

以杂交棉石杂2号和新陆早43的F1、F2代及亲本NT2、H2、4-14为试材, 通过测定不同生育时期各材料叶面积指数(LAI)、叶倾角(MTA)、冠层光截获率等指标, 分析了各指标变化对群体光合物质生产的影响。结果表明, 2个杂交棉F1代LAI具有超亲优势, 冠层光截获率具有中亲优势; LAI和冠层光截获率具有明显的母系遗传特性, 而MTA受到父本的显著影响。F2代冠层结构主要受F1代相关指标和衰退率的影响, LAI中亲优势减小了衰退率; 杂交棉F1代光合物质积累主要受亲本参数和超亲优势的影响, F2代主要受F1代参数的影响。杂交棉光合物质积累最大增长速率和直线增长期开始时间较晚, 直线增长期及活跃增长期较长, 最终积累量和最大增长速率较高。杂交棉F1代具有明显的光合生产和产量优势, F2代具有一定的产量优势。以选择具有优化冠层结构的亲本为基础, 组配具有较大MTA的父本和较大LAI的母本, 有利于改善杂交棉光合性能, 提高群体光能利用率, 进一步挖掘产量潜力, 为杂交棉高光效组合的选育及提高F2代应用提供理论依据。
A field experiment was conducted using the F1, F2 of two cotton hybrids of Shiza 2 and Xinluzao 43 and their parents NT2, H2, and 4-14. After measuring the leaf area index (LAI), leaf inclination angle (MTA) and light interception rate (LIR) at different growing periods, it is clear that the matter production of hybrid cotton was affected directly by the parameter changes of the canopy structure and photosynthetic characteristics of two cotton hybrids parents. The results revealed that the over-parent heterosis of F1 in two cotton hybrids was observed on LAI and mid-parent heterosis of them was provided with LIR. Both LAI and LIR were observed obviously with matrilineal genetic characteristics, while MTA was influenced by paternal inheritance. The canopy structure of F2in two cotton hybrids was mainly influenced by the related parameters of F1 in hybrid cotton indexes and their decreasing rate. The decreasing rate was deduced by the over-parent heterosis on LAI. The photosynthate accumulation of F1 in two cotton hybrids was mainly influenced by the numerical value of their parents and over-parent heterosis, and the photosynthate accumulation of F2 in two cotton hybrids was mainly influenced by the related parameters of F1. The initial time of the highest increasing velocity of photosynthate accumulation and the linear increasing period was later, the linear increasing period and the active increasing period of photosynthate accumulation were longer, the total photosynthate accumulation and the highest increasing velocity were larger in two cotton hybrids. The photosynthate and yield were very high in F1 of hybrid cotton, and there was a high yield only in F2 of hybrid cotton. Based on choosing the parents with the optimized canopy structure, we should make a well combination between father plant with larger MTA and mother plant with larger LAI, which will be provides used for improving hybrid cotton photosynthetic performance, increasing light use efficiency for enhancing yield potential further. The study provided come theoretical parameters for breeding high photosynthetic efficiency hybrid line and improving utilization of F2 in hybrid cotton.
