
{kind=link}
不结球白菜维生素C含量主基因+多基因遗传分析
[林婷婷, 王建军, 王立, 陈暄, 侯喜林, 李英*  ]
]
]
|
|


以高维生素C含量不结球白菜自交系乌塌菜和低维生素C含量不结球白菜自交系二青杂交获得的6个世代(P1、P2、F1、B1、B2和F2)株系为材料, 应用植物数量性状主基因+多基因混合遗传模型, 对不结球白菜中维生素C含量进行遗传分析。结果显示, 不结球白菜中维生素C含量受1对加性主基因+加性-显性多基因控制, 其中2011年结果中, 主基因的加性效应为13.15, 在B1、B2、F2世代中主基因的遗传率分别为54.38%、38.58%和18.69%, 多基因的遗传率分别为24.69%、36.92%和40.70%; 2013年结果中, 主基因的加性效应为6.04, 在B1、B2、F2世代中主基因的遗传率分别为1.88%、6.41%和45.04%, 多基因的遗传率分别为39.67%、16.57%和16.91%。可见, 不结球白菜维生素C性状受环境影响较大, 在不结球白菜高维生素C含量品种选育过程中, 要注重环境影响, 并可以通过分子标记辅助选择, 对性状进行改良。
The mixed major gene plus polygene inheritance model was used to investigate the inheritance law of vitamin C in non-heading Chinese cabbage (
随着人们生活水平的提高, 人们对蔬菜产品质量的要求日益提高, 不仅要求外观美、风味佳, 而且要求蔬菜具有丰富、均衡的营养[ 1]。维生素C是一种高活性物质, 能够提高人体免疫力, 预防癌症、心脏病、中风的发生, 也可作为抗衰老及抗逆境的重要生理指标。作为不结球白菜的重要品质性状之一, 维生素C的遗传方式属于数量遗传[ 2]。曾国平和曹寿椿[ 3]采用经典数量遗传方法的基因效应分析初步确定, 不结球白菜中维生素C含量符合加性-显性模型。张增翠等[ 2]研究显示不结球白菜中维生素C含量符合1对主基因+多基因混合遗传模型, 但没有确定具体的遗传模型, 也未确定基因间的效应和不同基因在遗传效应上的差异。近年来, 随着数量遗传分析方法的发展, 植物数量性状主基因+多基因混合遗传模型分析方法[ 4, 5]应运而生, 该方法不仅可以鉴别主基因, 检测多基因效应, 还能检测基因间的交互作用[ 6]和上位性, 并涉及基因与环境间的互作[ 7]。近年来, 植物数量性状的主基因+多基因混合遗传模型分析方法不仅在小麦、玉米、水稻等大田作物中广泛应用[ 8, 9, 10, 11], 也越来越多地被用于辣椒、青花菜、甘蓝、西葫芦等蔬菜作物的研究[ 12, 13, 14, 15, 16]。
本研究利用植物数量性状主基因+多基因多世代联合分析方法, 分析不结球白菜中维生素C含量的遗传, 以期为不结球白菜高维生素C育种和品质改良的世代选择提供理论依据。
从不结球白菜高代自交系中, 选择具有代表性的高维生素C含量的乌塌菜(P1)为母本, 低维生素C含量的二青(P2)为父本, 杂交得到F1, 分别与P1、P2回交得到B1、B2群体, F1自交得到F2群体。全部供试材料由南京农业大学白菜课题组提供。
