

{kind=link}
{kind=link}
{kind=link}
{kind=link}
荞麦茎秆解剖结构和木质素代谢及其与抗倒性的关系
[汪灿1 , 阮仁武1, 2 , 袁晓辉1, 2 , 胡丹1 , 杨浩1 , 林婷婷1 , 李燕1 , 易泽林1, 2, *  ]
]
]
|
|


以抗倒伏能力不同的4个荞麦品种研究表明, 荞麦茎秆抗倒伏能力与茎秆解剖结构和木质素代谢密切相关。倒伏率与茎秆抗折力参数(
Lodging is a major problem in buckwheat (
荞麦是蓼科(Polygonaceae)荞麦属( Fagpyrum)作物, 有苦荞( F. tataricum)和甜荞( F. esculentum) 2个栽培种, 起源于中国, 具有生长期短、适应性强的特点, 是很好的救灾填闲作物和重要的蜜源作物[ 1]。我国是荞麦的生产大国, 总产量居世界第2位, 出口量居世界第1位, 主产区较为集中, 华北、西北、东北地区以甜荞为主, 西南地区的四川、贵州、云南等省以苦荞为主[ 2]。荞麦是一种医食同源的食物, 具有很高的营养、药用及保健品质, 含有丰富的蛋白质、脂肪、淀粉、维生素、芦丁、矿质元素和植物纤维素, 对心血管疾病、糖尿病和便秘等有预防和治疗作用, 已成为21世纪人类的绿色食品之一[ 3]。倒伏是影响荞麦产量的重要因素之一[ 4], 轻者减产, 重者绝收, 极大地制约了我国荞麦生产的发展及农民种植荞麦的积极性。因此, 研究荞麦茎秆抗倒伏规律对实现荞麦高产、稳产具有重要意义。木质素作为细胞壁的主要成分之一, 其含量与茎秆抗倒伏性密切相关[ 5]。木质素含量的增加能提高茎秆的机械强度, 增加其抗压、抗倒伏能力[ 6, 7]; 反之易造成倒伏[ 8]。茎秆的抗倒伏能力与茎秆的解剖学结构有密切联系[ 9, 10]。茎秆机械组织层数越多、机械组织和茎壁越厚、维管束数量越多且维管束面积越大就越能增强茎秆的抗折力和抗倒伏能力[ 11, 12, 13]。目前, 关于茎秆解剖结构和木质素代谢与抗倒伏能力关系方面的研究多数是针对水稻、玉米、小麦等禾本科作物进行的, 抗倒伏能力强的小麦[ 14, 15]、玉米[ 16, 17]和水稻[ 10]品种, 其茎秆基部节间木质素含量高、相关合成酶活性强, 茎秆粗壮、机械组织细胞层数和维管束数目多且维管束面积大, 而在荞麦上尚未有系统的报道。为此, 本研究以抗倒伏能力不同的4个荞麦品种为材料, 研究其茎秆基部第2节间解剖结构、木质素含量及木质素代谢相关酶活性、茎秆抗折力参数和倒伏指数的变化规律, 探讨解剖结构和木质素代谢与茎秆抗倒伏能力的关系, 旨在为荞麦的抗倒伏栽培和品种选育提供理论依据。
西大花荞(高抗倒伏)、乌克兰大粒荞(易倒伏)及信农1号和宁荞1号(中抗倒伏)均为松散型植株, 除乌克兰大粒荞外均为国内品种, 信农1号、宁荞1号和乌克兰大粒荞的生育期均为63 d, 西大花荞为70 d。
