{kind=link}
{kind=link}
外源IAA、GA3和ABA影响不同穗型小麦分蘖发生的机制
引用本文
蔡铁, 徐海成, 尹燕枰, 杨卫兵, 彭佃亮, 倪英丽, 徐彩龙杨东清, 王振林. 外源IAA、GA3和ABA影响不同穗型小麦分蘖发生的机制 . 作物学报, 2013, 39(10): 1835-1842
[CAI Tie, XU Hai-Cheng, YIN Yan-Ping, YANG Wei-Bing, PENG Dian-Liang, NI Ying-Li, XU Cai-Long, YANG Dong-Qing, WANG Zhen-Lin. Mechanisms of Tiller Occurrence Affected by Exogenous IAA, GA3, and ABA in Wheat with Different Spike-types . Acta Agronomica Sinica, 2013, 39(10): 1835-1842]
Permissions
[CAI Tie, XU Hai-Cheng, YIN Yan-Ping, YANG Wei-Bing, PENG Dian-Liang, NI Ying-Li, XU Cai-Long, YANG Dong-Qing, WANG Zhen-Lin. Mechanisms of Tiller Occurrence Affected by Exogenous IAA, GA3, and ABA in Wheat with Different Spike-types . Acta Agronomica Sinica, 2013, 39(10): 1835-1842]
Copyright©2013, Editorial office of Acta Agronomica Sinica
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
外源IAA、GA3和ABA影响不同穗型小麦分蘖发生的机制
摘要
以山农15和山农8355为材料, 研究外源生长素(IAA)、赤霉素(GA3)、脱落酸(ABA)对小麦分蘖发生的影响及其作用方式。结果显示, 外源IAA和GA3可抑制分蘖的发生, 而外源ABA则仅减缓了分蘖发生速率。进一步分析外源激素对分蘖芽生长动态及分蘖节中内源激素变化的影响, 发现外源IAA和GA3完全抑制分蘖芽的生长, 而外源ABA明显减缓了分蘖芽的生长速率。外源IAA提高分蘖节中IAA含量和IAA/ZT比, 抑制ZT含量的提高; 外源GA3提高分蘖节中IAA含量、IAA/ZT比和ABA/ZT比, 抑制ZT含量的提高; 外源ABA提高分蘖节中ABA/ZT比, 减缓了ZT含量的升高速率。相关性分析显示, 分蘖芽的生长与ZT呈显著正相关, 与IAA、IAA/ZT比和ABA/ZT比呈显著负相关, 与GA3和ABA呈不显著的负相关。试验结果表明, IAA和ZT在小麦分蘖发生过程中起关键作用, 外源激素主要通过影响内源IAA、ZT含量以及IAA/ZT比和ABA/ZT比来影响分蘖芽的生长, 进而调控小麦分蘖的发生。
关键词:
小麦; 分蘖芽; 植物激素; 调控
Mechanisms of Tiller Occurrence Affected by Exogenous IAA, GA3, and ABA in Wheat with Different Spike-types
Abstract
Two winter wheat cultivars, Shannong 15 and Shannong 8355, were used to study the effects of exogenous indole-3-acetic acid (IAA), gibberellic acid 3 (GA3), and abscisic acid (ABA) on tiller occurrence in wheat and the underlying mechanism. Exogenous IAA and GA3 inhibited occurrence of wheat tiller, while exogenous ABA reduced the rate of tiller occurrence. Furthermore, exogenous IAA and GA3 completely inhibited growth of tiller bud, while exogenous ABA significantly slowed the growth rate of tiller bud. The growth of tiller bud was positively correlated with content of zeatin (ZT), but negatively correlated with IAA content and ratios of IAA/ZT and ABA/ZT. In contrast, the growth of tiller bud had slightly negative correlations with GA3 and ABA contents. These results suggest that IAA and ZT play a key role in regulating tiller occurrence in wheat, and exogenous hormones regulate the growth of tiller bud through changing contents of IAA and ZT and the ratios of IAA/ZT and ABA/ZT in tiller node.
