欢迎访问作物学报,今天是
作物学报 ›› 2022, Vol. 48 ›› Issue (11): 2765-2773.doi: 10.3724/SP.J.1006.2022.14143
刘星1( ), 苏良辰1, 李丽梅2, 李玲2,*()
), 苏良辰1, 李丽梅2, 李玲2,*()
LIU Xing1(), SU Liang-Chen1, LI Li-Mei2, LI Ling2,*()
摘要:
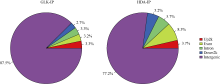
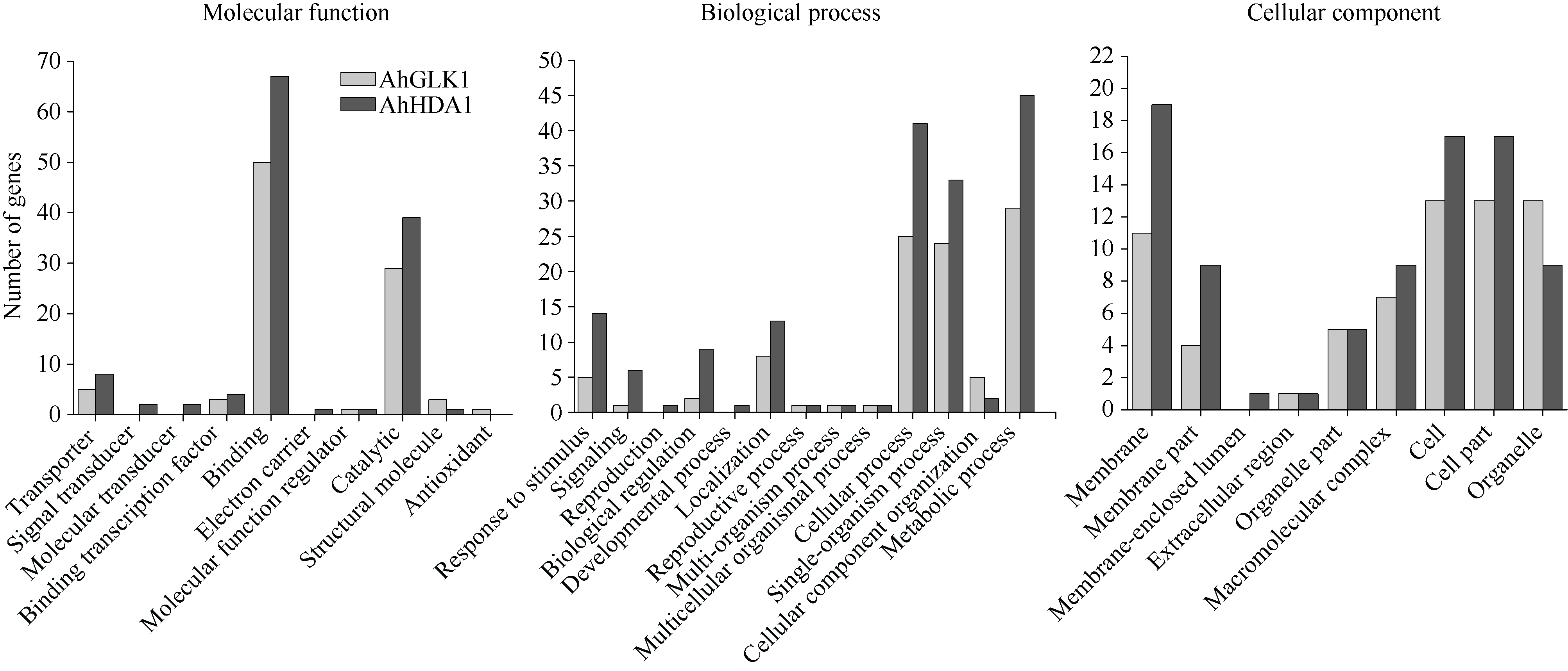
花生(Arachis hypogaea L.)是重要的经济油料作物, 其生长发育、产量与品质受干旱影响。为深入了解花生的抗旱机理, 本研究通过ChIP-seq对组蛋白去乙酰化酶AhHDA1和转录因子AhGLK1富集的DNA序列进行分析, 揭示两者调控的下游靶基因网络。通过比对分析, GLK-IP获得6571万clean beads, HDA-IP获得6390万clean beads, Input获得7006万clean beads, 唯一比对率分别为74.97%、76.81%和76.75%。GLK-IP获得714个peak, HDA-IP获得543个peak。Peak在基因的外显子、内含子、上游、下游和基因间等功能元件均有分布。GO富集结果显示, AhGLK1-IP和AhHDA1-IP的peak相关基因在分子功能中的富集分别为35.1%和32.8%, 在生物学过程中的富集分别为39.3%和44.2%, 在细胞组分中的富集分别为25.5%和22.8%。KEGG信号通路富集结果显示, AhGLK1-IP相关基因显著富集在“代谢途径(metabolic pathways)”、“抗生素生物合成(biosynthesis of antibiotics)”、“二羧酸代谢(glyoxylate and dicarboxylate metabolism)”、“不同环境中微生物代谢(microbial metabolism in diverse environments)”、“碳代谢(carbon metabolism)”、“次生代谢生物合成(biosynthesis of secondary metabolites)”和“氨基酸生物合成(biosynthesis of amino acids)。而AhHDA1-IP相关基因在“N聚糖生物合成(N-glycan biosynthesis)”、“精氨酸和脯氨酸代谢(arginine and proline metabolism)”和“苯丙氨酸代谢(phenylalanine metabolism)”通路显著富集。AhGLK1-IP和AhHDA1-IP共同富集的peak有4个, 在AhGLK1-IP和AhHDA1-IP特异富集的基序(motif)中存在共同的保守序列AGAA/T。研究结果为深入认识AhGLK1和AhHDA1基因的功能和了解花生响应干旱胁迫和旱后恢复生长中的调控机制具有参考价值。
| [1] |
Luo H, Guo J, Ren X, Chen W, Huang L, Zhou X, Chen Y, Liu N, Xiong F, Lei Y, Liao B, Jiang H. Chromosomes A07 and A05 associated with stable and major QTLs for pod weight and size in cultivated peanut (Arachis hypogaea L.). Theor Appl Genet, 2018, 131: 267-282.
