20]。去叶增强了超氧化物歧化酶(SOD)和过氧化物酶(POD)活性, 延缓了植株的衰老[ 21, 22], 而去除穗位叶以外的其他叶片将加速穗位叶片的衰老[ 23]。
关于高密度种植条件下通过去叶措施缓解两株型夏玉米群体及单株光合性能矛盾的研究较少, 因此, 本研究比较高密度种植条件下冠层结构改变后两株型玉米生育后期群体光合、单叶光合及抗氧化代谢特性对去叶的响应, 探讨不同品种在不同去叶处理下群体光合与单株光合性能的协调特征, 为高密度栽培条件下稳定或提高单株生产能力探讨新的技术途径, 同时也为耐密高产品种选育提供借鉴。
在山东农业大学黄淮海区域玉米技术创新中心(36°10′ N, 117°09′ E), 2012年6月15日播种, 10月7日收获。试验地为沙质壤土, 耕层0~20 cm土壤含有机质11.4 g kg-1、全氮0.71 g kg-1、碱解氮57.1 mg kg-1、速效磷25.6 mg kg-1、速效钾107.2 mg kg-1。供试品种为紧凑型玉米品种郑单958 (Zhengdan 958)和半紧凑型玉米品种金海5号(Jinhai 5)。两品种种植密度均为105 000株 hm-2, 行距为60 cm。
采用随机区组设计, 小区面积3 m × 20 m, 4次重复, 每小区种植630株玉米。取样和单株光合性能的测定在3个重复小区中进行, 第4个重复小区则用于群体光合速率的测定。各小区均施纯氮120 kg hm-2、P2O572 kg hm-2、K2O 96 kg hm-2。氮肥以开沟形式于拔节期施入50%, 大喇叭口期施入50%; P2O5、K2O则全部底施。在整个生育期内保证良好的水分供应, 及时浇水、除草、灭虫, 保证植株有良好的生长环境。
玉米开花后3 d做去叶处理, 即分别去除植株顶部2片叶(S1)、4片叶(S2)、6片叶(S3), 以不去叶植株为对照(S0), 每处理去叶植株为2520株。去叶时仔细且迅速, 尽量减少植株其他部位的折损。
各处理于花后0、13、26、39和52 d取样, 每次选取生长一致且有代表性植株5株, 取其穗位叶片, 液氮速冻30 min, -40℃冰箱保存, 用作酶活性及叶绿素含量的测定。
1.2.1 叶面积指数(LAI) 从每小区选取生长健壮且长势均匀的健康植株5株, 于开花期、灌浆期和乳熟期、蜡熟期和完熟期测定。单叶叶面积= L (叶片最大长度)×W (最大宽度)×0.75, LAI=单株叶面积×单位土地面积株数/单位土地面积。
1.2.2 群体透光率(LT) 采用LP-80冠层分析仪, 于开花期和灌浆中期(吐丝后30 d)选择晴朗无风的天气, 将棍棒式探头平行横伸于行内测定群体冠层上方、穗位叶层和群体底部(距地表10 cm处)透光率, 每个小区5个重复。