作物光合作用是产量形成的基础, 提高作物光合生产力是农学家长期探索的重要研究内容[1,2,3,4]。作物冠层结构显著影响群体光合速率[5]; 改善作物冠层结构是提高光合性能最有效的途径之一[6]。叶面积指数是衡量作物冠层结构的重要指标, 研究表明, 叶面积指数可估算作物冠层潜在光合生产力及作物干物质积累量[7]; 叶倾角的变化能显著影响冠层固定CO2的能力, 进而影响产量[8]。Stewart等[9]通过对大量玉米品种的研究表明, 叶面积和叶倾角是影响光合作用的主要因子; 作物群体光截获率对光合作用有重要作用[10], 改善作物冠层结构特征, 对增强群体光截获率, 提高群体生产力具有重要意义。育种学家已通过遗传改良手段, 选育出具有适宜 于光合作用的最佳叶面积和叶片角度的优良品 种[11,12,13]。棉花杂种优势利用是提高产量和改善品质的重要途径。近年来, 中国在棉花杂种优势的研究和利用方面取得了较大进展, 杂交棉F1代平均增产15%左右[14], 推动了棉花产量水平的提高[15]。然而, 如何通过亲本选配, 进一步发挥杂交种的光合生产优势, 挖掘其增产潜力已成为棉花等作物超高产育种和栽培研究关注的热点[16,17,18,19]。
新疆是我国最重要的商品棉生产基地, 自2001年以来, 新疆引进和选育了多个棉花杂交种并在生产上进行示范推广, 使棉花单产水平有了进一步提高[20,21]。棉花杂交种主要采用人工去雄制种, 成本高且繁殖系数低, 而新疆采用直播, 用种量大, 种植杂交种F1成本过高, 严重制约了高产杂交棉的大面积推广; 棉花是异源四倍体, F2代分离并不严重, 仍保持一定的杂种优势[22,23], 具有较好的增产潜力和广泛的适应性[24], 有一定的生产应用价值。目前已选配的一些杂交组合F2在新疆棉区试验示范, 表现出明显的产量优势, 探索推广种植杂交棉F2已受到棉花科技工作者的重视。因此从遗传角度, 选育能较多保留杂种优势的F2代是迅速大面积推广杂交棉超高产种植亟待解决的问题之一。本研究探讨亲本对杂交棉F1、F2代冠层结构特性的影响, 揭示杂交棉高光效冠层结构形成机制及其与物质生产之间的关系, 以期为超高产杂交棉亲本选配及高产栽培提供理论依据。
选用新疆棉区种植面积较大的棉花杂交种石杂2号和新陆早43的F1、F2代, 石杂2号父本NT2、母本H2, 新陆早43父本H2, 母本4-14; 同时选取常规棉品种新陆早33为对照(用于计算竞争优势), 共8个材料。
试验于2009—2010年在石河子大学农学试验站进行(45°19′ N, 86°03′ E)。采用宽膜覆盖栽培, 先铺膜后点播, 1膜4行, 行距为(60+20+40+20) cm宽窄行; 小区面积为43.5 m2, 随机排列, 重复3次; 留苗密度为每公顷16.5万株。播前深施高效有机肥1500 kg hm-2, 尿素240 kg hm-2, 三料磷肥75.3 kg hm-2作基肥。生育期滴灌12次, 滴水量为3600~4200 m3 hm-2, 8月底停水, 随水滴施尿素260 kg hm-2。用缩节胺化调6次, 用量为300~350 g hm-2。2009年4月19日播种, 4月26日出苗, 7月5日打顶; 2010年5月1日播种, 5月7日出苗, 7月10日打顶。其他管理措施同大田超高产棉花管理。
选择棉花生长发育的关键生育时期, 包括盛蕾期、盛花期、盛铃前期、盛铃后期、吐絮期测定冠层结构、群体光合生产等指标。
1.3.1 冠层结构指标 采用LAI-2000冠层仪(Li-Cor, USA), 参照Malone等的方法[25]测定叶面积指数和叶倾角, 先将探头水平放置于冠层上方, 按下测定按钮, 两声蜂鸣后将探头放入群体内, 仍保持水平, 按下测定按钮, 两声蜂鸣后水平均匀移动探头, 选择冠层内不同位置测量, 重复4次。
1.3.2 光截获率 采用高亮之等[26]介绍的方法, 在北京时间11:00~14:00, 用LI-250A光量子仪(Li-Cor, USA)测定植株顶部以上30 cm处自然光强Io (探头面水平向上)、植株反光In (探头面水平向下)、入射到冠层底部的光强I。反射率(LRR, %)= In/Io×100, 透射率(LLR, %)=I/Io×100, 光截获率(LIR, %)=100-LRR-LLR。