2012—2013连续2年在西南大学歇马科研基地(19°51′N, 106°37′E)进行田间试验。试验地土壤为沙壤土, 含有机质12.60 g kg-1、碱解氮73.20 mg kg-1、有效磷21.00 mg kg-1、速效钾106.00 mg kg-1、全氮0.84 g kg-1、全磷0.46 g kg-1、全钾17.50 g kg-1, pH 5.80。随机区组设计, 小区面积10 m2(2 m × 5 m), 3次重复, 分别于2012年8月26日和2013年8月28日播种, 人工条播, 基本苗90万株 hm-2, 行距33 cm, 种植6行, 小区之间留一空行, 区组间隔50 cm, 播种行与区组走向垂直, 试验地四周播种3行保护行, 播种前一次性施总养分≥45%的高浓度硫酸钾复合肥(含N 15%、P2O515%、K2O 15%) 300 kg hm-2作为种肥, 常规管理。
在荞麦收获前3 d调查各小区倒伏的株数和总株数, 倒伏率(%) = 倒伏株数/总株数×100%; 记录不同荞麦品种倒伏的时期。根据主茎与地面的夹角度数将倒伏程度分为0~5级[ 19], 0级为75°~90°、1级为60°~75°、2级为45°~60°、3级为30°~45°、4级为15°~30°、5级为0°~15°。待籽粒70%~80%成熟时单独收获和实测小区产量, 折合成公顷产量(kg hm-2)。
参照陈晓光等[ 20]和魏凤珍等[ 21]描述的方法。分别于开花期、灌浆结实期和成熟期取未倒伏的代表性植株3株, 取基部第2节间, 剥除叶鞘, 两端置高50 cm、间隔5 cm的支撑木架凹槽内, 在其中部悬挂一容器, 向容器内匀速倒入细沙, 至茎秆折断时停止, 用弹簧秤称取容器和细沙重, 即为茎秆抗折力参数(g); 量取茎秆基部至该茎(含穗、叶和鞘)平衡支点的距离, 即为茎秆重心高度(cm); 先剪去样品的根部, 将地上部洗净, 用滤纸吸干水分后称重, 即为茎秆鲜重(g)。倒伏指数(cm g g-1) = (茎秆重心高度×茎秆鲜重) /茎秆抗折力参数。
分别于第2节间形成后5、15、25、35和45 d, 在每个小区取未倒伏的典型植株10株, 将除去叶鞘的基部第2节间放入液氮中速冻, -20℃冰箱中保存, 用于木质素含量及其相关合成酶活性的测定。
1.5.1 木质素含量 参照陈晓光等[ 20]和林葵等[ 22]描述的方法, 准确称取0.5 g样品放于研钵中, 加入95%乙醇, 研磨至匀浆, 转移到离心管中, 2600× g离心5 min; 用95%乙醇洗涤沉淀物2次, 最后用乙醇︰正己烷=1︰2 (v/v)冲洗2次; 将沉淀物自然干燥并加入25%溴乙酰冰醋酸溶液, 使沉淀溶解, 加塞后于70℃水浴中保温30 min, 然后加入0.9 mL NaOH (2 mol L-1)、5 mL冰醋酸和0.1 mL羟胺盐酸(7.5 mol L-1), 最后用冰乙酸定容至15 mL。以蒸馏水代替底物进行同样反应为对照, 取上清液于280 nm处测定吸光值表示木质素含量(OD g-1 FW)。