透光率 (%) = I/I0×100%
式中, I0为冠层上方无障碍物时的光照强度, I为通过一定叶层之后的辐射强度。
1.2.3 群体光合速率(CAP)的测定 参照董树亭[ 24]方法并加以改进。分别于开花期、灌浆期、乳熟期和蜡熟期, 选择晴朗无云的天气, 用GXH-305型红外线CO2分析仪于10:00—12:00在田间直接测定, 测定时间为60 s, 3次重复, 在测定时间范围内, 箱内温度升高1℃左右, 对测定影响可忽略不计。同化箱用铝合金制成, 外罩透明聚酯薄膜, 其透光率在95%左右, 箱体分为上下两部分, 每个箱体高度为1.4 m, 长为1.2 m, 宽为1 m, 罩2行12株玉米。每层箱体内安装3只由电池驱动的60瓦风机以搅匀箱体内空气, 测定时下部箱体与水槽间填满水, 水槽在大口期提前安装到地里, 以确保测定时箱体的密封性, 同时使测定更简单易行。群体光合速率CAP (µmol CO2 m-2s-1) = ΔC × V/106 × 60/Δm × 44/22.4 × 6.313 × 273/(273 + T)/S, 式中, ΔC为间隔时间内前后2次测定的二氧化碳浓度差值(µL L-1); V为同化箱体积(L); Δm为测定时间(min); T为同化箱内温度(℃); S为测定群体所占土地面积(m2)。
1.2.4 穗位叶净光合速率( Pn)和叶绿素荧光参数的测定 于开花授粉前选择生长一致且有代表性植株5株挂牌标记, 自开花之日起每隔13 d左右选择晴朗无云的天气, 于上午10:00至12:00用Ciras-II型便携式光合仪(PP-Systems, UK)测定穗位叶净光合速率。同时用FMS2脉冲调制式荧光仪(Hansatech, UK)测定光适应下稳态荧光( Fs)、最大荧光( Fm)和暗适应20 min后的最大荧光( Fm)、初始荧光( Fo)。光系统II的实际光化学效率 ΦPSII=( Fm- Fs)/ Fm、光系统II的最大光化学效率 Fv/ Fm =( Fm- Fo)/ Fm。
1.2.5 叶绿素含量的测定 采用Arnon试验方法[ 25]测定穗位叶叶绿素含量。
1.2.6 酶类活性测定 将叶片表面尘土和污物洗净并擦干, 去除叶脉, 取叶片同一部位测定酶活性。称取0.5 g冷藏叶片, 加入5 mL预冷的磷酸缓冲液(pH 7.8), 冰浴研磨至匀浆, 于4℃下12 000× g离心20 min, 上清液即为酶提取液。用赵世杰等[ 26]的方法测定MDA含量; 按照Giannopolitis等[ 27]的方法测定SOD活性; 参照Klapheck等[ 28]的方法测定POD活性。
用Sigmaplot 10.0作图, 采用DPS 7.05软件统计分析并用LSD法检验处理间显著水平( P<0.05)。