doi: 10.1007/s00122-017-3000-7 |
| [2] | 邓斌, 李玲, 李晓云, 苏良辰, 崔威韬, 冯锦欣. AhHDA1异源表达影响拟南芥植株干旱性. 华南师范大学学报(自然科学版), 2016, 48(5): 52-57. |
| Deng B, Li L, Li X Y, Su L C, Cui W T, Feng J X. Heterologous expression of AhHDA1 affects drought resistance of Arabidopsis plants. J South Chin Nor (Nat Sci Edn), 2016, 48(5): 52-57. (in Chinese with English abstract) | |
| [3] |
Su L, Liu S, Liu X, Zhang B, Li M, Zeng L, Li L. Transcriptome profiling reveals histone deacetylase 1 gene overexpression improves flavonoid, isoflavonoid, and phenylpropanoid metabolism in Arachis hypogaea hairy roots. PeerJ, 2021, 9: e10976.
doi: 10.7717/peerj.10976 |
| [4] | 苏良辰. AhHDA1在花生响应干旱和ABA信号中的分子机制研究. 华南师范大学博士学位论文, 广东广州, 2017. |
| Su L C. Study on the Molecular Mechanism of AhHDA1 Responding to Drought and ABA Signal in Arachis hypogaea L. PhD Dissertation of South China Normal University, Guangzhou, Guangdong, China, 2017. (in Chinese with English abstract) | |
| [5] |
李媚娟, 苏良辰, 刘帅, 李晓云, 李玲. 花生AhHDA1互作蛋白AhGLK的筛选及特性分析. 作物学报, 2017, 43: 218-225.
doi: 10.3724/SP.J.1006.2017.00218 |
|
Li M J, Su L C, Liu S, Li X Y, Li L. Screening of AhHDA1 interacting-protein AhGLK and characterization in peanut (Arachis hypogaea L.). Acta Agron Sin, 2017, 43: 218-225. (in Chinese with English abstract)
doi: 10.3724/SP.J.1006.2017.00218 |
|
| [6] |
Liu X, Li L, Li M, Su L, Lian S, Zhang B, Li X, Ge K, Li L. AhGLK1 affects chlorophyll biosynthesis and photosynthesis in peanut leaves during recovery from drought. Sci Rep, 2018, 8: 2250.
