
{kind=link}
{kind=link}
{kind=link}
玉米生产上3个主推品种光合特性、干物质积累转运及灌浆特性
[徐田军*  , 吕天放
, 吕天放* , 赵久然* , 王荣焕* , 陈传永, 刘月娥, 刘秀芝, 王元东, 刘春阁]
, 吕天放, 赵久然, 王荣焕, 陈传永, 刘月娥, 刘秀芝, 王元东, 刘春阁]
|
|


第一作者联系方式: 徐田军, E-mail: xtjxtjbb@163.com; 吕天放, E-mail: 314565358@qq.com
以当前玉米生产主推品种郑单958、先玉335和京科968为试验材料, 考察其光合特性、干物质积累与转运及籽粒灌浆特性, 以揭示高产玉米品种的产量形成特性, 为玉米高产生产提供依据。结果表明: (1)产量以京科968最高、先玉335次之、郑单958最低, 京科968分别较郑单958和先玉335高14.55%和7.93%。(2)穗位叶净光合速率和冠层光合能力表现为京科968>先玉335>郑单958, 且吐丝期>乳熟期。京科968吐丝期和乳熟期的穗位叶净光合速率分别比先玉335高7.84%和16.78%, 比郑单958高22.23%和24.44%; 冠层光合能力分别较先玉335高38.77%和58.41%, 较郑单958高50.83%和56.49%。(3)花后干物质积累量、转移量、干物质转运率和干物质转运对籽粒贡献率均以京科968最高, 分别比先玉335高13.72%、21.20%、6.32%和4.77%, 比郑单958高31.87%、39.96%、18.49%和10.42%。(4)籽粒灌浆参数在不同品种间存在较大差异, 京科968与先玉335的平均灌浆速率(0.73和0.75 g 100-grain-1 d-1)相当, 且均高于郑单958 (0.67 g 100-grain-1 d-1); 活跃灌浆期以郑单958 (53.69 d)最长、京科968 (51.02 d)次之、先玉335 (48.95 d)最短。(5)相关分析表明, 产量与净光合速率显著正相关, 与花后干物质积累量及转运率极显著正相关。京科968具有较高的光合效率、花后干物质积累量及转运率、灌浆速率及较长的灌浆持续期, 是较郑单958和先玉335高产的重要原因。
, LYU Tian-Fang, ZHAO Jiu-Ran, WANG Rong-Huan, CHEN Chuan-Yong, LIU Yue-E, LIU Xiu-Zhi, WANG Yuan-Dong, LIU Chun-Ge
The objective of this study was to explore yield-forming characteristics of high yield maize varieties. Using three widely cultivated maize varieties Zhengdan 958 (ZD958), Xianyu 335 (XY335) and Jingke 968 (JK968), we tested grain yield, photosynthetic characteristics, dry matter accumulation and translocation, and grain filling characteristics. The yield of JK968 was the highest, and was 14.55% and 7.93% higher than that of ZD958 and XY335, respectively. The photosynthetic rate and canopy photosynthetic capacity showed JK968 > XY335 > ZD958 and silking stage > milk stage. The ear photosynthetic rate of JK968 at silking stage and milk stage was 7.84% and 16.78% higher than that of XY335, as well as 22.23% and 24.44% higher than that of ZD958, respectively. The photosynthetic capacity of JK968 at silking stage and milk stage was 38.77% and 58.41% higher than that of XY335, as well as 50.83% and 56.49% higher than that of ZD958, respectively. Dry matter accumulation after silking stage, the transfer amount of dry matter, translocation efficiency of dry matter and contribution to grain dry matter for JK968 were the highest, with 13.72%, 21.20%, 6.32%, and 4.77% higher than those of XY335, as well as 31.87%, 39.96%, 18.49%, and 10.42% higher than those of ZD958, respectively. There was a big difference in grain filling parameters among those different varieties, the average filling rate of JK968 (0.73 g 100-grain-1 d-1) and XY335 (0.75 g 100-grain-1 d-1) was higher than that of ZD958 (0.67 g 100-grain-1 d-1) and the active grain filling period of ZD958 (53.69 d) was longer than that of the JK968 (51.02 d) and XY335 (48.95 d). Maize yield was significantly correlated with net photosynthetic rate at P < 0.05, and with the dry matter accumulation after silking stage and the translocation rate at P < 0.01. Thus, JK968 had higher photosynthetic efficiency, dry matter accumulation after silking stage, translocation rate, grain filling rate and longer grain filling period, which is the main reason that JK968 had the higher yield than ZD958 and XY335.