Keyword:
Wheat; Tiller bud; Plant hormones; Regulation
引言
合理的群体结构是提高小麦单产的基础, 分蘖的发生与生长特性直接影响群体的结构与质量。分蘖发生可分为分蘖芽形成和伸长生长两个过程[1], 其中后者对环境条件更为敏感。除遗传因素外, 植物激素在分蘖的发生中起关键作用[2]。在各类植物激素中, 生长素(IAA)和细胞分裂素(CTK)在植物侧芽的生长过程中起重要作用[3,4,5], 其中IAA通过顶端优势抑制分蘖的发生, CTK能解除IAA引起的顶端优势, 促进分蘖的发生, 同时IAA也可抑制CTK对分蘖生长的促进效应[6,7]; 喷施赤霉素(GA3)能显著抑制作物分蘖的发生[8,9,10], GA类激素通过诱发IAA的合成或抑制IAA的分解而间接影响顶端优势[11]; 提高内源脱落酸(ABA)的含量可以导致分蘖死亡[3]。分蘖发生不只受某一激素绝对含量的影响, 还取决于激素间相对含量[4,12]。
玉米[13,14]和水稻[15,16]分蘖发生与植物激素的关系, 以及外源激素的调控作用已有相对系统的研究, 在小麦上仅报道了分蘖发生和生长过程中内源激素动态及其与分蘖数的关系[17,18], 而外源激素对小麦分蘖发生的调控效应少有研究。本试验研究外源激素对不同蘖位分蘖发生率, 分蘖节中内源激素变化和分蘖芽生长动态的影响, 旨在探讨其作用机制, 为定向调控小麦分蘖发生、生长, 构建合理群体结构提供依据。
1 材料与方法
1.1 试验材料与预备试验
选用小麦品种山农15和山农8355, 前者为小穗型品种, 后者为大穗型品种。IAA、GA3和ABA均购自Sigma公司(美国), 纯度>99%。
预备试验以主茎基部第2叶腋内分蘖芽为研究对象, 在其生长过程中于小麦苗叶面分别喷施15、30、45、60、90、120 mg L-1的IAA、GA3和ABA, 清水为对照(外源激素和清水均含吐温-20)。发现外源激素浓度为60 mg L-1时可显著抑制分蘖芽生长, 90、120 mg L-1激素处理与60 mg L-1激素处理无明显差异; 当起始分蘖芽长度大于0.5 cm时, 喷施外源激素对分蘖芽生长无明显抑制作用。从而确定外源激素调控分蘖芽生长的最适浓度为60 mg L-1, 分蘖芽受外源激素调控的临界长度值为0.5 cm左右。
1.2 田间试验
2011年10月8日, 将两品种种植于山东农业大学泰安实验农场, 小区面积为3 m×3 m = 9 m2, 完全随机区组设计。每公顷基本苗, 山农15为120万株, 山农8355为90万株, 行距0.25 m。小麦五至六叶期, 当主茎自基部起的第4叶腋内分蘖芽刚开始伸长生长时, 于17:00左右叶面喷施激素IAA、GA3或ABA, 连喷2 d; 每种激素处理4.5 m2, 喷施浓度均为60 mg L-1, 用量为100 mL m-2。为使激素更好地附着于叶片, 用0.5%吐温-20 (Sigma, V/ V)混合于激素溶液中, 以喷施清水(含吐温-20)为对照, 每处理3次重复。
1.3 人工气候箱试验
