

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻早衰叶突变体PLS2的遗传分析与基因定位
[张涛1, 2, * , 孙玉莹3, * , 郑建敏4, * , 程治军3 , 蒋开锋1, 2 , 杨莉1, 2 , 曹应江1, 2 , 游书梅1, 2 , 万建民3 , 郑家奎1, 2, 5, *  ]
]
]
|
|


叶片早衰引起叶绿素和其他大分子被降解, 叶片光合能力降低。这个过程常伴随着活性氧(ROS)的积累, 以及细胞中抗氧化酶(SOD、CAT和APX)活性的降低, 衰老相关基因(SAG)表达量上调, 最终导致整个植株过早成熟, 产量降低。因此, 研究水稻早衰遗传机制和基因功能对于水稻的遗传改良具有重要的作用和意义。PLS2是通过航天育种工程经空间辐射诱变得来的突变体, 在孕穗期表现早衰。与野生型相比, PLS2的光合能力降低, 株高变矮, 节间和穗长缩短, 分蘖数和有效分蘖数减少, 穗粒数和结实率明显下降, 千粒重降低, 穗发育不良, 灌浆不充分; 叶片的CAT活性显著降低、H2O2积累、死亡细胞增加, 叶绿体结构变差, 叶绿体中淀粉和嗜锇颗粒增多。黑暗处理加速突变体叶片衰老, 叶绿体超微结构球状化。利用PLS2/蜀恢527和PLS2/02428的隐性定位群体, 将
Leaf senescence induces degradation of chlorophyll and other macromolecules, reducing leaf photosynthetic capacity. This process is accompanied by the accumulation of reactive oxygen species (ROS), the decreasing of cell antioxidant enzyme (SOD, CAT, and APX) activity, and the increasing of aging related gene (SAG) expression, leading in early maturity and yield reduction. Therefore, studies on the genetic mechanism and gene function of premature senescence in rice, has the important effect and significance in genetic improvement of rice. PLS2 from space radiation mutation breeding project showed leaf senility, at booting stage. Compared with the wild type, in PLS2 the photosynthetic capacity decreased, the plant height, internode and panicle length shortened, tiller and effective tiller number reduced, number of grains per ear and seed setting rate were significantly lower, 1000-grain weight decreased, main panicle was stunted and grain-filling was not full. CAT activity decreased significantly in leaves, H2O2 accumulated, and the number of dead cell increased, chloroplast structures in leaves were worse, with more starches grains and osmiophilic granules. Dark treatment accelerated mutant leaf senescence, chloroplast ultrastructure was spheroidized. Using implicit localization population derived from PLS2/Shuhui 527 and PLS2/02428 the