2.1.1 叶面积指数 不同去叶处理对2种株型玉米花后LAI有显著影响(图1)。S2和S3处理自去叶后LAI一直处于较低水平, 灌浆期LAI表现为S0>S1>S2>S3, 差异均达显著水平。随灌浆进程的推进, 乳熟期后LAI表现为S1>S0>S2>S3, 且S1显著高于其他处理。完熟期, 以对照S0为基准, 郑单958 S1处理LAI较对照升高15.89%, 而金海5号S1较对照升高25.54%。由此可知, 高密度种植条件下适当去除植株顶部2片叶有利于维持籽粒灌浆期间LAI高值持续期, 且对半紧凑型品种金海5号调控效果更为明显。
2.1.2 群体透光率(LT) 由图2可知, 灌浆中期两株型玉米相同层次的透光率均随去叶程度的增强而增加, 穗位层透光率处理间差异均达显著水平, 而底层透光率除S1与S0差异不显著外, S2和S3均显著高于S0, 表明高密度群体去叶能显著改善穗位层光照状况。
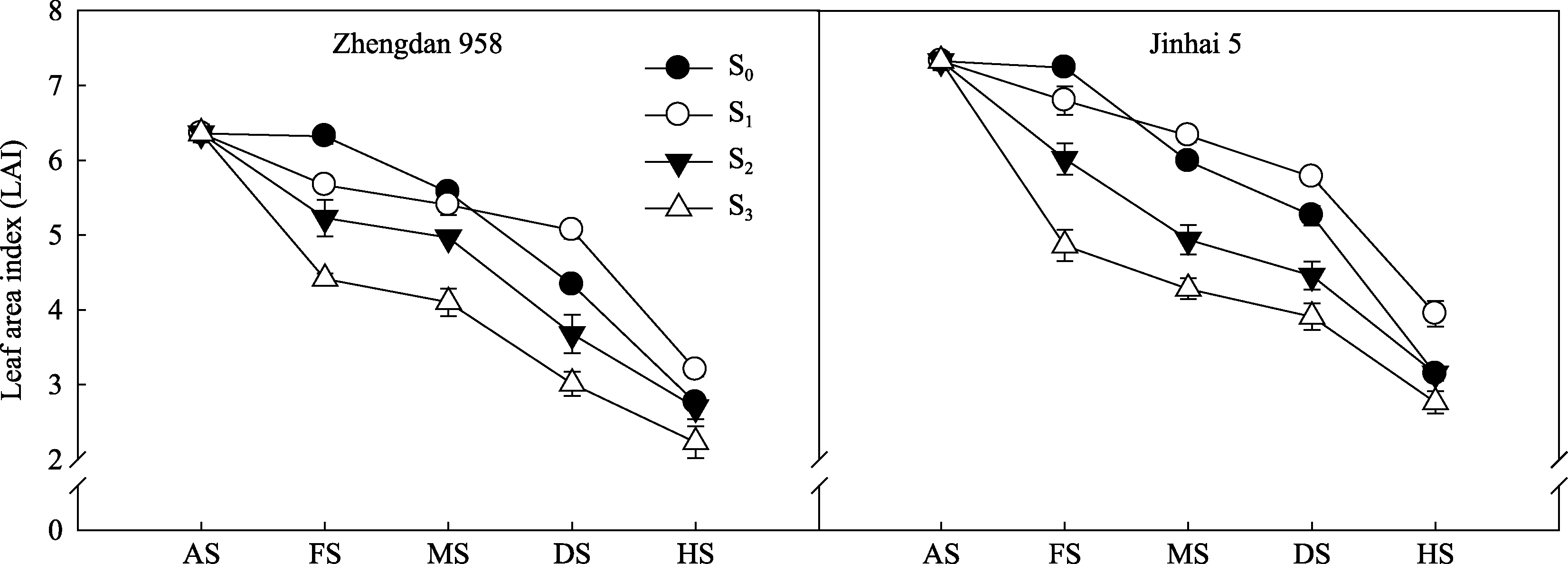 | 图1 去叶对2种株型玉米叶面积指数(LAI)的影响AS: 开花期; FS: 灌浆期; MS: 乳熟期; DS: 蜡熟期; HS: 完熟期。Fig. 1 Effects of leaf removal on leaf area index of two plant types of maizeAS: anthesis stage; FS: filling stage; MS: milking stage; DS: dough stage; HS: harvest stage. |
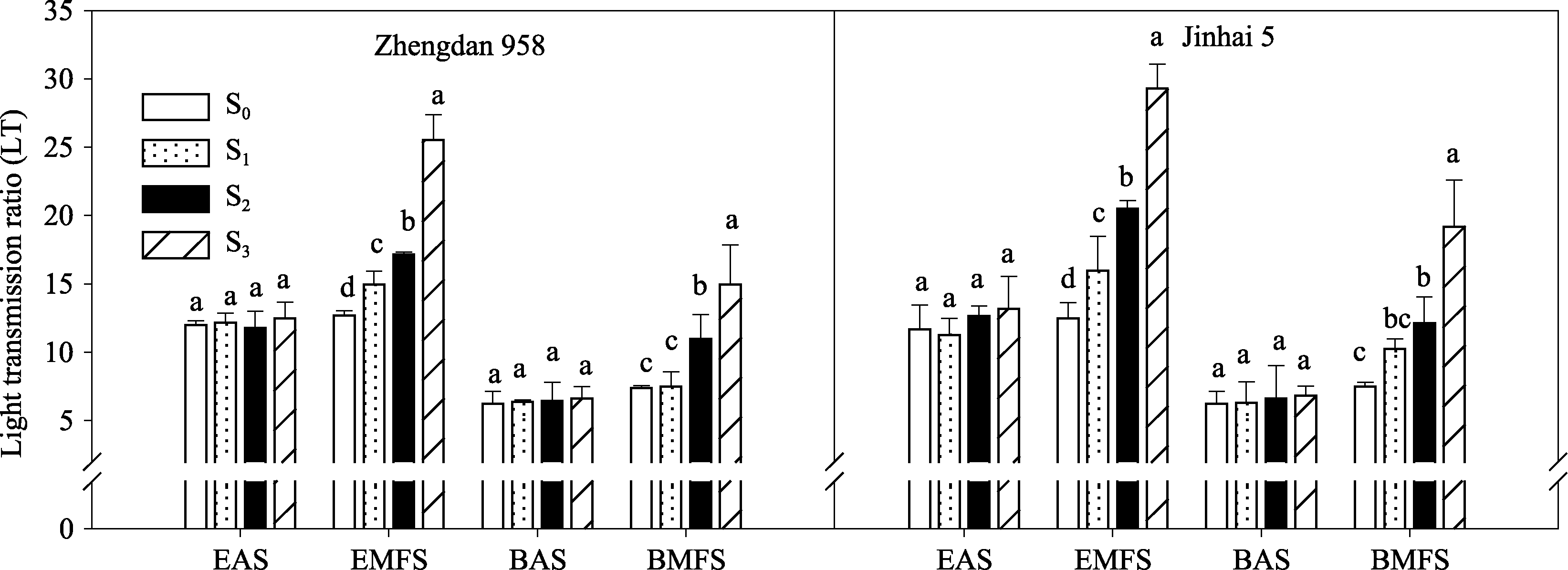 | 图2 去叶对2种株型玉米冠层透光率(LT)的影响EAS: 开花期穗位层; EMFS: 灌浆中期穗位层; BAS: 开花期底层; BMFS: 灌浆中期底层。Fig. 2 Effects of leaf removal on light transmission ratio (LT) of two plant types of maizeEAS: ear leaf layer at anthesis stage; EMFS: ear leaf layer at mid-filling stage; BAS: bottom layer at anthesis stage; BMFS: bottom layer at mid-filling stage. |
开花期和灌浆中期各处理相比较, 灌浆中期群体内不同层次透光率均高于开花期, 以开花期为基准, 灌浆中期紧凑型品种郑单958穗位层透光率各处理依次升高5.84%、24.71%、43.24%和104.47%; 半紧凑型品种金海5号依次升高6.86%、36.93%、75.58%和122.32%。可见, 去叶对半紧凑型玉米金海5号冠层光照状况调控效果更为明显。