将两品种种植于RXZ-500D人工智能气候箱(宁波江南仪器厂, 宁波市)内, 箱内环境模拟田间分蘖期的自然气候(图1)。在小麦四叶期, 当主茎基部第2叶腋内分蘖芽刚开始伸长生长时, 在同一天7:00和19:00两次叶面喷施IAA、GA3或ABA与0.5%吐温-20 ( V/ V)的混合液, 以喷施清水(含吐温-20)为对照。激素浓度均为60 mg L-1, 每株喷施1 mL。每处理3次重复。
1.4 测定项目与方法
1.4.1 分蘖发生率 在田间, 于处理后30 d观测小麦各蘖位的分蘖发生率, 以调查蘖位的实际分蘖数占总调查株数的百分比表示。每处理观测30株, 10株为一组取平均值。主茎记为0; 主茎上发生的分蘖称为一级分蘖, 依次用I, II, III, …表示; 从一级分蘖上长出的分蘖称为二级分蘖, 依次用p, 1, 2, …表示, 如最基部一级分蘖的第1和第2个二级分蘖分别记作I-p和I-1, 基部第2个一级分蘖上的第1和第2个二级分蘖分别记作II-p和II-1; 以此类推。喷施外源激素时期为小麦主茎五至六叶期, 分蘖I和II已正常发生, 分蘖III和I-p的分蘖芽长度已超过外源激素调控的临界值, 外源激素对主茎和分蘖I、II、III和I-p无明显作用, 且各分蘖均能正常发生。因此, 仅调查IV、I-1、II-p和II-1的发生率。
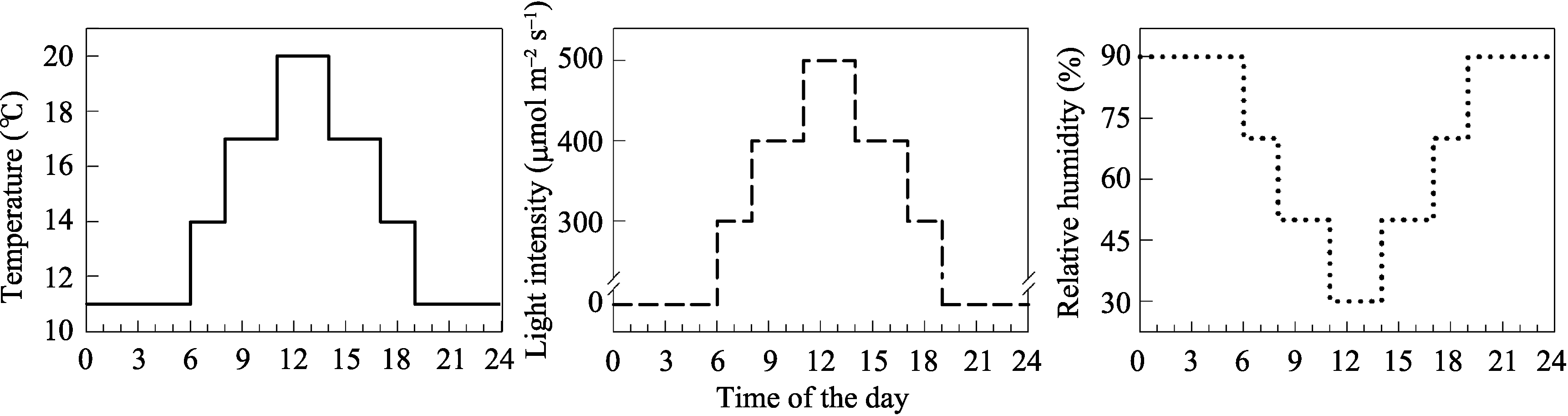 | 图1 人工气候箱内的温度、光照和相对湿度变化Fig. 1 Temperature, light intensity, and relative humidity in phytotron |
1.4.2 分蘖芽长度和鲜重 在人工气候箱试验中, 于处理后0、2、4和6 d分别取主茎基部第2叶腋内分蘖芽, 每处理取30株, 10株为一组, 分别测定其长度和鲜重。
1.4.3 分蘖节中内源激素含量 在人工气候箱试验中, 于处理后0、2、4和6 d分别从单株基部分离分蘖节, 每处理取30株, 10株为一组, 采用高效液相色谱法[19,20]测定其内源激素的含量。称取分蘖节(鲜样) 0.3 g左右, 加入液氮研磨成粉末, 再加入6 mL含有铜试剂(30 μg mL-1)的预冷乙腈, 置4℃冰箱浸提12 h, 随后10 000× g离心15 min, 取上清液。沉淀中再加入4 mL预冷乙腈提取第2次, 第3次加入2 mL提取, 最后合并上清液, 在37~40℃条件下旋转蒸发至干, 用2 mL三氯甲烷和0.4 mol L-1磷酸缓冲液2 mL去除色素, 重复3次, 合并溶液, 取上清液, 加入150 mg PVP除去酯类, 10 000× g离心15 min, 然后在37~40℃条件下旋转蒸发至干, 用1 mL流动相(甲醇︰乙腈︰0.6%乙酸为50︰5︰45, V︰ V︰ V)溶解, 用孔径为0.22 μm的有机滤膜过滤后上机分析。所用分析柱为Symmetry C18 (150 mm×4.6 mm, 5 μm; Waters, 美国)。柱温为25℃, 流动相流速0.5 mL min-1, 检测波长为254 nm。
1.5 数据分析
采用SPSS 18.0软件进行方差分析, 用Duncan’s新复极差(SSR)法进行多重比较。以外源激素处理 4 d后分蘖节中内源激素水平与分蘖芽长度、鲜重 为数据源进行相关分析[21]。用SigmaPlot 10.0软件绘图。