水稻的生物产量和经济产量的形成主要依赖于光合作用, 叶片是光合作用的主要器官, 叶片的生长、发育与产量形成密切相关, 其衰老是植物叶片发育的最后阶段。叶片的早衰受遗传和环境因素(生物或非生物胁迫)影响, 可导致植物光合能力降低, 引起作物籽粒灌浆不足、结实率下降、千粒重变小等, 致使作物产量严重下降, 这是世界范围内粮食产量损失的重要原因之一[ 1]。叶片的衰老是植物在长期进化过程中形成的适应性, 是植物发育过程中的必然需要经历的生命现象[ 2], 是一个复杂的生物学过程, 涉及叶绿体发育、叶绿素合成与代谢、细胞程序性死亡、激素合成与代谢等多种途径的基因调控[ 3]。在拟南芥等双子叶模式植物上的研究表明, 在叶片衰老过程中, 叶片气孔缩小, 叶绿素和其他大分子(蛋白质、脂类和核酸等)被降解, 叶片光合活性降低、叶色逐渐变黄, 光合合成相关基因在衰老叶片中表达量下调, 大分子分解和运输的基因表达量上调[ 4, 5]。
目前, 在水稻中已经鉴定了一些衰老相关基因(senescence-associated genes, SAGs), 主要有 Sgr(t)[ 6]、 Osh69[ 7]、 Os12[ 8]、 OsDos[ 9]、 SGR[ 10]、 Sgr[ 11]、 NYC1[ 12]、 YGL1[ 13]和 SPL28[ 14]。与水稻衰老相关基因定位的研究报道有 Pse(t)[ 15]、 Pls1[ 16]、 Sms1[ 17]、 Pls3[ 18]、 Es-t[ 19]、 Lad[ 20]、 esl2[ 21]、 esl3[ 22]、 psf[ 23]和 lms[ 24]等。但克隆的衰老基因中直接调控基因较少, 中国科学院遗传与发育生物学研究所克隆了 NOE1[ 25]基因, 该水稻突变体 noe1的过氧化氢酶(CAT)活性降低, 叶片中过高的H2O2诱发了NO和S-亚硝基硫醇(S-nitro-sothiol)的产生, 导致叶片早衰。
四川省农业科学院水稻高粱研究所利用三系杂交水稻恢复系泸恢H103经空间辐射诱变获得了1个隐性早衰突变体PLS2 (premature leaf senescence 2)。该突变体在拔节、孕穗期叶片开始黄化衰老, 黑暗处理会加快其衰老。前期对PLS2进行了系统的表型鉴定、遗传分析和基因初步定位研究, 将该基因定位于水稻第3染色体上, 与目前报道的衰老基因不等位, 是一个新的早衰突变体[ 26]。
水稻早衰突变体PLS2材料是由四川省农业科学院水稻高粱研究所选育的三系杂交水稻恢复系亲本泸恢H103, 于2006年参加农业部实践8号卫星航天育种工程, 经空间辐射诱变而来。经连续在四川、海南和北京多代种植, 该突变体在孕穗期衰老表型表现明显[ 26], 突变性状稳定、受环境影响小, 遗传稳定。
将水稻突变体PLS2和野生型泸恢H103分别种植在四川德阳四川省农业科学院水稻高粱研究所试验基地、北京中国农业科学院作物科学研究所温室、昌平试验基地、光照培养箱以及海南试验基地, F2群体种植在四川德阳四川省农业科学院水稻高粱研究所试验基地。分蘖拔节期, 观察野生型与突变体倒三叶叶片表型, 考察分蘖数、有效分蘖数、结实率、株高、穗长、千粒重、穗粒数、单株产量等农艺性状。在孕穗期(抽穗前)观察F2单株倒三、倒四叶表型, 此时期, 倒三叶大部分变黄且倒四叶有变黄迹象则记为突变体表型单株, 表现绿色(正常色)为野生型表型单株( 图1)。在人工气候室种植用于暗培养的野生型与突变体幼苗, 30℃恒温, 90%恒湿, 14 h光照/ 10 h黑暗周期。