1.5.2 苯丙氨酸解氨酶(PAL)活性 参照张志良和翟伟菁[ 23]的方法, 准确称取0.5 g样品放于研钵中, 加入6 mL硼酸钠-硼酸缓冲液(0.05 mol L-1, pH 8.8, 含5 mmol L-1巯基乙醇、1 mol L-1EDTA、少许PVP), 冰浴研磨至匀浆, 转入10 mL离心管中, 于4℃下以10 000× g离心15 min, 上清液即为酶提取液。取提取液0.2 mL, 加入1 mL由0.1 mol L-1硼酸钠缓冲液配制的L-苯丙氨酸(0.1 mol L-1)和2.8 mL蒸馏水, 摇匀, 37℃水浴反应30 min, 立即放入沸水中终止反应, 于290 nm处测定吸光值, 以1 mL蒸馏水代替底物进行同样反应为对照。以每小时 A290变化0.01为一个酶活力单位(U mg-1 FW)。
1.5.3 酪氨酸解氨酶(TAL)活性 酶液的提取同PAL。参照Kofalvi等[ 24]描述的方法, 取酶提取液0.2 mL, 加入2.8 mL硼酸缓冲液(0.1 mol L-1, pH 8.8), 及由此硼酸钠缓冲液配制的0.02 mol L-1的酪氨酸2 mL, 于37℃水浴反应30 min, 立即放入冰浴中终止反应, 以1 mL蒸馏水代替底物进行同样反应为对照, 于315 nm处测定吸光度值。以每小时 A315变化0.01为一个酶活力单位(U mg-1FW)。
1.5.4 4-香豆酸:CoA连接酶(4CL)活性 参照Knobloch等[ 25]描述的方法, 准确称取样品5 g, 放入冰浴研钵中并加入少许PVP和石英砂, 取10 mL 0.05 mol L-1 Tris-HCl缓冲液(pH 8.8, 含0.014 mol L-1的巯基乙醇和30%的甘油), 先加少量缓冲液和材料于冰浴中研磨成匀浆, 然后将剩余的缓冲液和已研磨成匀浆的材料转移到离心管中, 于4℃下以10 000× g离心15 min, 上清液即为酶提取液。取提取液0.4 mL, 加入反应液(香豆酸5 µmol mL-1, ATP 50 µmol mL-1, CoA-SH 1 µmol mL-1, MgSO4·7H2O15 µmol mL-1) 3 mL, 混匀后于40℃恒温水浴反应10 min, 于333 nm条件下测定吸光度, 以1 mL蒸馏水代替底物进行同样反应为对照, 以每小时 A333变化0.01为一个酶活力(U mg-1 FW)。
1.5.5 肉桂醇脱氢酶(CAD)活性 参照Morrison等[ 26]描述的方法, 准确称取0.4 g样品, 加入3 mL提取缓冲液(0.1 mmol L-1磷酸缓冲液, 内含15 mmol L-1巯基乙醇、1 mmol L-1 EDTA, pH 6.25)和少许PVP冰浴研磨至匀浆后于4℃下以10 000× g离心20 min, 上清液即为酶提取液。取1 mL 0.5 mol L-1的磷酸缓冲液, 1 mL NADP (2 mol L-1)、1 mL反式肉桂酸(1 mol L-1)和1 mL酶提取液, 在37℃反应30 min后, 于340 nm处测定吸光度值, 以煮沸1 min的酶液作为对照。以每小时 A340变化0.01为一个酶活力单位(U mg-1FW)。