2011年9月在南京农业大学大棚中种植P1、P2、F1、B1、B2和F2代群体, 其中, P1、P2和F1各18株, F2 178株, B1 59株, B2 58株。2013年9月在南京农业大学江浦农场露地栽培P1 24株、P2 22株、F1 23株、F2 215株、B1 63株和B2 61株。待植株长到第8周时进行试验。
在上午9:00, 取不结球白菜植株同一叶位叶片
的相同部位, 称0.2 g, 用铝箔纸包好立即放入液氮, 于-80℃冰箱保存。参照Bartoli等[ 17]的方法, 并进行改进, 将植物叶片放在研钵中, 加入0.1%的草酸1.5 mL充分研磨; 4℃条件下15 294 × g离心15 min, 取上清液; 在高效液相色谱仪上进样, 测定维生素C含量。色谱以0.1%的乙酸为流动相, 流速1 mL min-1, 柱温30℃, 检测波长为245 nm。
在2011年和2013年的结果中, 亲本P1和P2维生素C含量差异极显著, F1维生素C含量的平均值介于两亲本之间, 未显示超亲优势, 但数值上更偏向于维生素C含量高的P1(表1)。B1、B2和F2群体中维生素C含量次数分布(图1)表明, 不结球白菜中维生素C性状具有主基因-多基因的遗传特征, 可以进行下一步遗传分析。
| 表1 亲本和F1中的维生素C含量次数分布 Table 1 Frequency distribution of vitamin C content in populations of P1, P2, and F1 |
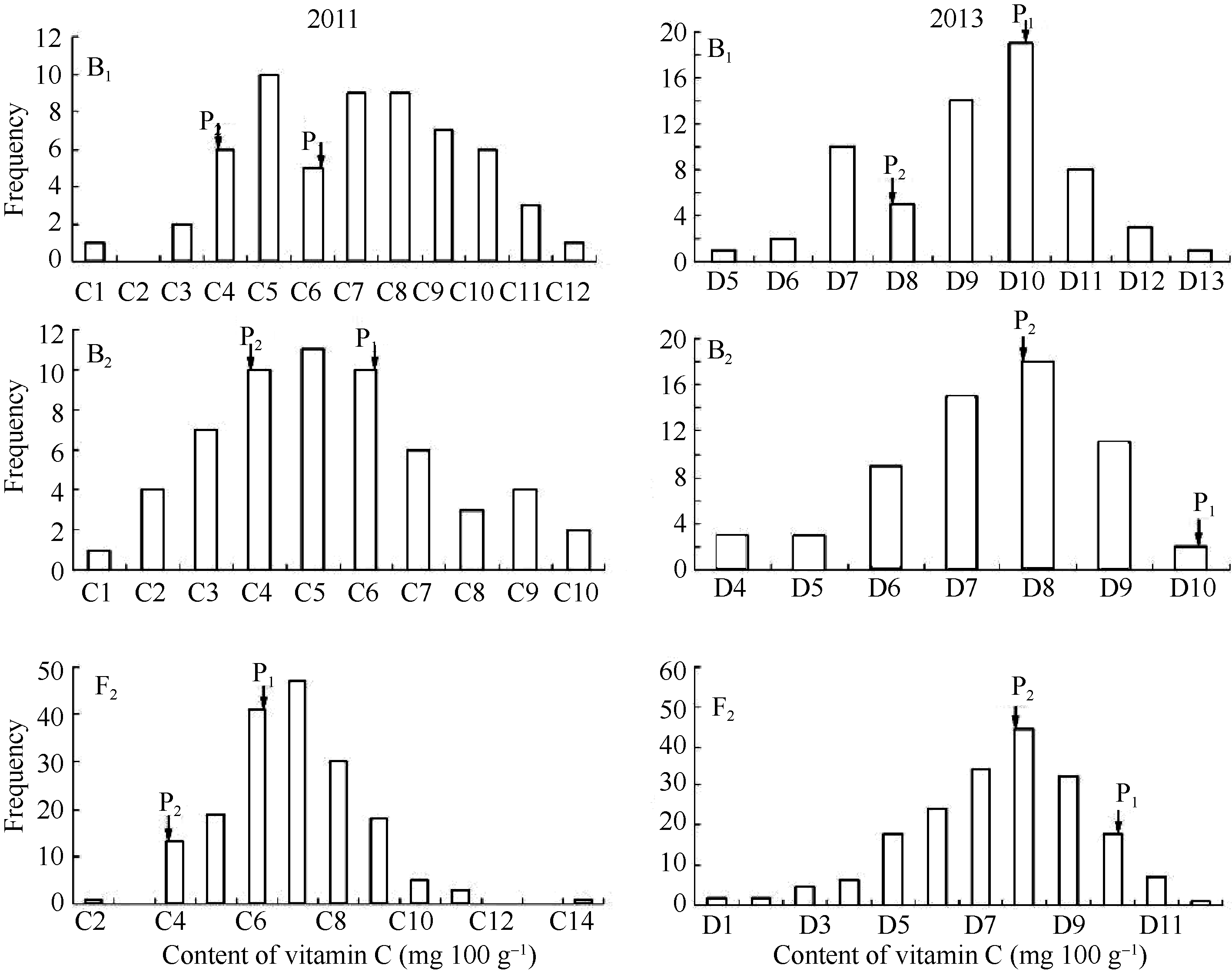 | 图1 B1、B2和F2群体的维生素C含量次数分布Fig. 1 Frequency distribution of vitamin C content in populations of B1, B2, and F2C1: 20-30; C2: 30-40; C3: 40-50; C4: 50-60; C5: 60-70; C6: 70-80; C7: 80-90; C8: 90-100; C9: 100-110; C10: 110-120; C11: 120-130; C12: 130-140; C13: 140-150; C14: 150-160; D1: 37-45; D1:45-53; D3: 53-61; D4: 61-69; D5: 69-77; D6: 77-85; D7: 85-93; D8: 93-101; D9: 101-109; D10: 109-117; D11: 117-125; D12: 125-133; D13:133-141. |
利用植物数量遗传性状主基因+多基因混合遗传模型的多世代联合分析方法, 根据2年对维生素C含量的分析, 分别获得1对主基因(A)、2对主基因(B)、多基因(C)、1对主基因+多基因(D)和2对主基因+多基因(E) 5类24种模型的极大对数似然值和AIC值(表2), 根据AIC值最小原则, 从各遗传模型中初步选备选模型。其中, 2011年大棚条件下, 维生素C性状的备选遗传模型为D-2、D-4和D-1模型; 2013年露地栽培条件下, 维生素C性状的备选遗传模型为D-2、D-3和B-1模型。