2008年在四川德阳, 以PLS2做母本分别与泸恢H103、蜀恢527、泸恢602、日本晴和02428杂交, 获得杂种F1。同年冬季在海南试验基地种植加代并收获F2种子, 2009—2010年夏在四川省农业科学院水稻高粱研究所德阳试验基地种植F2群体, 单苗栽插。孕穗期调查F2分离情况, 同时取各亲本、部分F2正常单株和全部F2衰老单株作为定位群体。
取人工气候室生长4周的野生型和突变体叶片, 在中国农业科学院食品加工研究所透射电镜室观察衰老及正常叶片样品情况。
采用改进的CTAB (cetyltrimethyl ammonium bromide)法提取亲本及定位群体的基因组DNA[ 27], 参照Rogers等的方法[ 28]利用SSR标记和InDel标记采用混合分离群体分析法(bulked segregant analysis, BSA)进行连锁分析。找到连锁标记后随机选取120株杂交F2群体中的衰老极端单株, 对初定位区间进行验证。最后用全部杂交F2代群体中的隐性极端单株, 进行基因精细定位和绘制包含目的基因所在区域的物理图谱。
参照林启冰等[ 29]的方法分析及开发设计InDel分子标记。
参考NCBI和Gramene网站上的注释和预测精细定位区间内的基因。用Primer Premier 3.0软件对区间内预测的基因设计测序引物, 用PCR方法分别扩增野生型泸恢H103和突变体PLS2的基因序列。琼脂糖凝胶电泳后, 用PCR产物纯化回收试剂盒(北京天根公司)回收目的片段, 并委托北京博迈德公司和中国农业科学院重大工程楼公共实验室测序。用DNAMAN软件对测序结果比对分析, 寻找区间内存在的核酸序列突变位点。
在温室条件下, PLS2在播种后6~8周, 第四叶片开始变黄衰老( 图1-A, B), 之后的叶片也随着生长的进程, 提前衰老。六叶期早衰表型相当明显, 叶绿素严重降解, 叶片黄化、干枯。随着植株的生长, 突变体叶片不断衰老并最终死亡。
在四川德阳(寡照、昼夜温差小)大田种植条件下, 插秧30 d左右后(秧龄35 d左右), 在抽穗期前, 倒三叶心叶抽出时, 先前抽出的倒四叶片就开始变黄衰老, 每个分蘖叶片的表现均类似; 始穗时先前抽出的倒三叶片就开始变黄衰老; 齐穗后倒二叶衰老; 灌浆完成时剑叶表现衰老, 完全成熟时剑叶完全衰老死亡。野生型品种泸恢H103在相同条件下可以保持25~30 d的绿色, 成熟时叶片颜色变化正常, 符合水稻自然衰老的规律。
 | 图1 野生型泸恢H103与突变体PLS2的表型A, B: 温室生长的六叶期野生型泸恢H103与突变体PLS2的表型; C: 野生型泸恢H103与突变体PLS2在抽穗期的表型; D: 野生型泸恢H103与突变体PLS2主穗的表型。Fig. 1 Phenotypes of wild type Luhui H103 and mutant PLS2 plantsA, B: Phenotypes of Luhui H103 and PLS2 plants at the 6th leaf stage; C: Phenotypes of WT and PLS2 plants at heading stage; D: Main panicle of wild type and PLS2 plants. |
在北京(强光、昼夜温差大)的大田种植条件下, 突变体在抽穗期衰老表型明显, 除少数叶片保持绿色外, 大部分叶片变黄、凋亡( 图1-C); 进入灌浆期, 突变体剑叶开始衰老、凋亡。而在相同环境下, 野生型泸恢H103的大部分叶片绿色维持到灌浆期结束。可以看出, 在各个生长阶段, 突变体PLS2叶片都比野生型的叶片表现出显著的早衰。最终导致突变体的穗发育不良, 籽粒灌浆不饱满、千粒重变小( 图1-D)。
两地大田种植结果比较: 在北京突变体抽穗期衰老表型明显, 在四川突变体孕穗期衰老表型明显, 在四川德阳(寡照、昼夜温差小)条件下比在北京(强光、昼夜温差大)条件下衰老时间提前10 d左右。这可能与光照强度及温度有关。