分别于第2节间形成后5、15、25、35和45 d, 从每个小区取未倒伏的典型植株3株, 将除去叶鞘的基部第2节间中部切取成0.3~0.5 cm的横切环, 放入FAA固定液(70%乙醇90 mL、冰醋酸5 mL、福尔马林5 mL)中固定24 h以上, 按照脱水→透明→浸蜡-包埋→切片→粘片→脱蜡→染色→胶封等步骤制成厚度为15 µm的永久性石蜡切片。置Nikon Eclipse E200生物显微镜下拍照, 数出大维管束(内环维管束)数目、小维管束(外环维管束)数目和机械组织层数, 用Image-Pro Plus 6.0图片分析软件测量维管束面积、机械组织厚度和茎壁厚度。维管束截面积按椭圆面积公式 S = π ab/4计算, 式中 a、 b分别为纵、横2个方向的最大直径。
用Microsoft Excel 2003整理数据和作图, 用DPS v3.01进行统计分析, 采用Duncan’s新复极差(SSR)法检测显著性, 采用逐步回归法进行回归分析。
2.1.1 产量及倒伏情况 西大花荞未倒伏, 宁荞1号和信农1号在成熟期倒伏, 乌克兰大粒荞在开花期倒伏。与西大花荞、宁荞1号和信农1号相比, 乌克兰大粒荞倒伏程度严重, 产量较低(表1)。表明不同荞麦品种抗倒伏能力差异大。
2.1.2 茎秆抗折力参数和倒伏指数的比较 在荞麦开花至成熟阶段, 茎秆抗折力参数呈先增大后降低的趋势, 在灌浆结实期达最大值。在同一生育期内, 西大花荞的茎秆抗折力参数显著高于乌克兰大粒荞, 宁荞1号和信农1号介于西大花荞和乌克兰大粒荞之间, 且两者无显著差异。不同荞麦品种的倒伏指数存在一定差异, 乌克兰大粒荞的倒伏指数显著高于西大花荞、宁荞1号和信农1号(表2), 表明乌克兰大粒荞的茎秆抗倒伏能力较差, 易倒伏。同一荞麦品种在不同生育期的倒伏指数不同, 4个荞麦品种的倒伏指数均在灌浆结实期达最大值(表2)。
| 表1 不同荞麦品种产量及倒伏情况 Table 1 Yield and lodging situation of different buckwheat cultivars |
| 表2 不同荞麦品种茎秆抗折力参数和倒伏指数变化 Table 2 Changes of snapping resistance parameter of culm and lodging index in different buckwheat cultivars |
说明荞麦最容易在灌浆结实期倒伏, 这可能与灌浆结实期荞麦茎秆物质的再转运和生殖生长旺盛导致茎秆重心高度上移有关。
2.2.1 木质素含量 荞麦茎秆第2节间木质素含量随其生长发育的进程快速增加。在同一时期, 西大花荞的茎秆木质素含量显著高于乌克兰大粒荞, 宁荞1号与信农1号的茎秆木质素含量介于西大花荞和乌克兰大粒荞之间(图1)。这与各品种茎秆抗折力参数的变化规律一致, 表明荞麦茎秆第2节间的木质素含量高, 其抗倒伏能力强。