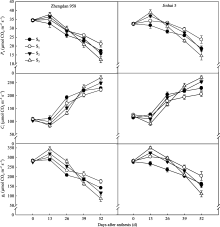
高密度种植条件下不同程度去叶对两株型玉米CAP的调控效果不同(图3)。S0和S1处理花后CAP均呈现先升高后降低的变化趋势, 灌浆期达到最大值, 而后呈不同程度下降; 而S2和S3处理自花后一直呈现下降趋势。花后两株型品种CAP各生育时期均表现为S1>S0>S2>S3, 且S1显著高于其他处理。
 | 图3 去叶对两种株型玉米群体光合速率(CAP)的影响AS: 开花期; FS: 灌浆期; MS: 乳熟期; DS: 蜡熟期。Fig. 3 Effects of leaf removal on canopy apparent photosynthesis (CAP) of two plant types of maizeAS: anthesis stage; FS: filling stage; MS: milking stage; DS: dough stage. |
郑单958 S1处理灌浆期之前保持较高CAP, 但之后下降迅速, 平均每天下降1.12 µmol CO2 m-2 s-1, 而金海5号S1处理CAP在产量形成的关键时期高值持续期较长, 下降趋势较缓, 平均每天下降0.811 µmol CO2 m-2 s-1。在蜡熟期, 郑单958 S1处理CAP较对照高23.85%, 而S2和S3处理分别降低31.43%和42.14%; 金海5号S1处理CAP较对照高38.08%, S2和S3处理分别降低26.36%和44.77%。由此可见, 适度去除植株顶部2片叶有利于改善生育后期CAP, 且对金海5号影响较大。
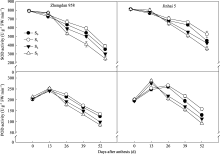
2.3.1 穗位叶光合特性 不同去叶处理对两株型玉米 Pn、 gs和 Ci的调控效果不同(图4)。花后0~26 d去六叶处理穗位叶 Pn和 gs显著高于其他处理, 之后迅速下降。自花后26 d起, 穗位叶 Pn、 gs均表现S1最大, 显著高于其他处理。以花后52 d为例, 郑单958穗位叶净光合速率 S1较S0升高22.58%, S2、S3分别降低8.80%和28.37%; 而金海5号S1升高26.67%, S2和S3依次降低3.65%和22.70%。与此同时, Ci变化趋势与 Pn相反。由此可知, 适度去叶(即去除顶部两片叶)有利于增加生育后期穗位叶净光合速率, 而过度去叶(S2和S3)虽可提高灌浆前期净光合速率, 但之后下降迅速, 不利于光合产物的积累。