玉米是我国种植面积最大、总产量最高的第一大粮食作物, 对保障国家粮食安全和满足市场需要发挥着主力军作用[1, 2]。玉米产量主要取决于穗粒数和粒重[3], 而粒重与光合特性、干物质积累分配、灌浆速率、灌浆持续期等密切相关[4, 5, 6]。
光合作用为作物的产量形成提供主要物质基础[7, 8]。叶片是作物光合作用的主要场所, 90%的干物质来自叶片的光合作用[9, 10], 光合特性与产量密切相关[11, 12]。提高玉米叶片的光合速率, 延长其光合有效功能期, 促进光合产物积累及转运, 有利于提高粒重, 进而增加产量[13, 14, 15]。光合产物对玉米籽粒干物质的贡献很大, 此外花前及花后临时贮藏在茎叶鞘等营养器官中的干物质的贡献也很重要[16, 17]。研究表明, 在一定范围内玉米干物质积累量与产量呈正相关, 干物质积累越多, 产量越高; 茎、叶、鞘等营养器官的干物质积累、转移与分配是玉米籽粒产量形成的重要因素, 其中叶片和苞叶的干物质转运量对籽粒贡献率较大, 茎鞘和穗轴的贡献率相对较小[18, 19, 20]。玉米籽粒的灌浆持续期和灌浆速率决定了粒重和产量[21, 22], 籽粒灌浆特性受播期[23, 24, 25]、密度[26]、肥料[27]等外部环境条件的影响, 但主要取决于品种的遗传特性[28]。
目前, 围绕我国玉米生产主推大品种的产量、光合效率、干物质积累分配及灌浆特性等系统深入研究的相关报道还较少。本文就此展开深入研究, 旨在为解析玉米高产机制和指导玉米高产高效生产提供理论参考。
以我国玉米生产中种植面积66.7万公顷以上的主推大品种郑单958 (ZD958)、先玉335 (XY335)和京科968 (JK968)为试验材料。2015年和2016年, 在北京市昌平区国家精准农业科技示范园区开展试验, 两年均于5月10日播种, 10月7日收获。试验田土壤0~20 cm耕层含有机质1.11%、全氮0.08%、有效氮65.6 μ g g-1、速效磷65.2 μ g g-1和速效钾124.02 μ g g-1。
12行区, 行长5 m, 行距0.60 m, 小区面积36 m2。随机区组排列, 3次重复。留苗密度为67 500株 hm-2。管理同当地大田生产。
1.3.1 叶面积指数 在三叶期、拔节期、大喇叭口期、吐丝期、乳熟期和成熟期测定每小区生长一致的6株。
叶面积=叶片最大长度× 最大宽度× 0.75
叶面积指数(LAI)=单株叶面积× 单位土地面积株数/单位土地面积
1.3.2 叶片光合速率及气体交换参数的测定 吐丝前选择生长一致的植株挂牌标记。分别在吐丝期和乳熟期, 用LI-6400XT光合测定仪测定穗位叶的净光合速率(Pn, μ mol CO2 m-2 s-1)。选择晴朗天气上午10:00— 12:00, 测定每处理3株, 重复3次。采用开放式气路, 穗位叶附近的CO2浓度为400 μ mol mol-1左右, 光合有效辐射为1600 μ mol m-2 s-1。
1.3.3 冠层光合能力 冠层光合能力是净光合速率和叶面积指数的综合反映。冠层光合能力=净光合速率(Pn)× 叶面积指数(LAI)[29]。
1.3.4 干物质积累、分配及转运 吐丝期和成熟期, 取代表性植株5株, 吐丝期按茎、叶、鞘, 成熟期按茎、叶、鞘、雌穗(穗轴、苞叶和籽粒)分别进行处理。105℃杀青30 min, 然后80℃烘干至恒重, 分别称重。计算单株玉米器官干物质转运量、转运率等[30]。
花后干物质积累量(kg hm-2) = 成熟期地上部干物质积累量-吐丝期地上部干物质积累量
干物质转移量(kg hm-2) = 吐丝期地上部干物质积累量-成熟期地上部营养器官干物质积累量
干物质转运率(%) = 干物质转移量/吐丝期地上部干物质积累量× 100
干物质转移对籽粒的贡献率(%)=干物质转移量/籽粒干重× 100
收获指数=籽粒产量/收获时总干物质重