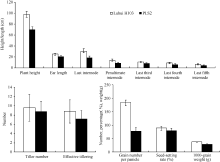
由于突变体叶片早衰发生的时期比较早, 其光合能力降低影响到整个植株的生长发育, 其株高变矮, 节间和穗长缩短, 分蘖数和有效分蘖数减少; 在穗发育时营养供应不足, 每穗粒数明显减少, 灌浆不充分; 后期叶片枯死, 导致千粒重明显变小。从 表1、 表2和 图2可以看出, 北京、四川德阳两点突变体株高均降低20%以上, 每穗粒数减少30%以上, 变异达显著水平; 千粒重下降20%以上, 变异达显著水平; 结实率在北京点变化不显著, 在德阳点下降48%, 变异达极显著水平; 植株产量也因为叶片的过早衰老而降低67%, 差异极显著。由此可见衰老对于作物产量的影响最为重要的性状是每穗粒数、结实率和千粒重。
| 表1 野生型与突变体株高、穗长和节间的长度比较 Table 1 Comparison of plant height, ear length, and internodes between wild type and mutant (cm) |
| 表2 野生型与突变体的单株产量、分蘖数、有效穗数、每穗粒数、结实率和千粒重比较 Table 2 Comparison of tiller number, effective tiller number, grain number per panicle, and seed-setting rate between wild type and mutant |
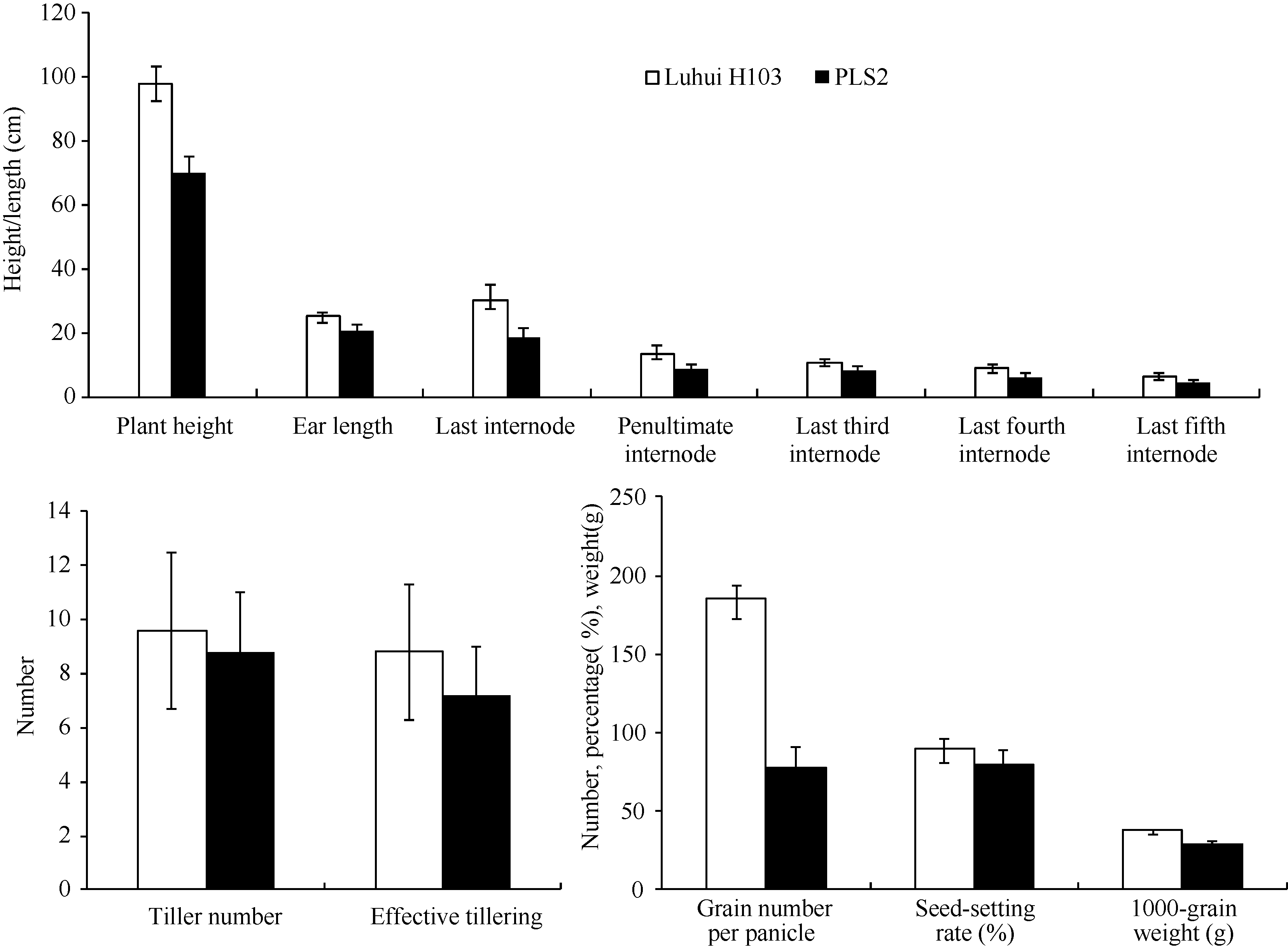 | 图2 野生型沪恢H103与突变体PLS2农艺性状比较Fig. 2 Agronomic trait comparisons between wild type Luhui H103 and mutant PLS2 plants |
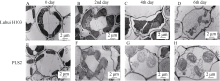
取人工气候室生长4周的野生型和突变体的幼苗, 持续黑暗处理, 每隔1 d取一次样品(第四片叶片)固定, 在透射电镜下观察叶绿体的降解情况。随着处理时间的延长, 野生型也表现出衰老的表型, 在黑暗处理6 d时, 野生型的叶绿体降解, 很少能观察到完整的叶绿体( 图3-D)。在相同生长条件下, 突变体各个时期的叶绿体结构都比较蓬松; 突变体在黑暗处理第2天, 其叶绿体色浅且形状不规则, 与野生型黑暗处理第4天的叶绿体形态相近( 图3-C, F), 之后叶绿体的体积反而变大, 充斥着淀粉粒, 表明叶绿体也出现降解问题( 图3-G, H)。持续黑暗加速叶片的衰老, 这与突变体在四川(寡照、低温差)比在北京(强光照、高温差)提前衰老的结果一致。
 | 图3 生长4周的野生型泸恢H103与突变体PLS2在持续黑暗下的叶绿体降解情况上排为野生型叶绿体在黑暗处理前(A)、第2天(B)、第4天(C)和第6天(D)的降解情况, 下排为突变体叶绿体在黑暗处理前(E)、第2天(F)、第4天(G)和第6天(H)的降解情况。Fig. 3 Chloroplast degradation of wide type Luhui H103 and mutant PLS2 leaves in continuous dark treatmentA: WT before the treatment; B: WT in the second day of the treatment; C: WT in the fourth day of the treatment; D: WT in the sixth day of the treatment; E: PLS2 before the treatment; F: PLS2 in the second day of the treatment; G: PLS2 in the fourth day of the treatment; H: PLS2 in the sixth day of the treatment. |