品种间比较, 去叶前金海5号穗位叶净光合速率低于郑单958, 而去叶后测定结果表明, 金海5号S1处理穗位叶净光合速率灌浆中后期均高于郑单958, 表明适度去叶更有利于半紧凑型玉米金海5号光合能力的提高。
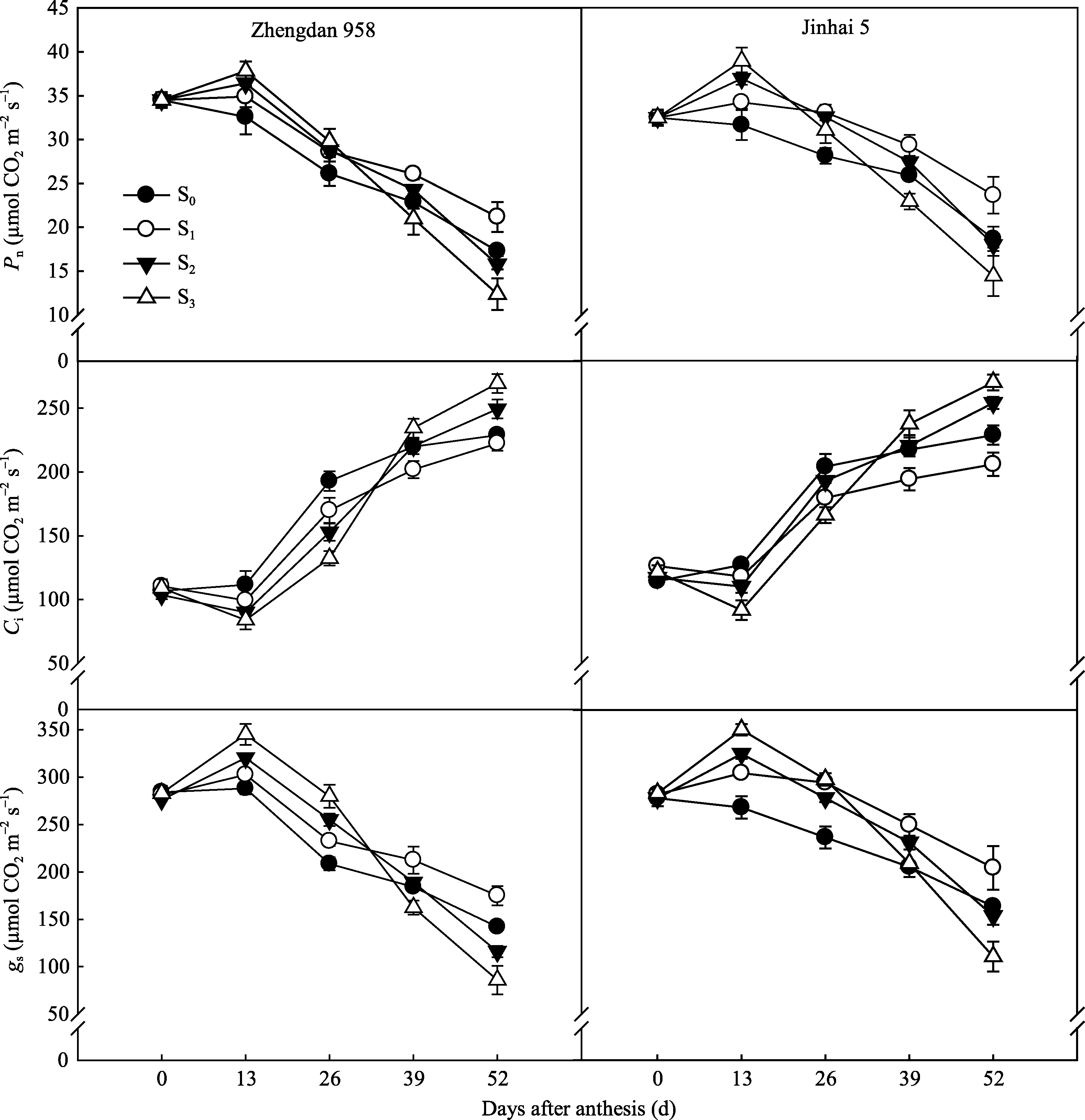 | 图4 去叶对两种株型玉米穗位叶 Pn、 gs和 Ci的影响Fig. 4 Effects of leaf removal on net photosynthetic rate ( Pn), stomatal conductance ( gs), and intercellular CO2 concentration ( Ci) in ear leaves of two plant types of maize |
2.3.2 穗位叶荧光特性 由图5可知, 花后13 d, 郑单958穗位叶 Fv/ Fm随去叶程度的增强有所增加, 而之后S1具有较高的 Fv/ Fm。花后52 d, 两品种S1处理 Fv/ Fm和 ΦPSII均显著高于其他处理。去叶对 Fv/ Fm和 ΦPSII的影响亦因品种类型而异。金海5号去二叶处理在籽粒灌浆期间 Fv/ Fm和 ΦPSII高值持续期较郑单958长。表明密度为105 000株 hm-2的种植条件下, 去除植株顶部2片叶有利于植株光能利用效率的提高, 且金海5号提高幅度较大。
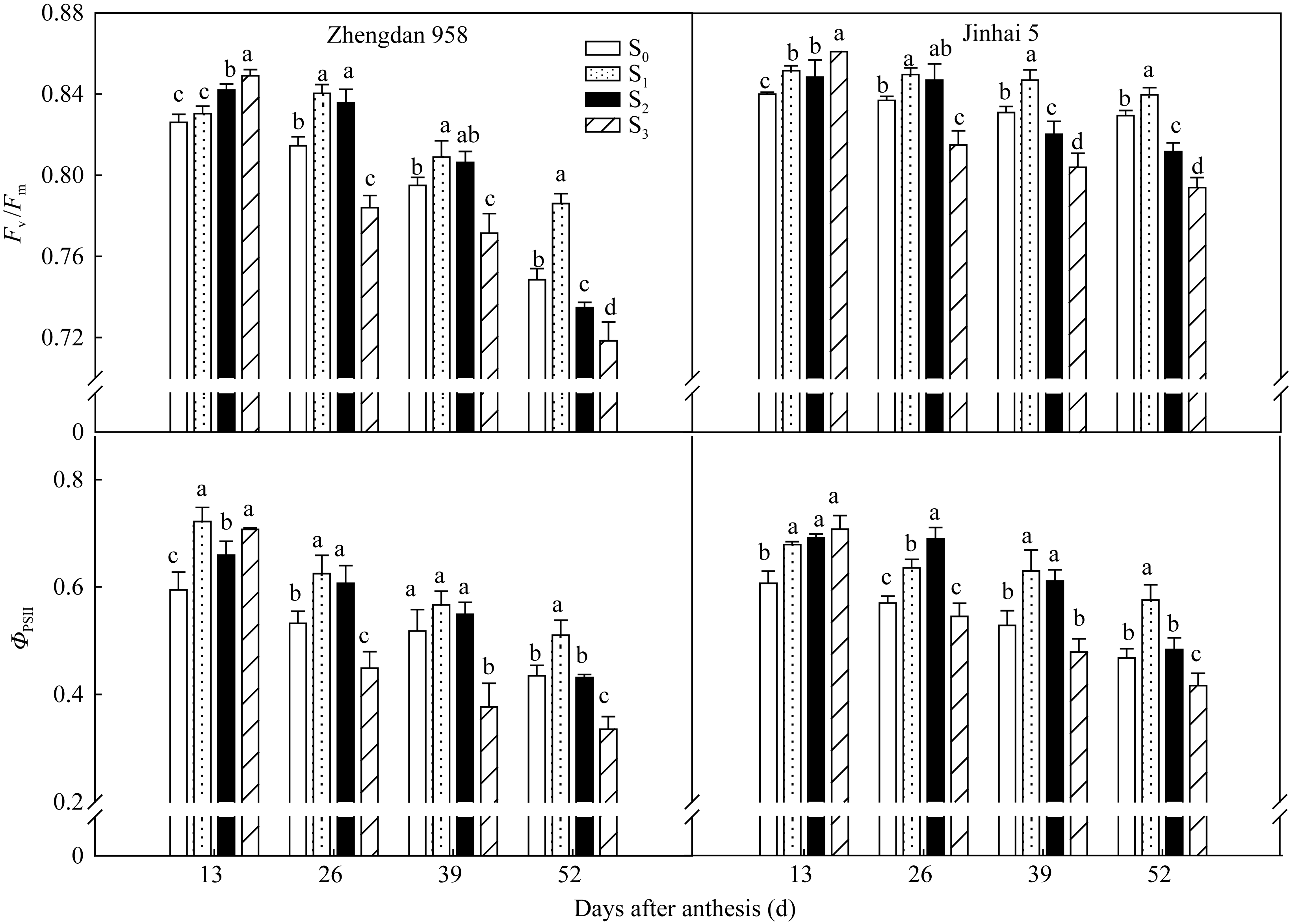 | 图5 去叶对2种株型玉米穗位叶 Fv/ Fm和 ΦPSII的影响Fig. 5 Effects of leaf removal on Fv/ Fm and ΦPSII in ear leaves of two plant types of maize |
花后玉米穗位叶叶绿素含量呈先升高后降低的变化趋势, 花后13 d达到最大值, 之后呈现不同程度降低(图6)。花后13 d, S2和S3处理玉米穗位叶叶绿素含量均显著高于S0, 且增幅随去叶程度的增强而增大, 之后过度去叶植株(S2和S3)叶绿素含量迅速下降; 花后13~52 d, 穗位叶叶绿素含量表现为S1>S0>S2>S3, 且S1显著高于其他处理。结果表明, 适度去叶使穗位叶在灌浆中后期保持较高的叶绿素含量, 有利于光合作用的进行, 而过度去叶只提高了生育前期的叶绿素含量。
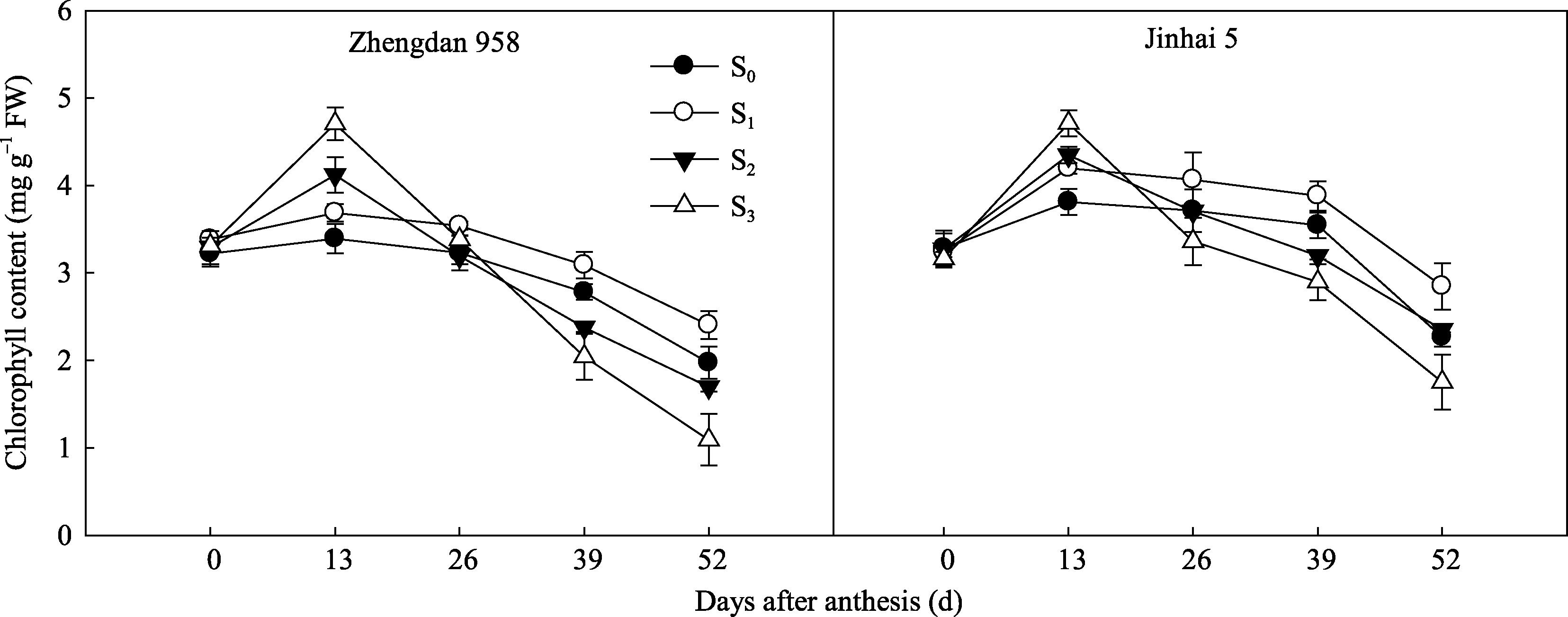 | 图6 去叶对2种株型玉米穗位叶叶绿素含量的影响Fig. 6 Effects of leaf removal on chlorophyll content in ear leaves of two plant types of maize |