1.3.3 光合物质积累与产量 在各小区选择代表性植株5株, 从子叶节处剪取地上部分, 分解成叶片、茎和蕾铃等器官, 分别装入纸袋, 105℃下杀青30 min, 80℃下烘干后称重。产量因子及计产方法为: 于收获期各处理在各小区选有代表性植株10株, 考察不同果枝单铃重, 并折算各个处理单铃重; 每处理随机选取3个点, 每个取样点面积2.8 m2, 调查各样点全部株数和铃数, 折算出单株结铃数和单位面积总铃数; 以实收籽棉计算产量。
采用中亲优势、超亲优势和衰退率、竞争优势来分析2个杂交棉的杂种优势及遗传表现。中亲优势(%)=(F-P)/P×100, 超亲优势(%)=(F1-PH)/PH×100, 竞争优势(%)=(F-CK)/CK×100, 自交衰退ID2(%)= (F1-F2)/F1×100; 其中F1为杂交1代, F2为杂交2代, P为双亲平均值, PH为高亲值, CK为推广的标准对照品种新陆早33; 采用CurveExpert 1.3对光合物质积累情况进行Logistic方程拟合, 用Microsoft Excel 2003和SPSS 11.0分析处理试验数据, 用SigmaPlot 10.0作图。
表1可见, 2年试验中, 2个杂交棉F1具有明显的超亲优势和竞争优势, F2代具有较强的竞争优势; 石杂2号F1代和新陆早43 F1代皮棉产量均实现3000 kg hm-2以上的高产水平, F1代超亲优势达39.1%~46.3%, 竞争优势为26.4%~45.9%, F2代产量虽然衰退9.2%~12.8%, 但仍具有14.9%~27.4%的竞争优势。分析产量构成因子可以看出, 杂交棉F1代由于单株铃数、铃重及衣分具有不同程度的超亲优势, 产量大幅度提高, 单株铃数超亲优势为13.4%~ 15.3%, 单铃重8.3%~12.6%, 衣分8.2%~12.8%。可见F1代杂种优势是各产量构成因子具有超亲优势 的综合结果; F2代单铃重和衣分与F1无明显差异, 但单株铃数衰退了9.6%~12.1%, 限制了产量水平的提高。
叶面积指数(LAI)的大小直接影响作物对光能的截获率, 进而影响群体光合生产。试验表明(图1), 在测定生育时期内, 2个杂交棉F1代LAI均表现出明显的超亲优势, 以盛蕾期、盛铃后期和吐絮期尤为明显; 新陆早43的F1代LAI明显高于石杂2号F1代。回归分析表明, 母本对杂交种F1代LAI的直接作用更大, 其中石杂2号的直接通径系数为0.7622, 新陆早43为0.5787。F2代出现一定程度的衰退, 但在盛铃后期和吐絮期表现出中亲优势。在相应生育时期内, 新陆早43亲本较石杂2号亲本平均值高8.9%~37.8%, 石杂2号F1代超亲优势在盛蕾期为21.2%~23.9%, 盛铃后期和吐絮期为28.0%~36.5%; 新陆早43 F1代超亲优势盛蕾期为27.4%~33.9%, 盛铃后期和吐絮期为31.1%~64.9%。F2代盛蕾期到盛花期衰退32.6%~75.9%, 盛铃前期到吐絮期衰退7.7%~19.5%; 进一步分析表明, 盛蕾期到盛铃前期F2代无明显中亲优势, 盛铃后期和吐絮期中亲优势石杂2号F2代为18.2%~32.1%, 新陆早43 F2代为29.0%~60.6%。可见杂交棉F1代超亲优势和亲本值越大, 叶面积指数越高; F2代生育后期中亲优势越大, 衰退率越低。
| 表1 杂交棉亲本及F1、F2代产量及产量构成因素(2009-2010)Table 1 Yield and components of hybrid cottons’ parents and their F1, F2(2009-2010) |
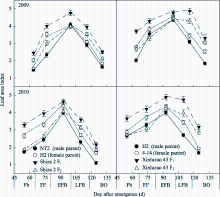 | 图1 杂交棉亲本及F1、F2代叶面积指数的生育时期变化Fb: 盛蕾期; FF: 盛花期; EFB: 盛铃前期; LFB: 盛铃后期; BO: 吐絮期。Fig. 1 Changes of leaf area index of hybrid cottons’ parents and their F1, F2 at different growth stagesFb: full budding; FF: full flowering; EFB: early full bolling; LFB: later full bolling; BO: boll opening. |