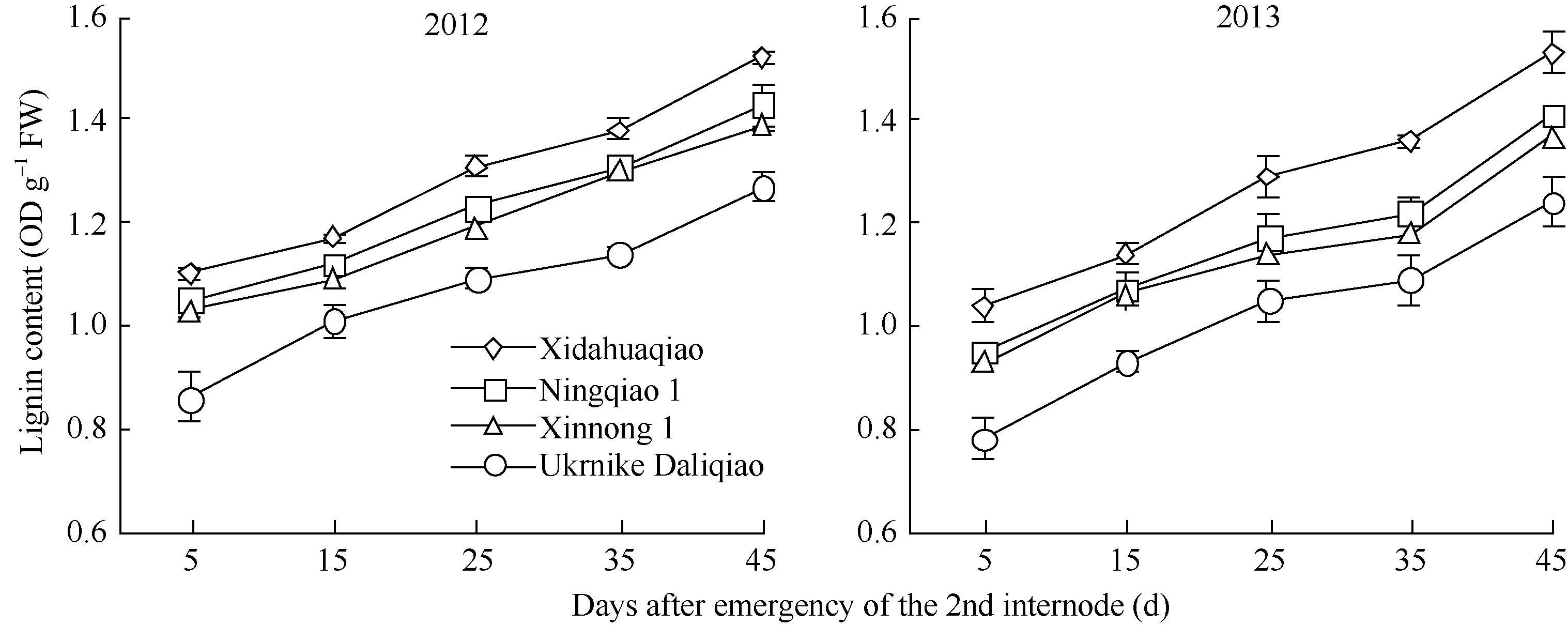 | 图1 不同荞麦品种木质素含量的变化Fig. 1 Changes of lignin content in different buckwheat cultivars |
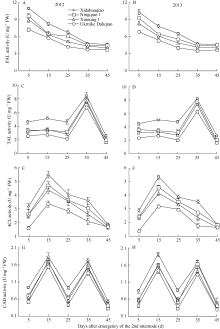
2.2.2 PAL活性 各荞麦品种茎秆第2节间PAL活性在灌浆结实期(第2节间形成后35 d)前随茎秆的生育进程呈下降趋势, 且西大花荞的PAL活性显著高于乌克兰大粒荞, 宁荞1号和信农1号的PAL活性介于西大花荞和乌克兰大粒荞之间; 在灌浆结实至成熟阶段, PAL活性变化不明显, 且品种间PAL活性差异不显著(图2-A, B)。说明抗倒荞麦品种茎秆第2节间PAL活性反应迅速, 并为木质素合成提供保障。
2.2.3 TAL活性 各荞麦品种茎秆第2节间TAL活性表现为在第2节间形成后25 d前变化不明显, 而后迅速升高后再迅速下降, 最大值出现在灌浆结实期(第2节间形成后35 d)。在荞麦茎秆第2节间生育过程中, TAL活性表现为西大花荞>宁荞1号>信农1号>乌克兰大粒荞(图2-C, D)。
2.2.4 4CL活性 随荞麦茎秆第2节间的生育进程, 各荞麦品种4CL活性表现为在第2节间形成初期
 | 图2 不同荞麦品种木质素合成相关酶活性的变化Fig. 2 Changes of metabolism-related enzyme activities in different buckwheat cultivars |
迅速升高, 而后迅速下降, 最大值出现在第2节间物质充实期(第2节间形成后15 d)。在荞麦籽粒成熟前, 荞麦茎秆第2节间4CL活性表现为西大花荞>宁荞1号>信农1号>乌克兰大粒荞, 而成熟后品种间4CL活性差异不显著(图2-E, F)。
2.2.5 CAD活性 在荞麦茎秆第2节间生育过程中, CAD活性呈双峰变化趋势, 第1个峰值出现在第2节间物质充实期(第2节间形成后15 d), 第2个峰值出现在灌浆结实期(第2节间形成后35 d)。茎秆第2节间CAD活性表现为西大花荞>宁荞1号、信农1号>乌克兰大粒荞(图2-G, H)。表明抗倒伏能力强的荞麦品种茎秆第2节间CAD活性强, 有利于木质素的合成。
2.3.1 机械组织层数 各荞麦品种茎秆第2节间机械组织层数在灌浆结实期(第2节间形成后35 d)前随茎秆生育进程逐渐增加, 而后变化不明显(图3-A, B), 表明荞麦茎秆基部节间的基本结构在灌浆结实期已构建完成, 机械组织层数已成定局。不同抗倒伏能力荞麦品种茎秆机械组织层数存在差异, 表现为西大花荞显著高于乌克兰大粒荞, 宁荞1号和信农1号介于西大花荞和乌克兰大粒荞之间(图3-A, B)。这是不同抗倒伏荞麦品种茎秆机械强度不同的明显特征。因此, 在荞麦茎秆发育过程中, 可根据其机械组织层数来预测其茎秆质量的的优劣。