2.5.1 MDA含量 开花至灌浆后期两品种穗位叶丙二醛(MDA)含量呈不断增加的变化趋势。郑单958穗位叶MDA含量花后13 d迅速增加, 而金海5号于花后26 d迅速增加, 较郑单958推迟13 d左右。灌浆中后期, MDA含量表现为S3>S2>S0>S1, 且S1处理显著低于其他处理(图7)。表明高密度种植条件下去二叶植株穗位叶膜脂过氧化程度较低, 便于其在灌浆期保持较高的生理活性, 而过度去叶则加重了穗位叶膜脂过氧化程度。
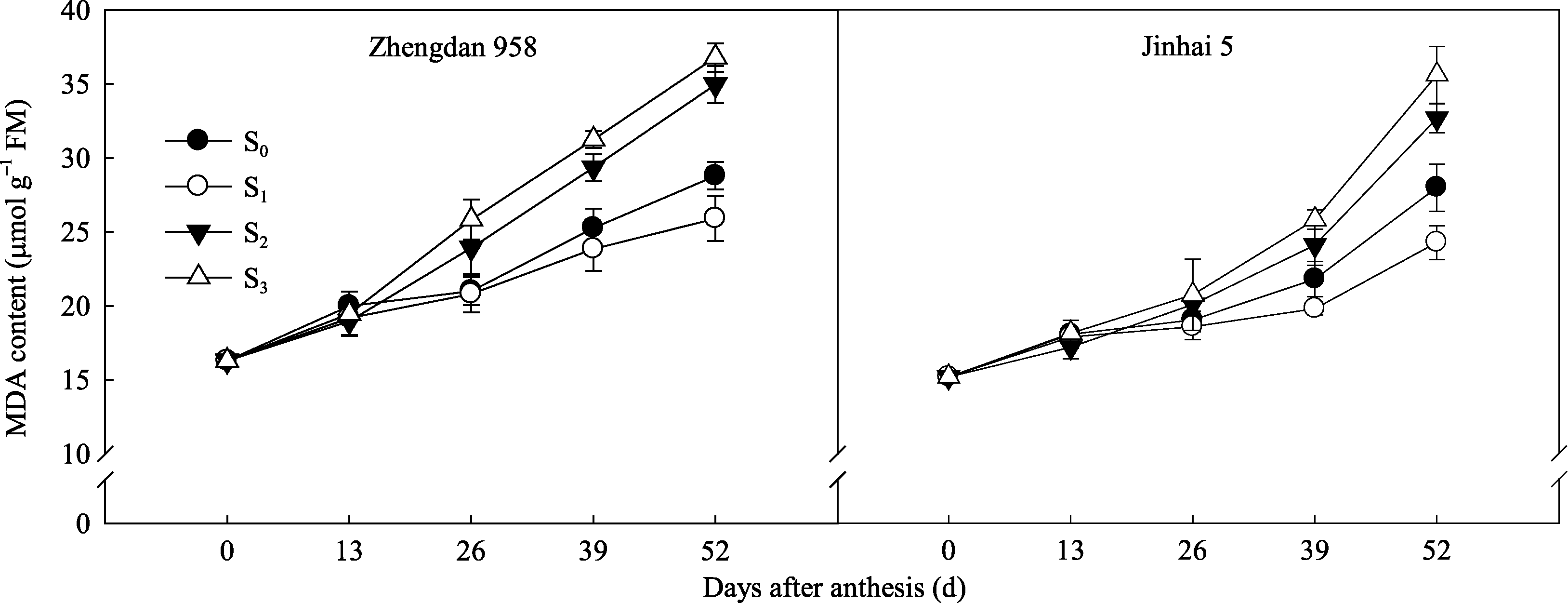 | 图7 去叶对2种株型玉米穗位叶MDA含量的影响Fig. 7 Effects of leaf removal on MDA content in ear leaves of two plant types of maize |
2.5.2 抗氧化代谢酶活性 各处理花后穗位叶SOD活性均呈下降趋势, 而不同去叶处理对其影响程度不同, 开花26 d后S1处理明显增强, 而S2和S3处理则显著低于S1处理; 花后52 d, 郑单958 S1处理较对照S0高9.97%, 而S2和S3处理则分别降低14.48%和29.89%; 金海5号S1高于对照16.34%, S2和S3分别下降5.86%和20.22% (图8)。POD活性自花后13 d起变化趋势与SOD一致。由此可知, 过度去叶加速了叶片的衰老, 而去除植株顶部2片叶有利于灌浆中后期叶片活性氧清除能力的提高, 延缓了叶片的衰老, 且对半紧凑型品种金海5号的调控作用更大。
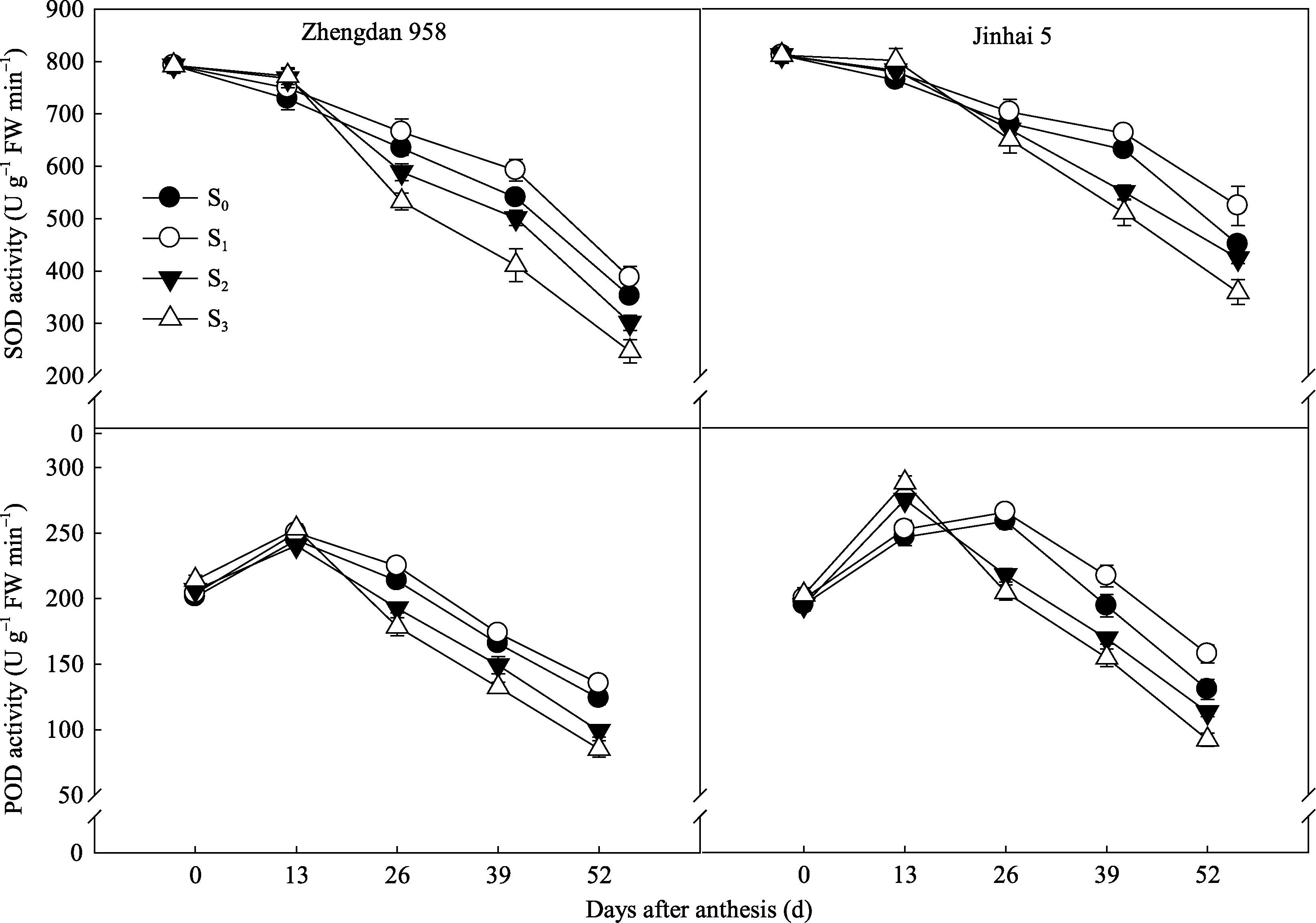 | 图8 去叶对2种株型玉米穗位叶抗氧化酶活性的影响Fig. 8 Effects of leaf removal on antioxidant enzyme activity in ear leaves of two plant types of maize |
由表1可知, 去除植株顶部4片叶和6片叶(S2和S3)使穗粒数和穗长大幅降低, 由过度去叶造成的光合源强度的大幅减少显著降低了籽粒产量; 而高密度种植条件下适当去除植株顶部2片叶(S1)后穗粒数较S0无明显变化, 但千粒重显著增加, 其籽粒产量显著高于其他处理。两品种剪叶处理后穗粗与S0相比均无显著差异。
| 表1 不同去叶处理下两株型玉米产量及产量构成因素 Table 1 Grain yield and yield components in two plant types of maize with leaf removal treatment |