2 结果与分析
2.1 外源激素对小麦分蘖发生的影响
对照的IV、I-1、II-p和II-1分蘖均正常发生; 喷施IAA和GA3显著降低了其发生率, 而喷施ABA仅降低IV和II-1的发生率(表1)。表明喷施IAA和GA3完全抑制小麦分蘖的发生, 而外源ABA则仅抑制部分蘖位分蘖的发生。两品种各蘖位分蘖发生率对外源激素的响应基本一致。
2.2 外源激素对分蘖芽生长的影响
相同处理中两品种分蘖芽生长动态趋势一致(图2)。对照分蘖芽长度和鲜重于处理2 d后急剧增加, 且显著高于其他处理。外源IAA和GA3处理后6 d内分蘖芽一直处于生长停滞状态。外源ABA处理4 d后分蘖芽长度和鲜重缓慢增加, 至6 d时, 其长度和鲜重显著高于同期GA3和IAA处理, 但明显低于对照。说明外源IAA和GA3可抑制分蘖芽生长, 而外源ABA显著降低分蘖芽的生长速率。
2.3 外源激素对分蘖节中内源激素含量的影响
2.3.1 分蘖节中内源激素绝对含量的变化 分蘖芽生长过程中, 分蘖节中内源ZT含量呈明显上升趋势。喷施IAA和GA3显著降低ZT含量。而ABA处理中, 内源ZT含量于处理后缓慢上升, 显著高于同期IAA和GA3处理, 但明显低于对照(表2)。
喷施IAA和GA3后0~4 d, 分蘖节中IAA含量明显升高, 随后降低; 至处理后6 d, 所有处理间无显著性差异; 对照和ABA处理, 分蘖节中IAA含量略有升高或降低, 但无明显规律性(表2)。
| 表1 喷施外源激素对小麦各蘖位的分蘖发生率的影响 Table 1 Percentage of tiller occurrence at different tiller positions in wheat treated by exogenous hormones (%) |
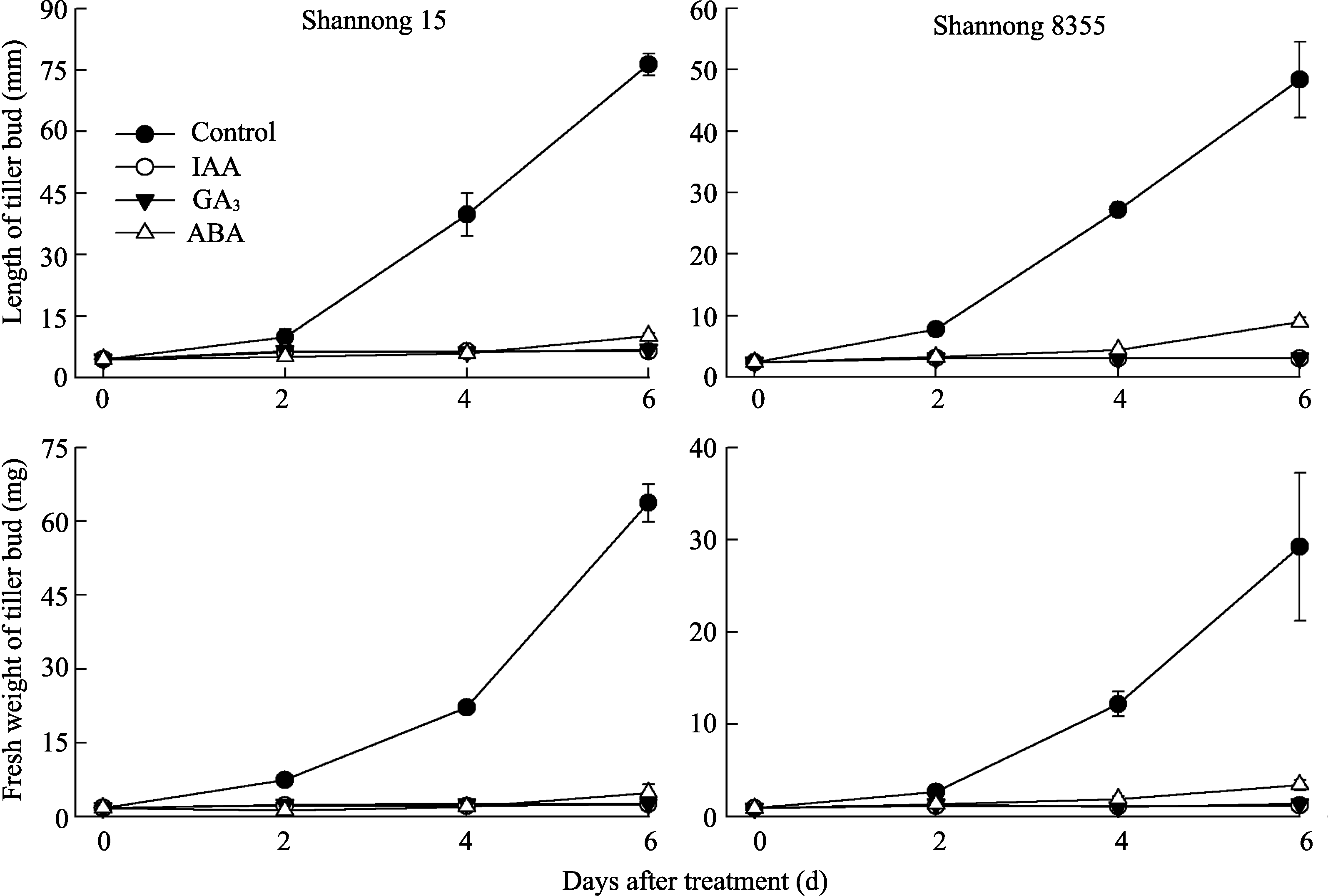 | 图2 外源激素对分蘖芽长度和鲜重的影响数据源自人工气候箱试验。数据为3次重复的平均值±标准差, 每个观测值为10株的平均值。Fig. 2 Effects of exogenous hormones on length and fresh weight of tiller bud |
Control(对照)为叶面喷施含吐温-20的清水; 3种激素的喷施浓度均为60 mg L-1。
Data originate from the phytotron experiments. Data are means ± SD of three replicates, and each measurement was the average of ten plants.
Control indicates water spray with Tween-20 added, and the concentration of all hormones is 60 mg L-1.
在处理后6 d内, 对照及IAA、ABA处理的分蘖节中GA3含量基本保持不变, 且处理间无显著差异; 喷施GA3处理后, 分蘖节中GA3含量呈先升高后降低趋势, 于处理后2 d达到峰值, 后迅速下降, 处理4 d后所有处理间其含量均无显著差异(表2)。