2.3.2 机械组织厚度 随生育进程, 各荞麦品种茎秆第2节间机械组织厚度呈先增后减的趋势, 在灌浆结实期(第2节间形成后35 d), 荞麦茎秆机械组织厚度达最大值(图3-C, D)。在同一生育时期, 茎秆机械组织厚度表现为西大花荞>宁荞1号、信农1号>乌克兰大粒荞(图3-C, D), 这是不同抗倒伏荞麦品种茎秆机械强度不同的又一明显特征。
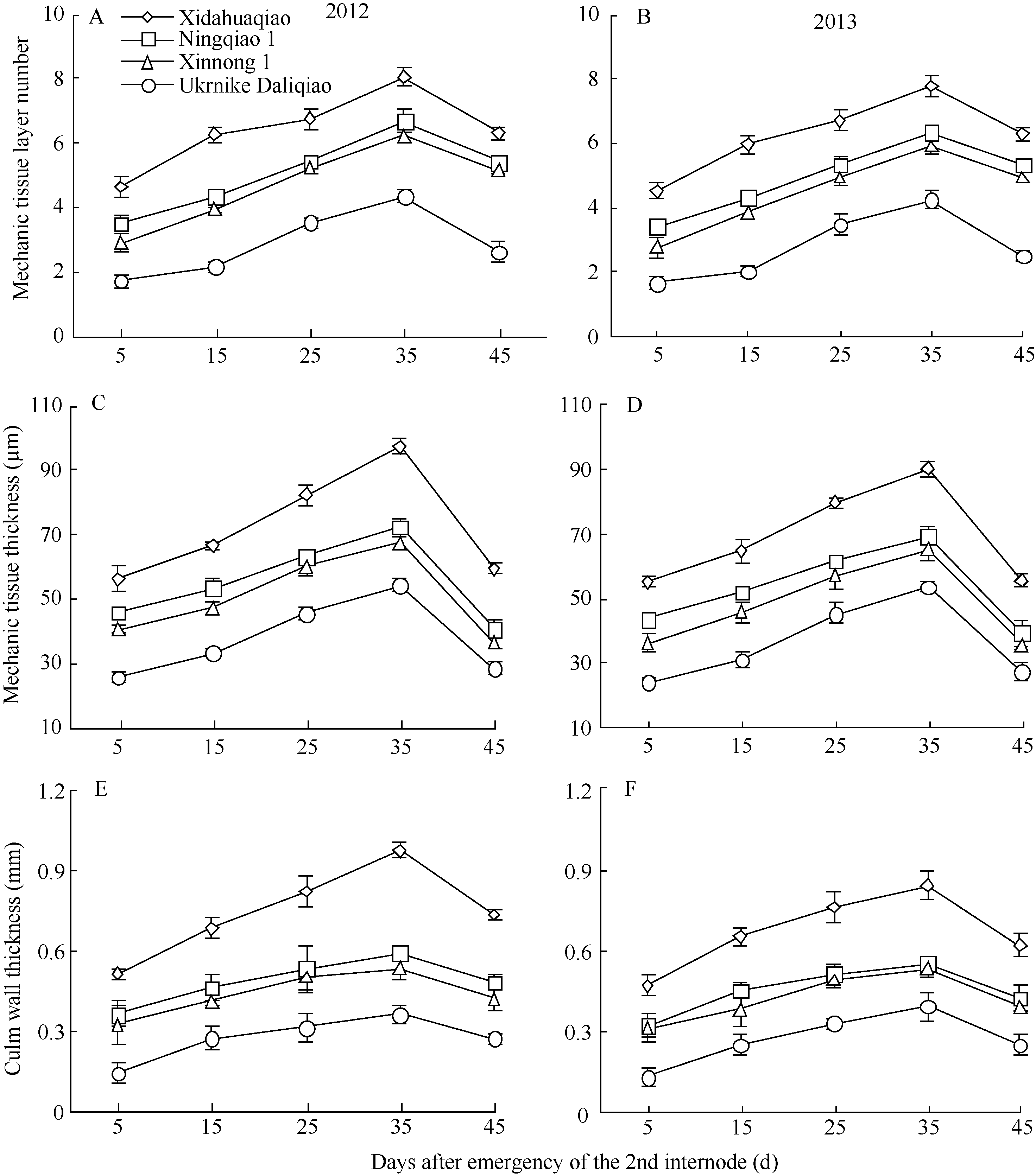 | 图3 不同荞麦品种机械组织层数、机械组织厚度和茎壁厚度的变化Fig. 3 Changes of mechanic tissue layer number, mechanic tissue thickness, and culm wall thickness in different buckwheat cultivars |
2.3.3 茎壁厚度 荞麦茎秆第2节间茎壁厚度随第2节间生育进程表现为先升高后下降的趋势(图3-E, F)。同一生育期内, 西大花荞茎秆茎壁厚度显著高于乌克兰大粒荞, 宁荞1号和信农1号介于西大花荞和乌克兰大粒荞之间(图3-E, F)。表明茎壁较厚的荞麦品种, 其抗倒伏能力强。
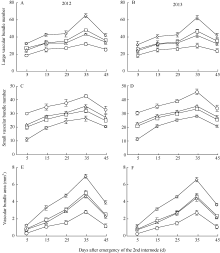
2.3.4 维管束数目 随生育进程, 各荞麦品种茎秆第2节间大维管束数目和小维管束数目均表现为先增加后降低的趋势, 且最大值均出现在灌浆结实期(第2节间形成后35 d), 在相同生育期, 西大花荞、宁荞1号和信农1号的大维管束数目多于其小维管束数目, 乌克兰大粒荞的大维管束数目少于其小维管束数目(图4-A~D)。同一生育期内, 茎秆第2节间大维管束数目表现为西大花荞>宁荞1号>信农1号>乌克兰大粒荞(图4-A, B), 小维管束数目表现为西大花荞<宁荞1号<信农1号<乌克兰大粒荞(图4-C, D)。表明抗倒伏能力强的荞麦品种, 其茎秆第2节间大维管束数目多, 而小维管束数目较少。
2.3.5 维管束面积 在荞麦茎秆第2节间生育过程中, 茎秆第2节间维管束面积表现为在灌浆结实期(第2节间形成后35 d)前缓慢增加, 而后迅速降低(图4-E, F)。茎秆第2节间维管束面积表现为西大花荞显著高于乌克兰大粒荞, 宁荞1号和信农1号介于西大花荞和乌克兰大粒荞之间(图4-E, F)。表明茎秆第2节间维管束面积大的荞麦品种, 其抗倒伏能力强。
| 表3 荞麦茎秆解剖结构和木质素代谢与抗倒伏能力相关分析 Table 3 Correlation coefficients among anatomical structure, lignin metabolism, and lodging resistance of culm in buckwheat |