1.3.5 籽粒灌浆参数 吐丝前选择生长健壮一致的果穗挂牌标记, 统一授粉。于授粉后第15、22、29、36、43、50、57、64、71和78 d取样。每次从各小区取3穗, 取中部籽粒100粒, 称鲜重, 在105℃烘箱中杀青30 min后, 80℃烘干至恒重, 测定最终百粒重。以授粉后天数(d)为自变量、授粉后每隔7 d测得的百粒重为因变量(W), 参照朱庆森等[31]的方法, 利用Richards方程W=A(1+Be-Ct)-1/D模拟籽粒灌浆过程。籽粒灌浆速率: F = ACBe-Ct/(1+Be-Ct)(D+1)/D, 式中: W为粒重(g); A为最终粒重(g); t为授粉后天数(d); B、C、D为回归方程所确定的参数, 计算下列灌浆特征参数:
最大灌浆速率Gmax= (CWmax/D)[1-(Wmax/A)D]
平均灌浆速率Gave= AC/(2D+4)
达到最大灌浆速率的时间Tmax= (lnB-lnD)/C
灌浆活跃期(约完成总积累量的90%) P = 2(D+2)/C
1.3.6 产量及产量构成因素 成熟期收获每个小区中间的4行果穗, 随机选取30个考察穗行数、行粒数、千粒重。收获穗全部脱粒后自然风干, 用水分仪测定水分后, 按14%含水量折合成公顷产量。
采用Microsoft Excel 2007处理和计算数据, 采用SPSS 19.0统计软件进行方差分析和多重比较(采用LSD法), 采用SigmaPlot 10.0软件作图。
由表1可知, 2015年和2016年参试品种的生育期表现为京科968和郑单958相当, 均为126 d和127 d, 先玉335较郑单958和先玉335少3 d。
| 表1 参试品种生育期 Table 1 Growth stage of the tested varieties (d) |
由表2可知, 产量、穗粒数和百粒重在品种和年际间差异显著, 以京科968最高、先玉335次之、郑单958最低。其中, 京科968产量、实收穗数、穗粒数和百粒重分别较先玉335高7.93%、3.28%、0.23%和5.43%, 较郑单958高14.55%、2.20%、3.15%和15.53%。
| 表2 参试品种的产量及产量构成因素 Table 2 Yield and yield components of the tested varieties |
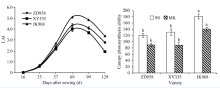
2.3.1 光合作用 由图1可知, 净光合速率、气孔导度、胞间CO2浓度和蒸腾速率在品种和生育时期间均存在显著差异。吐丝期和乳熟期的穗位叶净光合速率表现为京科968 (35.35 μ mol m-2 s-1和28.72 μ mol m-2 s-1) > 先玉335 (32.58 μ mol m-2 s-1和23.93 μ mol m-2 s-1) > 郑单958 (27.49 μ mol m-2 s-1和21.70 μ mol m-2 s-1), 且在乳熟期京科968净光合速率显著高于先玉335和郑单958, 气孔导度表现为京科968 (0.65 μ mol m-2 s-1和0.58 μ mol m-2 s-1) > 先玉335 (0.59 μ mol m-2 s-1和0.41 μ mol m-2 s-1) > 郑单958 (0.31 μ mol m-2 s-1和0.21 μ mol m-2 s-1), 而胞间CO2浓度表现为郑单958> 先玉335> 京科968; 蒸腾速率表现为先玉335> 京科968> 郑单958; 不同生育时期间, 表现为灌浆期高于乳熟期。
 | 图1 参试品种的净光合速率、气孔导度、胞间CO2浓度和蒸腾速率 SS: 吐丝期; MK: 乳熟期。ZD958: 郑单958; XY335: 先玉335; JK968: 京科968。误差线上字母不同表示品种类型间存在显著差异 (P < 0.05)。Fig. 1 Comparison of net photosynthetic rate, stomatal conductance, intercellular CO2 concentration and transpiration rate of the tested varieties SS: silking stage; MK: milking stage. ZD958: Zhengdan 958; XY335: Xianyu 335; JK968: Jingke 968. Different letters above error bars indicate significant difference between two types of varieties (P < 0.05). |