处理后6 d内, 对照和IAA处理中ABA含量基本保持不变; 而ABA和GA3处理中ABA含量均呈先升后降趋势, 其峰值分别出现在处理后2 d和4 d。喷施ABA后分蘖节中ABA含量显著高于其他处理, 两品种趋势一致; 而喷施GA3后ABA含量在山农8355中随时间持续上升, 但其水平未达到外源ABA处理的含量, 在山农15中表现为处理后2 d低于对照, 之后升高, 超过对照和IAA处理(表2)。
2.3.2 分蘖节中激素相对含量的变化 对照和ABA处理中山农15分蘖节中IAA/ZT比基本保持不变, 而山农8355则表现下降趋势, 且处理间无显著差异。IAA和GA3处理中两品种分蘖节内IAA/ZT比变化基本一致, 均为处理后呈先升后降趋势。其中, GA3处理的该比值在处理后2 d达到峰值, 而IAA处理的峰值出现在处理后4 d, 且均明显高于对照和ABA处理(表3)。
分蘖芽生长过程中, ABA/ZT比呈下降趋势, 而IAA处理中其比值基本保持不变, GA3和ABA处理中其比值则呈先升后降趋势。其中, 外源ABA处理后, ABA/ZT比于处理后2 d达到峰值, 而GA3处理中其峰值出现在处理后4 d (表3)。
2.4 分蘖芽生长与内源激素含量的关系
相关性分析显示, 分蘖芽长度和鲜重与ZT呈显著正相关, 与IAA、IAA/ZT比和ABA/ZT比呈显著负相关(表4)。
3 讨论
3.1 外源激素对小麦分蘖发生的影响
除遗传因素[22,23]和营养物质代谢[24]外, 植物激素对作物分蘖发生有重要作用。通过外源激素处理, 可调控分蘖的发生[3,15]。控蘖剂(人工激素类)可抑制水稻分蘖发生, 且调控的目标蘖位与施药时叶龄关系密切[25]。本试验结果与之类似, 由于喷施外源激素时期为小麦主茎五至六叶期, 故对分蘖I、II、III、I-p无促控作用, 而仅影响更高蘖位的分蘖发生。IAA通过顶端优势抑制分蘖发生[6,11], 控制分蘖数目。外源GA3也可以抑制作物分蘖发生[8,9,10]。提高内源ABA含量可导致分蘖衰亡[3], 通过烯效唑干拌种可促进小麦分蘖的发生[24], 喷施乙烯利能增加甘蔗分蘖[26]。本研究显示, 喷施IAA和GA3可完全抑制分蘖的发生, 而喷施ABA则仅降低分蘖IV和II-1的发生率, 抑制部分蘖位分蘖的发生。
| 表2 外源激素对分蘖节中不同激素含量的影响 Table 2 Effect of exogenous hormones on the contents of different hormones in tiller node (ng g-1 FW) |
| 表3 外源激素对分蘖节中激素相对含量的影响 Table 3 Effect of exogenous hormones on the ratio of IAA to ZT in tiller node |
| 表4 内源激素与分蘖芽长度和鲜重的相关性分析 Table 4 Correlation coefficients of hormone contents in tiller nodes with the lengths and fresh weights of tiller buds |