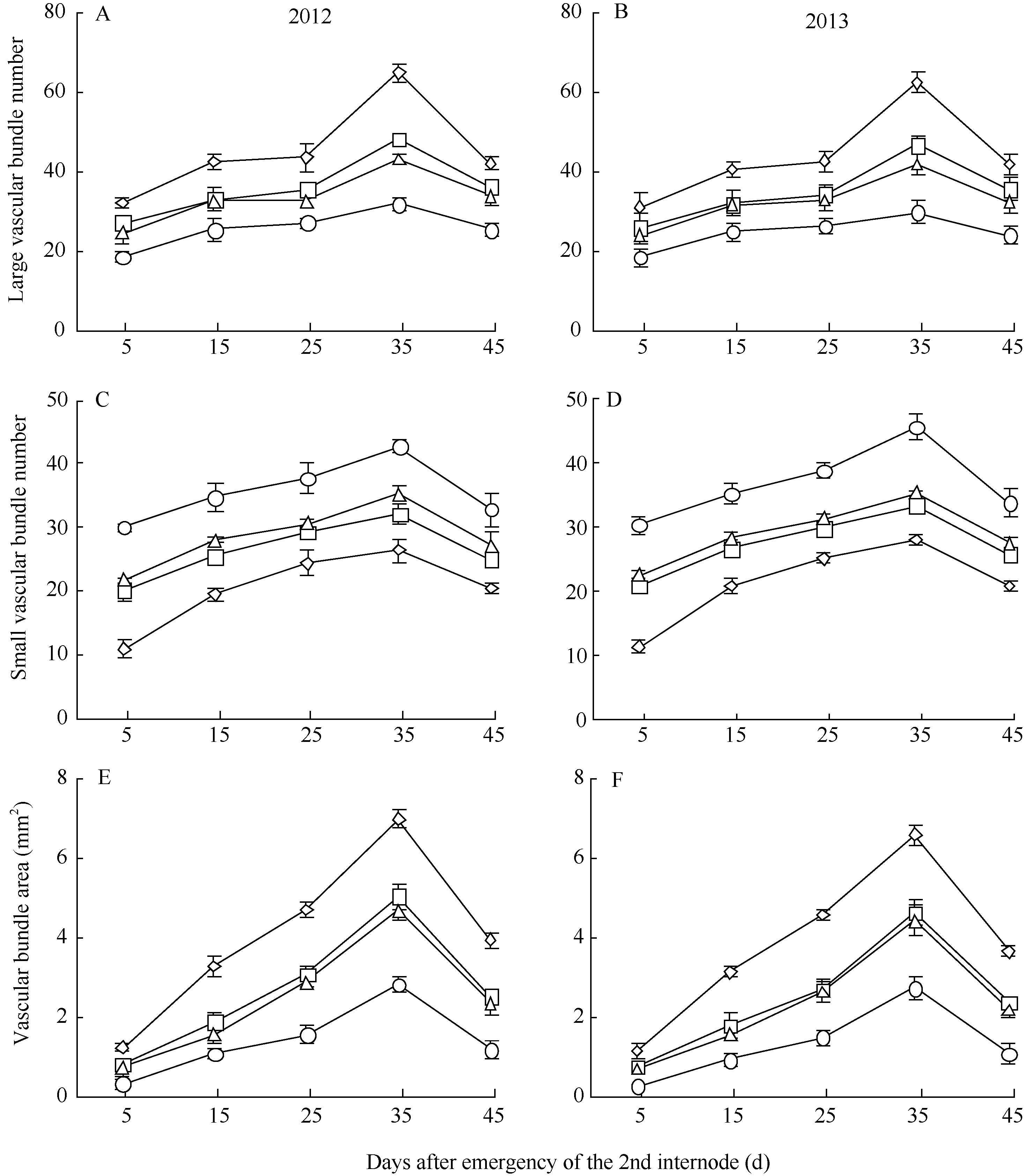 | 图4 不同荞麦品种维管束数目和维管束面积的变化Fig. 4 Changes of vascular bundle number and vascular bundle area in different buckwheat cultivars |
2.4.1 木质素代谢与抗倒伏能力 荞麦茎秆木质素含量与茎秆抗折力参数呈极显著正相关, 与倒伏率呈极显著负相关, 与倒伏指数呈显著负相关(表3)。表明荞麦茎秆木质素含量与茎秆抗倒伏能力密切相关, 木质素含量高, 荞麦茎秆抗倒伏能力强。木质素含量与PAL、4CL和CAD活性呈极显著正相关, 与TAL活性相关不显著(表3)。表明茎秆PAL、4CL和CAD活性是木质素含量增加的酶学基础。
2.4.2 解剖结构与抗倒伏能力 茎秆抗折力参数与机械组织层数、机械组织厚度、茎壁厚度和大维管束数目呈极显著正相关, 与维管束面积呈显著正相关; 倒伏指数与荞麦茎秆第2节间机械组织层数、茎壁厚度和大维管束数目呈极显著负相关, 与机械组织厚度和维管束面积呈显著负相关; 倒伏率与荞麦茎秆第2节间机械组织厚度、茎壁厚度和管束面积呈极显著负相关, 与机械组织层数和大维管束数目呈显著负相关; 小维管束数目与茎秆抗折力参数、倒伏指数和倒伏率的相关性不显著(表3)。表明荞麦茎秆抗倒伏能力与茎秆解剖结构密切相关, 茎秆抗倒伏能力大的荞麦品种, 其茎秆第2节间机械组织层数多, 机械组织和茎壁较厚, 大维管束数目多且维管束面积大。
为了明确影响荞麦茎秆木质素含量的关键酶和差别, 综合荞麦茎秆第2节间的PAL ( x1)、TAL ( x2)、4CL ( x3)和CAD ( x4)活性与木质素含量的多元回归分析, 剔除不显著项后, 建立的回归方程为 y = 0.017 + 0.352 x1 + 0.090 x3 ( R2=0.961, N=4, F=305.075, P< 0.0001)。PAL活性和4CL活性的偏相关系数分别为0.825和0.601, 表明荞麦茎秆的木质素合成受PAL和4CL的影响最大。
木质素具有提高细胞壁强度和增强茎秆机械强度的作用, 其含量与茎秆刚性密切相关[ 27]。研究表明, 小麦倒伏是茎秆木质素缺乏造成的, 木质素含量的增加可显著提高茎秆的机械强度, 增强茎秆抗压和倒伏能力[ 6, 7, 8], 且抗倒伏能力强的品种, 木质素含量高[ 28, 29]; 黄杰恒等[ 30]研究发现, 增加木质素含量, 可以增强油菜的抗倒伏能力。本研究表明, 与乌克兰大粒荞相比, 西大花荞的的茎秆抗折力参数大、倒伏指数和倒伏率小, 抗倒伏能力强, 其茎秆第2节间木质素含量在茎秆发育过程中始终较高; 相关分析表明, 木质素含量与茎秆抗折力参数呈极显著正相关, 与倒伏率呈极显著负相关, 与倒伏指数呈显著负相关。说明抗倒伏能力强的荞麦品种其茎秆木质素含量高。由此推断, 茎秆木质素含量可以作为荞麦茎秆抗倒伏能力的一个重要评价指标。