作物叶片的空间状态在很大程度上影响光能的利用。试验表明(图2), 在测定生育时期内, 新陆早43 F1代的叶倾角(MTA)高于石杂2号F1代, 2个杂交棉F1、F2均未表现出超亲优势或中亲优势。回归分析表明, 父本对杂交棉F1代MTA的大小起主要作用, 其中石杂2号的直接通径系数为0.6091, 新陆早43为0.9051。F2代衰退率表现为石杂2号8.9%~19.7%, 新陆早43为7.2%~15.2%。父本MTA大小直接影响杂交棉F1代MTA参数高低, F1代参数大小及遗传衰退率的高低决定F2代MTA的高低。
作物光合作用及干物质生产能力与冠层光截获状况密切相关[27]。研究表明(图3), 在测定生育时期内, 新陆早43 F1代冠层光截获率高于石杂2号F1代, 2个杂交棉F1表现出一定的中亲优势; F2代仅在盛铃前期表现出一定的中亲优势, 盛蕾期和吐絮期衰退较为明显。回归分析表明, 石杂2号母本对F1代冠层光截获率直接通径系数为0.6698, 新陆早43为0.7372。杂交棉F1代在盛蕾期和吐絮期中亲优势为3.6%~10.7%, F2代在盛铃前期中亲优势为0.7%~ 1.8%。F2代盛蕾期和吐絮期衰退7.8%~14.6%。可见, 亲本选配主要改善杂交棉F1代生育早期和后期的冠层光截获率, F2代在生育前期和后期较高的衰退率直接影响了光合作用以及干物质生产能力。
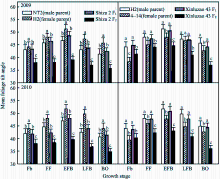 | 图2 杂交棉亲本及F1、F2代叶倾角的生育时期变化同一生育时期标以不同字母的标值在0.05 水平上差异显著。Fig. 2 Changes of mean foliage tilt angle of hybrid cottons’ parents and their F1, F2 at different growth stagesBoxes with superscripts of different letters are significantly different at the 0.05 probability level within a growth stage in the same year. |
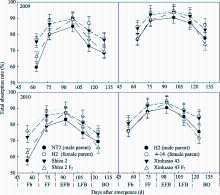 | 图3 杂交棉亲本及F1、F2代冠层光截获率的生育时期变化Fig. 3 Changes of canopy light interception rate of hybrid cottons’ parents and their F1, F2 at different growth stages |
表2表明, 杂交棉F1代总光合物质积累量为 23 916.3~30 442.5 kg hm-2, 超亲优势达47.7%~ 58.9%; F2代有15.7%~30.9%的衰退率。Logistic方程对杂交棉F1代及其亲本光合物质积累过程拟合表明, 杂交棉F1代较亲本光合物质积累增长直线期长6.6~8.8 d, 物质积累活跃期(P)长14.9~20.0 d; 最大增长速率高25.9%~63.2%; F2代较杂交棉光合物质积累增长直线期和物质积累活跃期(P)衰退9.3%~ 11.4%, 最大增长速率下降5.6%~38.8%。进一步分析表明, 亲本直线增长期开始和结束时间及最大速率出现时间较早, F2代直线增长期和最大增长速率出现时间明显晚于亲本, 但直线增长期结束时间早于杂交棉F1代; 新陆早43 F1代和石杂2号F1代相比, 直线增长期开始和结束时间及最大增长速率出现时间均较晚, 直线增长期和物质积累活跃期较长, 最大增长速率较大。杂交棉F2较双亲均值光合物质积累增长直线期长2.2~2.9 d, 物质积累活跃期(P)长5.1~6.5 d; 最大增长速率高7.5%~20.9%; 最大速率出现的时间晚0.6~5.1 d; 总光合物质积累量高14.5%~28.7%。