3.2 外源激素对小麦分蘖芽生长的影响
小麦分蘖由分蘖芽发育而来, 植物激素在其伸长生长过程中起着关键作用。IAA通过极性运输抑制侧芽生长[5], 研究表明喷施NAA (人工生长素)可使水稻分蘖芽完全停止生长[17]。本研究中, 喷施IAA后完全抑制了小麦分蘖芽的生长; 外源GA3亦可抑制分蘖芽的生长, 这与Woodward和Marshall的报道一致[27]。据报道, 外源ABA能显著地抑制侧芽生长[28], 小麦分蘖中ABA含量的增加会抑制其进一步发育而引起衰亡[17]。本研究中有不同发现, 外源ABA处理后, 小麦分蘖芽生长并没有受到完全抑制, 仅延缓了分蘖芽生长速率; 这与外源ABA对水稻分蘖芽生长的影响一致[15]。其原因可能与外施激素的部位和时期不同有关。由于分蘖的发生以其第一完全叶生出分蘖鞘为标志, 外源ABA显著降低分蘖芽的生长速率, 推迟其伸出分蘖鞘时间, 从而表现为喷施ABA仅降低部分蘖位分蘖的发生率。从作用效果方面分析, 外源IAA、GA3对分蘖芽的调控效应明显高于外源ABA。
3.3 内源激素与小麦分蘖芽生长的关系
Michel等[29]指出, 外界环境主要通过改变内源激素含量引发生理上的效应进而影响分蘖生长。CTK和IAA在植物侧芽的生长过程中起重要作用[4], 其中, CTK在茎部[30]和根部[31]合成, 运输至叶腋处促进侧芽生长。较高的ZR (细胞分裂素类)可促进玉米分蘖芽萌发, 有利于分蘖发生[13]。外源6-BA(人工细胞分裂素)可提高分蘖节中细胞分裂素, 促进水稻分蘖芽生长[16]。外源细胞分裂素可促进小麦分蘖芽生长[7], 较高的细胞分裂素含量有利于小麦分蘖发生[32]。本试验中, 在分蘖芽生长过程中, 内源ZT呈上升趋势, 而喷施IAA、GA3和ABA抑制分蘖生长同时, 显著抑制内源ZT积累; 相关分析显示, 分蘖芽生长与内源ZT含量呈显著正相关, 表明内源ZT含量提高有利于分蘖芽生长。与CTK作用相反, IAA对侧芽生长起负效应, 其在植物形态学顶端合成, 通过极性运输向下转运至基部抑制侧芽生长[5]。降低IAA的含量可以打破侧芽休眠使其萌发[24]。外源生长素可消除CTK对侧芽生长的促进作用[33,34]。本研究中, 外源IAA和GA3处理后提高了分蘖节中IAA含量, 完全抑制分蘖芽生长; 内源IAA含量与分蘖芽生长呈显著负相关, 说明IAA与分蘖芽生长直接相关。但更多的研究表明, 调控侧芽生长, 虽依赖于IAA和ZT的绝对含量, 但更取决于二者的相对含量[4,12]。研究表明, 较低IAA/ZR比可促进玉米分蘖, 较高IAA/ZR比则抑制分蘖的发生, 促进分蘖的衰亡[13]。刘杨等[15]也报道, 提高IAA/Z+ZR比不利于水稻分蘖芽生长。小麦分蘖发生与IAA和Z+ZR含量相关, 但二者间的比值与分蘖发生关系更为密切, 当IAA/Z+ZR比低时, 利于分蘖发生, 反之则不利于分蘖发生[32]。小麦分蘖能力与内源激素IAA和ZR含量有关, 尤其与两者间比值及其变化趋势密切相关, 在IAA/ZR+Z比下降过程中, 小麦分蘖能力逐渐增加[35]。本研究结果与此相一致, 且外源IAA和GA3显著提高了内源IAA含量, 抑制了内源ZT含量的提高, 进而提高IAA/ZT比, 通过提高IAA与ZT比值, 而完全抑制分蘖芽生长。这可能是外源IAA和GA3调控分蘖芽生长的主要内因, 同时, 也印证了外源NAA和GA3对水稻分蘖芽调控的推 测[15]。