doi: 10.1038/s41598-018-20542-7 |
| [7] |
Liu X, Li L, Zhang B, Zeng L, Li L. AhHDA1-mediated AhGLK1 promoted chlorophyll synthesis and photosynthesis regulates recovery growth of peanut leaves after water stress. Plant Sci, 2020, 294: 110461.
doi: 10.1016/j.plantsci.2020.110461 |
| [8] | 蔡大伟. 基于ChIP-seq全基因组识别毛竹笋尖与鞭笋尖组蛋白修饰位点. 福建农林大学硕士学位论文, 福建福州, 2017. |
| Cai D W. Genome-wide Mapping of Histone Modifications of Shoot Tips and Rhizome Shoot Tips in Moso bamboo by ChIP-seq. MS Thesis of FuJian Agriculture and Forestry University, Fuzhou, Fujian, China, 2017. (in Chinese with English abstract) | |
| [9] |
Lismer A, Lambrot R, Lafleur C, Dumeaux V, Kimmins S. ChIP-seq protocol for sperm cells and embryos to assess environmental impacts and epigenetic inheritance. STAR Protoc, 2021, 2: 100602.
doi: 10.1016/j.xpro.2021.100602 |
| [10] |
Liu S, Li M, Su L, Ge K, Li L, Li X, Liu X, Li L. Negative feedback regulation of ABA biosynthesis in peanut (Arachis hypogaea): a transcription factor complex inhibits AhNCED1 expression during water stress. Sci Rep, 2016, 6: 37943.
doi: 10.1038/srep37943 |
| [11] |
Trapnell C, Williams B A, Pertea G, Mortazavi A, Kwan G, van Baren M J, Salzberg S L, Wold B J, Pachter L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol, 2010, 28: 511-515.
doi: 10.1038/nbt.1621 pmid: 20436464 |
| [12] | Emes M J, Tetlow I J, Bowsher C G. Integration of metabolism within non-photosynthetic plastids, and with the cytosol. Regul Primary Metabolic Pathways Plants, 1999, 42: 117-136. |
| [13] | 苏良辰, 钟钰婷, 李玲. AhHDA1对花生毛状根干旱的生理调节作用. 植物生理学报, 2017, 53: 1893-1900. |
| Su L C, Zhong Y T, Li L. Physiological regulation effect of AhHDA1 on peanut hairy roots to drought stress. PlantPhysiol J, 2017, 53: 1893-1900. (in Chinese with English abstract) | |
| [14] |
Waters M T, Wang P, Korkaric M, Capper R G, Saunders N J, Langdale J A. GLK transcription factors coordinate expression of the photosynthetic apparatus in Arabidopsis. Plant Cell, 2009, 21: 1109-1128.
doi: 10.1105/tpc.108.065250 pmid: 19376934 |
| [15] |
Savitch L V, Subramaniam R, Allard G C, Singh J. The GLK1 ‘regulon’ encodes disease defense related proteins and confers resistance to Fusarium graminearum in Arabidopsis. BiochemBiophys Res Commun, 2007, 359: 234-238.
doi: 10.1016/j.bbrc.2007.05.084 |
| [16] |
Zubo Y O, Blakley I C, Franco-Zorrilla J M, Yamburenko M V, Solano R, Kieber J J, Loraine A E, Schaller G E. Coordination of chloroplast development through the action of the GNC and GLK transcription factor families. Plant Physiol, 2018, 178: 130-147.