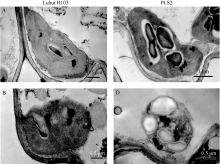
大田生长阶段, 在抽穗前取突变体中既有绿色部分也有黄色部分的叶片进行固定, 对应地, 选取野生型同生育期条件下的相同部位叶片。从 图4可以看出, 突变体绿色部位与黄色部位的叶绿体结构均比野生型的要差, 而且叶绿体中淀粉和嗜锇颗粒也较多。
 | 图4 野生型泸恢H103与突变体PLS2在抽穗之前的叶绿体结构左侧为野生型叶片的绿色部分(A)和黄色部分(B)叶绿体结构, 右侧为突变体叶片的绿色部分(C)和黄色部分(D)叶绿体结构。Fig. 4 Chloroplast structures of wide type Luhui H103 and mutant PLS2 leaves before heading stageA, B: Chloroplast structures of WT blade (corresponding leaf with C and D); C, D: Chloroplast structures of green and yellow parts in senescencing leaf of PLS2. |
早衰突变体PLS2分别与泸恢H103、蜀恢527、日本晴(粳稻)和02428 (粳稻)杂交, 无论在海南或四川种植, F1叶色与野生型相同, 说明该早衰性状为隐性遗传。F2在四川德阳试验田种植, 抽穗期前调查叶片表型及分离情况, 突变体叶与正常叶分离非常明显, 早衰叶和正常叶的分离都符合1∶3的分离比( 表3), 说明叶片早衰性状受一对隐性基因控制。
| 表3 早衰叶突变体和不同亲本杂交F1、F2的表现 Table 3 Performance of presenile leaf mutants and F1, F2 of hybrid with different parents |
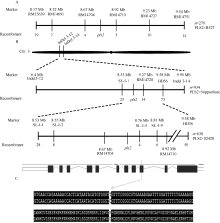
筛选覆盖水稻全基因组的537对SSR分子标记, 得到55对在早衰突变体PLS2与蜀恢527间有多态性的引物, 利用270个F2突变个体单株将目的基因 pls2定位在标记RM14704 (8.67 Mb)与RM14710 (8.92 Mb)之间, 物理距离约370 kb。为进一步精细定位, 以PLS2/日本晴和PLS2/02428的籼粳交F2群体作为定位群体, 在标记RM14704与RM14710之间设计InDel标记( 表4), 对PLS2×日本晴F2群体中934个隐性极端单株进行连锁分析, 将目的基因 pls2定位在第3染色体短臂上SL-I-1 (8.53 Mb)与RM14728 (9.27 Mb)之间( 图5)。随后利用638个“PLS2×02428” F2群体中的隐性极端单株进行连锁分析, 将目的基因 pls2定位在标记SL-I-2 (8.55 Mb)与SL-I-5 (8.76 Mb)之间( 图5和 表4), 整合2次定位的连锁图谱, 目的基因 pls2最终被定位在标记RM14704 (8 674 283 bp)与SL-I-5 (8 758 394 bp)之间, 物理距离84.11 kb。
| 表4 基因定位所用的引物 Table 4 Primers used in fine mapping |
 | 图5 pls2基因的图位克隆A, B: pls2基因的精细定位; C: 定位区间基因 LOC_Os03g15840的编码区存在碱基改变。Fig. 5 Map-based cloning of pls2 geneA, B: fine mapping of pls2 gene; C: sequence variations of genes in the mapping interval between WT and pls2. |
参考网站(http://rice.plantbiology.msu.edu/analyses_search_locus.shtml)对定位区间内的基因功能进行预测( 表5), 该区间包含14个基因( 表5)。