除IAA及CTK类物质外, 关于GA类激素与分蘖的关系也有许多报道, 喷施GA3能显著抑制作物分蘖发生[8,9,10]。张祖德[36]推测外源GA3通过影响植株营养来抑制分蘖芽生长, 但刘杨等[15]认为分蘖芽的生长与植株中碳氮代谢没有必然的关系, 其生长主要受激素代谢调控。本试验通过对内源GA3与分蘖芽长度、鲜重的相关分析, 发现其与分蘖生长无显著相关性。Phillips[11]认为GA类激素通过诱发IAA的合成或抑制IAA的分解而间接影响顶端优势。本研究中, 外源GA3通过提高内源IAA含量, 抑制内源ZT含量的提高, 进而改变IAA/ZT比, 使分蘖芽生长停滞。从而我们推测, 外源GA3通过改变内源IAA与ZT含量的平衡进而影响分蘖发生、生长。此外, 外源GA3除影响内源IAA和ZT含量外, 还显著提高了内源ABA含量, 这与外源GA3对水稻中内源ABA影响类似[15]。之前研究认为, 赤霉素和脱落酸在调节植物生长诸多方面存在相互关系[37], 因此我们推测外源GA3调控分蘖芽生长极可能还与内源ABA有关。
ABA与侧芽休眠关系密切[38], 水稻再生芽中较高的ABA含量是抑制其生长的主要因素[39]。也有研究报道, 小麦分蘖中ABA含量的提高会抑制分蘖进一步发育而导致衰亡[17]。本研究中, 喷施ABA虽提高了内源ABA含量, 但并未完全抑制分蘖芽生长, 仅减缓了其生长速率。Liu等[21]研究表明ABA与水稻分蘖芽生长无显著相关性, 本试验相关性分析结果与此相一致, 表明ABA可能不是调控分蘖芽生长的主要因素。外源ABA对分蘖发生调控可能取决于内源ABA与其他激素间的平衡。许多研究结果显示, 较高的ABA/ZR比可抑制分蘖的发生, 促进分蘖的衰亡[13,24,40]。本研究中喷施GA3和ABA后抑制分蘖芽生长, 但显著提高了ABA/ZT比, 且ABA/ZT比与分蘖芽生长呈显著负相关, 从而推测, ABA主要通过与ZT的平衡来影响分蘖芽的生长。同时, 喷施ABA虽提高ABA/ZT比, 但并未使分蘖芽生长停滞, 说明ABA/ZT比对分蘖芽的作用明显低于IAA/ZT比。关于激素间的互作方式与效应还需进一步研究。
4 结论
小麦分蘖发生受多种激素共同调控, 外源激素通过影响内源激素含量, 以改变内源激素间的相对平衡而作用于分蘖发生。其中, 外源IAA通过提高分蘖节中IAA含量、抑制ZT含量的提高, 以提高IAA/ZT比使分蘖芽生长停滞, 从而抑制分蘖发生; 外源GA3主要通过提高分蘖节中IAA/ZT比和ABA/ZT比, 使分蘖芽生长停滞, 来抑制分蘖发生; 外源ABA通过减缓分蘖节中ZT含量的上升速率, 以提高ABA/ZT比来延缓分蘖芽生长速率, 从而减缓分蘖发生速率。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|