doi: 10.1104/pp.18.00414 pmid: 30002259 |
| [1] | 张胜忠, 胡晓辉, 慈敦伟, 杨伟强, 王菲菲, 邱俊兰, 张天雨, 钟文, 于豪諒, 孙冬平, 邵战功, 苗华荣, 陈静. 基于三维模型重构的花生网纹厚度性状QTL分析[J]. 作物学报, 2022, 48(8): 1894-1904. |
| [2] | 白冬梅, 薛云云, 黄莉, 淮东欣, 田跃霞, 王鹏冬, 张鑫, 张蕙琪, 李娜, 姜慧芳, 廖伯寿. 不同花生品种芽期耐寒性鉴定及评价指标筛选[J]. 作物学报, 2022, 48(8): 2066-2079. |
| [3] | 徐扬, 张智猛, 丁红, 秦斐斐, 张冠初, 戴良香. 钙肥对酸性红壤花生种子萌发及种子际微生物菌群结构的调控[J]. 作物学报, 2022, 48(8): 2088-2099. |
| [4] | 杨欢, 周颖, 陈平, 杜青, 郑本川, 蒲甜, 温晶, 杨文钰, 雍太文. 玉米-豆科作物带状间套作对养分吸收利用及产量优势的影响[J]. 作物学报, 2022, 48(6): 1476-1487. |
| [5] | 李海芬, 魏浩, 温世杰, 鲁清, 刘浩, 李少雄, 洪彦彬, 陈小平, 梁炫强. 花生电压依赖性阴离子通道基因(AhVDAC)的克隆及在果针向地性反应中表达分析[J]. 作物学报, 2022, 48(6): 1558-1565. |
| [6] | 刘嘉欣, 兰玉, 徐倩玉, 李红叶, 周新宇, 赵璇, 甘毅, 刘宏波, 郑月萍, 詹仪花, 张刚, 郑志富. 耐三唑并嘧啶类除草剂花生种质创制与鉴定[J]. 作物学报, 2022, 48(4): 1027-1034. |
| [7] | 丁红, 徐扬, 张冠初, 秦斐斐, 戴良香, 张智猛. 不同生育期干旱与氮肥施用对花生氮素吸收利用的影响[J]. 作物学报, 2022, 48(3): 695-703. |
| [8] | 黄莉, 陈玉宁, 罗怀勇, 周小静, 刘念, 陈伟刚, 雷永, 廖伯寿, 姜慧芳. 花生种子大小相关性状QTL定位研究进展[J]. 作物学报, 2022, 48(2): 280-291. |
| [9] | 王艺璇, 贾浩, 逯杰, 石晓宇, 赵明宇, 高真真, 赵炯超, 褚庆全. 黄淮海农作区花生生产水足迹及耗水结构分析[J]. 作物学报, 2022, 48(12): 3203-3214. |
| [10] | 胡美玲, 薛晓梦, 吴洁, 郅晨阳, 刘念, 陈小平, 王瑾, 晏立英, 王欣, 陈玉宁, 康彦平, 王志慧, 淮东欣, 姜慧芳, 雷永, 廖伯寿. 花生籽仁脂肪和蔗糖含量的胚、细胞质、母体遗传效应分析[J]. 作物学报, 2022, 48(11): 2724-2732. |
| [11] | 王建国, 耿耘, 杨佃卿, 郭峰, 杨莎, 李新国, 唐朝辉, 张佳蕾, 万书波. 单粒精播对中、高产旱地花生群体质量及养分利用的影响[J]. 作物学报, 2022, 48(11): 2866-2878. |
| [12] | 孙棋棋, 郑永美, 于天一, 吴月, 杨吉顺, 吴正锋, 吴菊香, 李尚霞. 施氮对不同结瘤特性花生土壤固氮菌多样性和群落组成的影响[J]. 作物学报, 2022, 48(10): 2575-2587. |
| [13] | 汪颖, 高芳, 刘兆新, 赵继浩, 赖华江, 潘小怡, 毕晨, 李向东, 杨东清. 利用WGCNA鉴定花生主茎生长基因共表达模块[J]. 作物学报, 2021, 47(9): 1639-1653. |
| [14] | 王建国, 张佳蕾, 郭峰, 唐朝辉, 杨莎, 彭振英, 孟静静, 崔利, 李新国, 万书波. 钙与氮肥互作对花生干物质和氮素积累分配及产量的影响[J]. 作物学报, 2021, 47(9): 1666-1679. |
| [15] | 石磊, 苗利娟, 黄冰艳, 高伟, 张忠信, 齐飞艳, 刘娟, 董文召, 张新友. 花生AhFAD2-1基因启动子及5'-UTR内含子功能验证及其低温胁迫应答[J]. 作物学报, 2021, 47(9): 1703-1711. |
|
||