功能叶片的光合作用与其光截获量密切相关[ 29]。研究表明, 去除部分叶片能改善冠层内部光照条件, 增加剩余叶片的净光合速率及气孔导度[ 12, 13, 30]。本试验条件下, 去叶对穗位叶净光合速率的影响因去叶强度而异: 花后0~13 d各去叶处理 Pn均有所增加, 此后去四叶与去六叶植株穗位叶 Pn、 Fv/ Fm和 ΦPSII迅速下降, 这可能一定程度上与植株过早衰老有关[ 20]; 而去二叶植株在籽粒有效灌浆期间有较强的单叶光合能力, 表明去除植株顶部2片叶有利于剩余穗位叶片净光合速率及光能利用率的提高, 这与郝梦波等[ 12]研究结果一致。此外, 本试验中 gs与 Pn变化趋势一致, 而 Ci与 Pn变化趋势相反, 表明去叶对植株 Pn的影响主要受非气孔因素限制。分析原因发现, 这一定程度上与去叶后冠层内光照情况显著改善有关, 例如, 灌浆中期郑单958和金海5号去二叶处理穗位层透光率较对照分别增加24.71%和36.93%, 改善了的光照条件有利于单株光合作用。
在作物正常的生长代谢过程中(比如光合作用和呼吸作用), 细胞中依旧有活性氧的存在[ 31]。一旦植株体内活性氧的产生与清除平衡机制遭到破坏, 植株将遭受活性氧迫害[ 32]。然而, 植物体内存在一系列的活性氧清除酶系(SOD、POD等), 它们能有效地清除活性氧, 使植株在受到胁迫的情况下保持较长时间的功能期[ 33, 34]。前人研究发现, 源库比是影响植株叶片衰老的重要因素之一[ 35, 36]。Borrás等[ 37]认为抑制部分库器官授粉, 相对较大的源库比例延缓了玉米叶片的衰老。Tollenaar和Daynard等[ 23]的试验表明去除穗位叶以外的其他叶片加速了穗位叶的衰老, 而Dreccer等[ 20]认为去叶与植株的衰老状况无关。本试验结果表明, 高密度种植条件下去除植株顶部2片叶使穗位叶在生育后期具有显著增高的SOD、POD活性, 同时MDA含量较低; 而去除顶部4片叶和6片叶植株仅在花后13 d内保持较高的SOD、POD活性, 之后迅速下降, 同时MDA含量显著升高, 叶片衰老进程被加速。由此可知, 密度为105 000株 hm-2种植条件下, 去除植株顶部2片叶改善了高密度群体的源库关系, 使叶片在籽粒形成关键时期保持较高的抗氧化酶活性, 及时清除植物体内的有害物质, 延缓了叶片的衰老, 这是保持较高净光合速率的酶学基础, 使叶片在花后13~52 d籽粒有效灌浆期间保持显著升高的 Pn; 而过度去叶(去四叶和去六叶)植株由于源库比例失调, 活性氧产生与清除平衡体系被破坏, 抗氧化酶活性较低, 膜脂过氧化作用增强, 这是其 Pn下降的主要原因。