2.3.2 冠层光合能力 由图2可知, 参试品种的叶面积指数随生育期天数呈S型变化, 播种后65 d之前差别不大, 之后则差异较大, 表现为京科968> 郑单958> 先玉335。叶面积指数在吐丝期达峰值, 其中京科968最大, 为5.32。先玉335和郑单958分别为4.62和4.35。由图1 (净光合速率)和图2可知, 冠层光合能力在品种间表现为京科968> 先玉335> 郑单958, 不同生育期间表现为吐丝期> 乳熟期, 其中吐丝期京科968的冠层光合能力较先玉335和郑单958高38.77%和50.83%, 乳熟期分别高58.41%和56.49%。
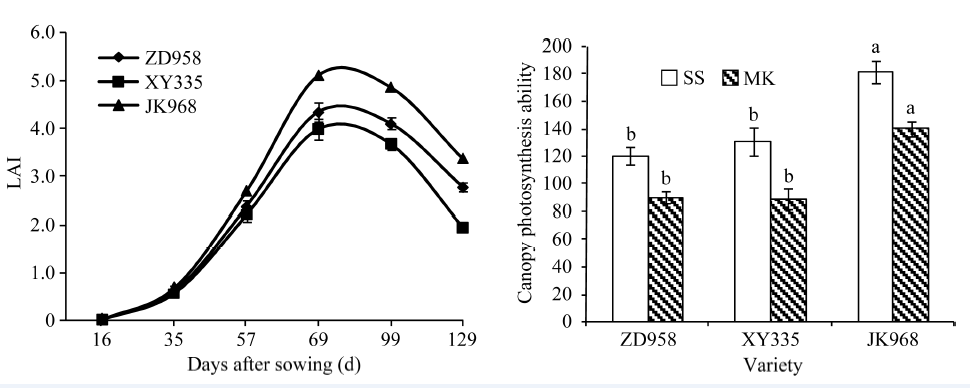 | 图2 参试品种叶面积指数和冠层光合能力 缩写同图1。误差线上字母不同表示品种类型间存在显著差异(P < 0.05)。Fig. 2 Leaf area index and canopy photosynthetic ability of the tested varieties Abbreviations are the same as those given in Fig. 1. Different letters above error bars indicate significant difference between two types of varieties (P < 0.05). |
由表3可知, 花后干物质的积累量、转移量、转运率以及干物质转运对籽粒的贡献率均表现为京科968> 先玉335> 郑单958。其中, 京科968的花后干物质积累量、转移量和转运率分别比先玉335高13.72%、21.2%、6.33%, 比郑单958高31.87%、39.96%和18.49%。收获指数在品种间差异不显著, 京科968为0.56, 郑单958和先玉335均为0.55。
| 表3 参试品种的花后干物质积累、分配及转运 Table 3 Accumulation and transportation of dry matter after silking of the tested varieties |
2.5.1 粒重和灌浆速率 由图3可知, 籽粒灌浆期间百粒重呈“ 慢— 快— 慢” 的S型变化趋势, 籽粒灌浆速率呈“ 先升后降” 的单峰变化。最大灌浆速率表现为先玉335> 京科968> 先玉335; 到达最大灌浆速率的时间以先玉335最早、郑单958次之、京科968最晚。
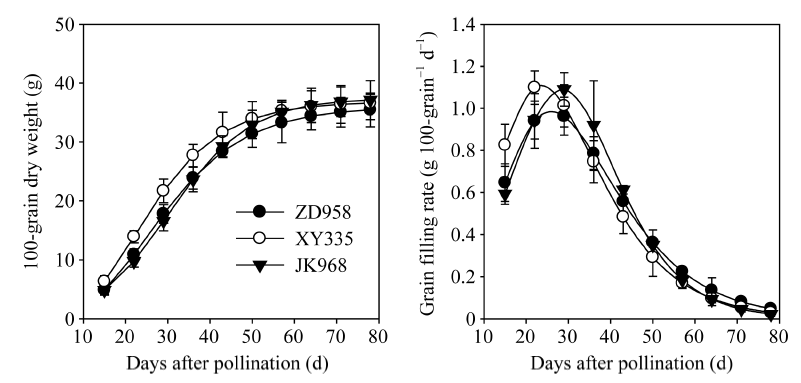 | 图3 参试品种的籽粒灌浆动态和灌浆速率 缩写同图1。Abbreviations are the same as those given in Fig. 1.Fig. 3 Grain filling dynamics and grain filling rate of the tested varieties |
2.5.2 灌浆参数 由表4可知, 用Richards模型可以较好地拟合籽粒灌浆过程, 决定系数为0.9932~0.9987。3个品种达到最大灌浆速率的时间(Tmax)以先玉335最早(23.48 d), 郑单958次之(25.97 d), 京科968最晚(28.54 d); 最大灌浆速率(Gmax)和平均灌浆速率(Gave)表现为先玉335> 京科968> 郑单958; 达到最大灌浆速率时的生长量(Wmax)表现为京科968> 先玉335> 郑单958, 活跃灌浆期(P)表现为郑单958 (53.69 d)> 京科968 (51.02 d)> 先玉335 (48.95 d)。由此可见, 郑单958虽然活跃灌浆期长, 但平均灌浆速率低; 先玉335虽然灌浆速率高, 但活跃灌浆期短, 京科968灌浆速率与先玉335相当, 但活跃灌浆期相对较长。