通过遗传改良手段, 可选育具有适宜光合作用的冠层结构, 如叶面积和叶片角度等[11], 在选育良种的过程中, 选取有效光合性能指标早已受到人们的广泛关注[28]。杂交种性状受其选配的亲本显著影响[14,17,19,29], 在生长势上表现出非常明显的杂种优势[30]。本研究表明, 超亲优势显著影响杂交棉F1代的LAI, 冠层光截获率则受中亲优势影响; MTA并未表现出中亲优势。回归分析表明, LAI和冠层光截获率受母本影响较大, 具有明显的母系遗传特性, 而MTA显著受到父本的影响。杂交种的基因来自双亲, 基因表达方面存在的差异是导致杂种优势的重要原因[31,32]; 基因差异表达是构成F1杂种优势性状差异的分子基础[33,34]; 表型性状的差异可以从基因表达模式上得以反映[35]。有研究表明亲本基因差异表达与F1杂种优势呈正相关[14,35,36,37,38]。2个杂交棉F1代LAI、MTA和冠层光截获率等冠层结构指标杂种优势与亲本差异表达呈正相关。可见, 选择较大MTA和LAI及光合功能较强的父本并配以具有较高光合特性的母本, 能有效改善杂交棉F1代冠层结构, 提高光合功能。本研究表明, 杂交棉F1代光合物质积累量没有明显表现出母系或父系遗传, 这可能与本试验选用材料有关, 但也可能说明光合物质积累是一个数量遗传性状, 受到冠层结构各个指标的影响; 但试验结果表明杂交棉F1代光合物质积累量与亲本平均值和超亲优势呈显著正相关, 因此在选 用配合力较好的亲本基础上组配生长势较强的亲 本是提高杂交棉F1代光合物质积累能力的一个有效途径。
| 表2 杂交棉亲本及F1、F2代光合物质积累特征Table 2 Photosynthate accumulation characteristics parents and F1, F2 of cotton hybrids |
杂交棉F2代在某些性状上表现出中亲优势[39], 有可能出现光合速率高于F1代材料[40]。本试验结果表明, F2代个别指标具有中亲优势, 冠层结构指标和光合特性指标较F1均出现不同程度的衰退。LAI有一定的中亲优势, 减小了F2代衰退率; 冠层光截获率在盛铃前期表现出中亲优势, 未对衰退率产生影响。研究表明F2代部分指标具有的中亲优势起到减轻衰退程度的作用, 冠层优化程度主要取决于F1代的相关指标和遗传衰退率。可见, 亲本对F2代LAI衰退程度具有明显的影响, 并且通过影响F1代相关指标间接影响F2代。杂交棉F2代生育后期光合物质积累具有较明显的中亲优势, 使光合物质积累量明显高于亲本, 因此产量较双亲均值提高26.5%~ 33.7%, 具有一定的杂种优势。研究表明, F2代光合物质积累量与衰退率无关, 主要受F1代相关指标影响。F2代产量较推广的标准对照品种新陆早33高14.9%~27.4%, 尤其是新陆早43 F2, 2年试验结果皮棉产量均实现了3000 kg hm-2以上的高产水平, 2009年较石杂2号F1低0.9%, 2010年较石杂2号F1高0.6%, 与石杂2号F1相差不大, 具有较大的生产应用价值。因此亲本选育很大程度上决定了F2代的竞争优势, 影响其生产应用价值。
作物产量是一个综合性状, 受植株多个形态、生理以及农艺性状的影响[41]。作物高产要求具有较高群体光合生产[40], 通过对冠层结构的改造, 增加光能利用率, 制造更多的光合产物, 增大光合物质积累。杂交棉具有较高的LAI, 能截获更多光能, 光合物质积累增长直线期和活跃期较长、最大速率较大及最终光合物质积累较多, 直线增长期开始和最大速率结束时间及出现时间较晚, 最终光合物质积累较多, 有利于产量提高。因此通过选配亲本优化杂交棉冠层, 提高光合性能, 改善群体光合生产是提高产量的有效途径。冠层形态与结构决定于品种, 要选育冠层结构合理的杂交种, 最大限度地提高光能利用率, 就必须正确选配亲本。在选配亲本过程中不仅要考虑生理功能[42], 而且要重视亲本冠层特性。超亲优势、中亲优势及F2代衰退率对挖掘产量潜力具有重要影响, 而如何改变及设计其参数有待进一步研究。
杂交棉F1冠层优化和光合物质生产力的提高是以亲本为基础, F2代冠层特性主要受杂交棉F1代相关指标和衰退率的影响, 部分指标的中亲优势影响衰退率; 杂交棉F1代光合物质积累主要受亲本参数和超亲优势的影响; 杂交棉光合物质积累最大增大速率出现时间和直线增长期开始时间较晚, 最终积累量和最大增大速率较大以及直线增长期和活跃期较长; 杂交棉F1代具有明显的光合生产和产量优势, F2代具有一定的产量竞争优势。选择组配具有较大MTA的父本、较大LAI的母本, 有利于杂交棉F1代提高光能利用率, 进一步挖掘产量潜力以及提高F2代的应用价值。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|