对备选模型进行适合性检验, 选择统计量达到显著水平个数最少的模型作为最适遗传模型(表3)。2011年维生素C的备选模型中, D-2、D-4和D-1模型的统计量均未达到显著水平, 2013年维生素C的备选模型中, D-2、D-3和D-1模型的统计量均有一个达到显著水平。结合各模型的AIC值(表2), 比较选择结果表明, 2年的最优遗传模型均是D-2模型, 即1对加性主基因+加性-显性多基因模型。
| 表2 各遗传模型的极大对数似然值和AIC值 Table 2 Estimation of max-likelihood-value and AIC value of different genetic models |
| 表3 备选模型的适合性检验 Table 3 Fitness test of candidate genetic models |
根据模型的极大似然估计值, 按照盖钧镒等[ 4, 5]的方法, 估计模型的一阶遗传参数和二阶遗传参数(表4)。2011年大棚试验中, 控制不结球白菜维生素C含量的主基因加性效应值是13.15, 多基因的加性和显性效应值分别是-1.93和18.33, 而多基因的显性势能比值为-9.51, 表明多基因呈负向完全显性。数据显示, 主基因的加性效应明显高于多基因的加性效应, 而多基因的显性效应明显, 具有负向超亲优势。从效应的绝对值来看, 维生素C含量的显性效应明显高于加性效应。B1、B2、F2世代中的主基因遗传率分别为54.38%、38.58%和18.69%, 多基因遗传率分别为24.69%、36.92%和40.7%, 主基因方差占总遗传方差的比例分别为68.78%、51.10%和31.47%。在F2群体中, 主基因+多基因遗传效率决定了维生素C含量表型变异的59.39%, 其余40.61%的变异由环境因素决定, 表明环境因素对维生素C含量存在较大影响。
2013年露地栽培的调查结果显示, 控制不结球白菜维生素C含量的主基因加性效应值是6.04, 多基因的加性和显性效应值分别是2.31和-2.95, 多基因的显性势能比值为-1.28, 其显性效应大于加性效应, 有负向超显性作用。B1、B2、F2世代中的主基因遗传率分别为1.88%、6.41%和45.04%, 多基因的遗传率分别为39.67%、16.57%和16.91%。在F2群体中, 主基因+多基因遗传效率决定了维生素C含量表型变异的61.95%, 还有38.05%的变异受环境因素影响, 且在B1、B2群体中, 维生素C含量受环境因素的影响更大。这表明在露地环境中, 环境对维生素C含量的遗传存在较大影响。在2年结果中维生素C含量的遗传参数值相差较大, 表明环境对遗传参数有较大影响。F2群体中主基因与多基因的遗传比例变化较大, 但整体遗传效率比较稳定, 在育种中适合在F2群体中进行选择。
| 表4 维生素C含量的D-2模型遗传参数的估计 Table 4 Estimates of genetic parameters of vitamin C in D-2 model |
维生素C含量是不结球白菜的重要品质与农艺性状, 其遗传效应属于数量遗传, 受多基因控制, 遗传机制非常复杂[ 2, 18]。因此对维生素C含量遗传规律的研究就成为不结球白菜优质育种的必要前提, 同时也可以为合理的世代选择提供理论依据。张增翠等[ 2]采用王建康等[ 19]的数量性状遗传模型, 发现不结球白菜还原型维生素C符合1对主基因+多基因的混合遗传模型。本研究采用了盖钧镒等[ 4, 5]的主基因+多基因遗传体系分离分析方法。该方法在王健康等[ 19]遗传模型的基础上, 将混合基因遗传模式扩展到5类24种。本研究结果表明不结球白菜的维生素C含量符合1对加性主基因+加性-显性多基因的遗传模型, 这与张增翠等[ 2]的结果类似。