冠层结构是影响作物群体光分布和光合特性的因素之一[ 7]。高密度种植条件下适宜的叶面积指数及有效的空间配置有利于形成高效的冠层结构, 便于同化物的形成[ 38, 39]。本试验研究结果表明, 适当去除植株顶部2片叶延长了高密度群体乳熟期后LAI高值持续期, 同化源较充足, 便于籽粒灌浆[ 40]。冠层透光率的变化趋势与LAI相反, 穗位层透光率随去叶程度的增强显著增加, 这与郝梦波等[ 12]研究结果基本一致; 而去除四片叶和六片叶虽显著改善了冠层内部光照条件, 但过多的漏光损失不利于光能利用率的提高。
群体光合速率与冠层结构密切相关, 而种植密度是调控冠层最简便的措施[ 7, 24]。随密度的进一步提高, 群体光合速率呈降低趋势[ 3]。本研究发现, 高密度种植条件下去二叶群体自去叶后一直保持较高的CAP, 而去4片叶和6片叶玉米群体光合速率自始至终一直保持较低水平。2种株型玉米去六叶处理之所以在乳熟期前保持较低CAP, 主要是较低的叶面积指数所致, 尽管此时穗位叶 Pn较高; 而适度去除顶部2片叶, 两株型玉米CAP自花后一直保持较高水平, 且显著高于其他处理, 与其适宜的LAI及灌浆中后期较好的 Pn及改善了的冠层通风透光条件有关, 有效地缓解了密植条件下两株型玉米群体光合与单株光合性能的矛盾, 保证了在较高叶面积指数的前提下单株生产力的提高。前人研究发现, 高密度群体中的个体为了增强自身的竞争力, 形成了较大的叶面积, 这会造成营养及能量的浪费, 去除这些多余的叶片可使植物对获取的有限物质和能量进行最合理、最优化的分配, 提高现存能量的利用率, 获得较高的籽粒产量[ 9, 10, 11], 本研究结果与之一致。
胡昌浩等[ 41]研究发现, 在适宜低密度下紧凑型品种具有较高的群体光合速率, 而平展型品种大口期以后具有较高的群体光合速率; 徐庆章等[ 42]通过人工改型后测定群体光合速率表明, 改为紧凑型玉米比改为平展型玉米高19.09%。然而, 本研究结果表明, 高密度种植下, 适度去除植株顶部2片叶对CAP调控效果明显, 例如, 花后52 d, 郑单958较对照升高23.85%, 而金海5号则升高38.08%, 这与金海5号玉米植株去除顶部两叶片后较高的LAI有关, 同时较好的抗氧化酶活性及较低的膜脂过氧化程度保证了较高的 Pn。
在高密度种植条件下, 去除植株顶部2片叶使两种株型玉米在灌浆中后期保持较高LAI及较长的穗位叶 Pn高值持续期, 同时抗氧化酶活性强、膜脂过氧化程度低, 在籽粒形成和灌浆的关键阶段保持较高CAP, 有效缓解了高密度种植条件下两株型玉米生育后期群体光合与单叶光合的矛盾, 保证了较高LAI前提下具有较好的单株光合性能, 最终获得较高的籽粒产量。此外, 半紧凑型玉米品种金海5号对去除顶部2片叶响应更为敏感。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|