PAL位于苯丙氨酸代谢途径的入口, 是莽草酸途径中的一个限速酶[ 20], 它催化苯丙氨酸脱氨生成肉桂酸[ 29]。PAL活性下降时, 植物体内的木质素含量降低, 而PAL活性过量表达后木质素含量明显增加[ 31], 且抗倒伏能力强的小麦品种, 其茎秆PAL活性强[ 20]。本研究表明, 与易倒伏品种乌克兰大粒荞相比, 西大花荞茎秆第2节间PAL活性较高, 且PAL活性与木质素含量呈极显著正相关, PAL活性提高, 荞麦茎秆木质素含量增加。可见, PAL活性与荞麦抗倒伏能力密切相关。TAL催化酪氨酸脱氨直接生成香豆酸, 其仅存在于禾本科植物中[ 32]。本研究中, 荞麦茎秆第2节间存在一定量的TAL活性, 而TAL活性与木质素含量、茎秆抗折力参数、倒伏指数和倒伏率的相关性均不显著。因此, 荞麦茎秆抗倒伏能力与TAL活性不密切相关。4CL参与木质素合成的链接反应, 肉桂酸在4CL作用下生成相应的CoA酯[ 33]。研究表明, 抑制4CL的转基因植物的木质素含量均明显下降[ 34, 35]。本研究结果表明, 与乌克兰大粒荞相比, 西大花荞茎秆4CL活性提高, 其木质素含量增加, 抗倒伏能力强, 且4CL活性与木质素呈极显著正相关, 与倒伏率呈极显著负相关, 与倒伏指数呈显著负相关, 说明茎秆4CL活性在一定程度上能体现荞麦茎秆的抗倒伏能力。CAD参与木质素合成的最后一步还原反应, 是木质素合成中的重要酶类[ 36]。本研究中, 荞麦茎秆CAD活性与木质素含量呈极显著正相关, 与倒伏指数呈显著负相关, 与倒伏率呈极显著负相关, 与茎秆抗折力参数也呈正相关。说明CAD活性也能体现荞麦茎秆的抗倒伏能力。
茎秆抗倒伏能力与茎秆解剖结构密切相关。研究表明, 小麦基部茎秆的机械组织细胞层数越多、细胞壁越厚、木质化程度越高, 其抗倒性越强[ 37]。Pinthus[ 38]研究发现, 抗倒伏麦类作物品种维管束的直径和面积均大于不抗倒品种; 玉米的抗倒伏能力取决于机械组织和维管束的数量和质量[ 9]; 杨艳华等[ 12]认为抗倒伏水稻品种基部节间的维管束数目较多, 细胞层数较少, 细胞体积大, 维管束鞘较厚。本研究结果表明, 倒伏率与机械组织厚度、茎壁厚度和管束面积呈极显著负相关, 与机械组织层数和大维管束数目呈显著负相关, 而与小维管束数目的相关性不显著。说明荞麦茎秆机械组织层数和厚度、茎壁厚度、大维管束数目、维管束面积的大小也是造成荞麦植株抗倒伏能力高低的重要原因, 茎秆机械组织层数越多、机械组织和茎壁越厚、大维管束数目越多且维管束面积越大的荞麦品种, 其茎秆抗倒伏能力强。
荞麦茎秆抗倒伏能力与茎秆解剖结构和木质素代谢密切相关。茎秆木质素含量、机械组织层数、机械组织厚度、茎壁厚度、大维管束数目和维管束面积可以作为荞麦茎秆抗倒伏能力重要评价指标。茎秆木质素含量高、机械组织层数多、机械组织和茎壁厚、大维管束数目多且维管束面积大的荞麦品种, 其茎秆抗折力参数大、倒伏指数小、抗倒伏能力强。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|