本研究中, 2011年F2群体的主基因遗传效应小于多基因遗传效应, 遗传率分别为18.69%和40.70%; 而2013年F2群体的主基因和多基因遗传率分别是45.04%和16.91%, 这表明对于不结球白菜维生素C含量性状, F2群体中主基因+多基因的遗传效应占60%左右, 还有很大一部分是受环境因素影响。另外, 2011和2013年的主基因遗传率相差较大, 说明环境因素造成的变异对不结球白菜维生素C含量的影响很大。许伟利等[ 20]对华南型黄瓜维生素C含量的遗传分析中发现, 温室栽培条件下, 环境方差占表型方差的比例为25.78%~63.66%, 说明环境对维生素C含量的遗传有较大影响。而Massot等[ 21]研究发现光照等环境因素对维生素C基因表达的影响以番茄叶片中大于果实中, 因此本研究中不结球白菜叶片中维生素C含量很可能受环境影响较大。另外Ioannidi等[ 22]发现温度会通过诱导番茄中维生素C相关基因的表达, 而影响维生素C的含量。在本研究中, 大棚试验与露地试验的环境条件差别较大, 这也可能是影响不结球白菜维生素C含量遗传的原因。因此, 温度、光照等环境因素的差异, 极有可能造成2年试验中F2世代主基因遗传率相差较大。
在不结球白菜维生素C含量的遗传中, 环境方差占表型方差的比例较高, 说明在维生素C含量的遗传中基因型与环境之间存在较强的互作效应[ 23]。因此在不结球白菜高维生素C含量品质的育种过程中, 应创造良好的栽培条件, 降低环境对维生素C含量的影响。因为环境对维生素C含量影响大, 所以在育种中要加强每代植株维生素C含量的检测, 以确保选择的准确可靠。本研究将植物数量性状遗传的主基因+多基因分离分析体系应用于不结球白菜维生素C含量的遗传分析中, 发现控制不结球白菜维生素C含量的遗传效应属于主基因+多基因效应。随着测序技术和分子标记技术的发展, 在今后的研究中可以利用现有基因组测序数据进行分子标记设计和开发, 通过本研究构建的群体进行分子标记的筛选, 从而获得控制维生素C含量性状主效基因的分子标记, 并将相关基因定位到不结球白菜遗传连锁图谱上。此外, 将主基因+多基因的遗传分析结果与QTL定位数据相结合, 可以进一步验证数量遗传中主效基因的数量和作用, 从而提高遗传分析结果的准确度和可靠性[ 24]。因此, 在不结球白菜维生素C含量性状分子遗传育种中, 除研究其遗传规律外, 下一步还需挖掘主效基因以及相关QTL, 为相关多基因的聚合奠定基础。由于环境因素能够影响数量性状基因的表达, 导致不同环境下遗传分析结果的差异[ 25], 因此需要利用现有6个世代材料构建稳定的不结球白菜重组自交系群体, 通过设计多个环境和多个重复条件, 提高不结球白菜维生素C含量遗传分析结果的精度, 为今后的QTL定位和分子标记开发奠定基础。
不结球白菜的维生素C含量受1对加性主基因+加性-显性多基因控制, 受环境影响较大。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|