| 表4 参试品种的籽粒灌浆特征参数 Table 4 Grain filling parameters of the tested varieties |
由表5可知, 产量与花后干物质积累量(0.99* * )、干物质转运率(0.96* * )极显著正相关, 与净光合速率(0.97* )显著正相关, 与平均灌浆速率正相关, 但未达到显著水平, 与活跃灌浆期呈负相关, 但不显著。
| 表5 灌浆参数、干物质转运率和光合特性与产量的相关性分析 Table 5 Correlation analysis of grain filling parameters, dry matter transfer rate and photosynthetic characteristics and yield |
叶片是玉米进行光合作用的主要器官, 是截获光能的物质载体[32]。叶面积大小及光合作用强弱对玉米生长发育及产量有重要影响[33]。前人研究表明, 延长叶片持绿期及其有效光合作用时间, 提高玉米吐丝后群体光合速率, 能够显著提高产量[34]。本研究相关分析表明, 叶片光合速率与玉米产量呈显著正相关(相关系数为0.97* )。净光合速率、气孔导度、胞间CO2浓度和蒸腾速率在品种和生育时期间均存在较大差异, 且在乳熟期京科968净光合速率显著高于先玉335和郑单958。冠层光合能力是净光合速率与叶面积指数的综合反映。京科968的冠层光合能力较先玉335和郑单958高48.6%和53.7%。在灌浆期, 参试玉米品种叶面积指数随生育期推进呈下降趋势, 且先玉335和郑单958叶片衰老进程高于京科968。京科968生育后期较强的净光合速率与较高的叶面积是其获得较高产量和粒重的重要生理基础之一, 这与郑友军等研究结果一致[35]。灌浆期是玉米籽粒产量形成关键时期, 也是植株生理功能渐进衰退时期, 产量最高的京科968在乳熟期仍具有较强的光合能力, 净光合速率下降相对缓慢, 延长光合同化物的积累时间, 保证充足的光合产物供应玉米籽粒灌浆。
籽粒干物质也来自花后物质积累与转运。作物经济产量由干物质积累量决定, 花后物质生产与分配对产量具有重要影响, 光合产物的积累与分配决定作物的群体产量[36]。本研究表明, 花后干物质积累量、干物质转运率与产量之间存在极显著正相关(相关系数为0.99* * 和0.96* * ), 而不同品种的干物质积累、转运与分配差别各异, 表现为花后与花前干物质生产的比例不同, 最终表现出产量水平的差异。京科968花后干物质积累量和干物质转运量分别比郑单958高31.87%和39.96%; 干物质转移率比先玉335高6.33%、比郑单958高18.49%。干物质转移对籽粒的贡献率以京科968最高, 先玉335次之, 郑单958最低。京科968的干物质向籽粒转运与分配的能力高于先玉335和郑单958, 有利于获得较高的产量水平。
籽粒灌浆期是玉米产量形成的重要阶段。灌浆速率受基因型和环境条件共同影响, 灌浆时间和灌浆速率决定了玉米籽粒的干物质积累量, 灌浆速率和灌浆过程持续天数均与粒重显著相关[37]。马赟花等[38]和Jorge[39]研究发现玉米产量的提高主要是籽粒灌浆时间延长的结果, 建议把灌浆时间作为高产玉米选择的有效指标; 而本课题组研究认为, 灌浆速率高是提高玉米粒重和产量的重要因素[23, 24, 25]。本研究表明, 当前我国玉米生产3个主推大品种中, 郑单958活跃灌浆期长但平均灌浆速率低, 产量最低; 先玉335灌浆速率高但活跃灌浆期短, 产量中等; 京科968灌浆速率与先玉335相当, 活跃灌浆期相对较长, 产量最高。由此可见, 京科968更易实现高产, 表现出干物质转运率较高、光合速率较高、灌浆速率高、活跃灌浆期长, 进而获得高产。这与黄振喜等研究结果一致[40]。
玉米产量与花后干物质积累量、干物质转运率极显著正相关, 与净光合速率显著正相关。京科968具有较高的花后干物质积累量、干物质转移量和干物质转移率, 较高的光合速率及冠层光合能力, 以及相对较高的灌浆速率和较长的活跃灌浆期, 是较郑单958和先玉335高产的重要原因。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|