| 表5 定位区间内基因的功能预测 Table 5 Predicted functions of genes in the mapping interval |
对区间内的14个基因测序, 包括基因上游3 kb启动子区域和下游1 kb的终止子区域, 发现基因 LOC_Os03g15840的编码区存在碱基改变。基因 LOC_Os03g15840全长3268 bp, 包含11个外显子和10个内含子, CDS为1248 bp, 编码416个氨基酸, pls2基因在 LOC_Os03g15840的第9个外显子中的第41位的碱基C被替换为T, 导致精氨酸被半胱氨酸所替换( 图5-C)。通过在NCBI上的比对分析发现, 它是一个进化上非常保守的位点。对对照和5个水稻品种IRAT129、日本晴、南京11、泸恢H103 (野生型)、宜香1B和一个矮秆突变体的测序结果表明,该位点不存在序列上的差异( 图6-A)。进一步比较3个水稻的同源基因以及高粱、玉米、拟南芥、蓖麻、短柄草、黄瓜、绿藻等21条氨基酸序列发现, 该位点非常保守( 图6-B), 该基因进化上与大麦、二穗短柄草亲缘关系最近, 与高粱、玉米和水稻中另一个同源基因亲缘关系较近( 图6-C)。推测这个编码含有糖基转移酶结构域蛋白的基因可能是候选基因。
 | 图6 突变体基因 pls2在 LOC_Os03g15840的突变位点以及该基因的进化分析A: 基因 LOC_Os03g15840在不同水稻品种间的DNA序列比对分析; B: LOC_Os03g15840在不同物种间的氨基酸序列比对分析; C: pls2基因的进化分析。Fig. 6 Mutation position in pls2 of LOC_Os03g15840 compared with the homologous genes, and the phylogenetic tree of pls2A: Sequence BLAST of the LOC_Os03g15840 of pls2 between different rice varieties; B: Sequence BLAST of the LOC_Os03g15840 of pls2 between different plant species; C: Phylogenetic tree of pls2. |
衰老是植物体生长发育的最后阶段。在衰老过程中, 植株体内的营养物质再分配, 由衰老器官转移到生殖器官等代谢旺盛的部位。植株的衰老对其营养生长和生殖生长都具有重要的影响[ 30]。过早衰老会导致产量大幅度降低, 延迟衰老会增加产量, 研究表明, 水稻叶片每推迟1 d衰老, 产量可增加2%[ 31]。可见研究衰老并寻找切实可行的延缓衰老的措施对于提高水稻产量具有重要的意义。
突变体PLS2在播种后6~8周出现衰老表型(温室), 第4片叶片开始抽出, 老的叶片出现早衰, 叶绿素严重降解, 叶片黄化、干枯, 六叶期就能被清楚地区别开突变体和野生型。随着植株的生长, 植株下部叶片也逐渐提前衰老, 突变体在抽穗期衰老表型最为明显, 除少数叶片保持绿色外, 大部分叶片变黄、死亡; 进入灌浆期, 突变体剑叶开始衰老、死亡。而野生型在相同环境下, 大部分叶片绿色维持到灌浆期结束。由于突变体叶片从苗期开始早衰, 光合能力降低, 从而影响到整个植株的生长发育, 其株高变矮、节间和穗长缩短, 分蘖数和有效分蘖数减少, 每穗粒数明显减少, 千粒重明显降低。持续黑暗能够诱导衰老的发生, 对生长4周的野生型和突变体的幼苗进行遮光黑暗处理, 突变体叶绿体降解速率与野生型相比更为快速。突变体各个时期的叶绿体结构都比较蓬松, 黑暗处理第2天的突变体其叶绿体形状不规则, 之后充斥着淀粉粒。
随后将突变体基因 pls2定位在水稻第3染色体标记RM14704 (8 674 283 bp)与SL-I-5 (8 758 394 bp)间, 物理距离84.11 kb, 包含14个基因。目前该区域未见存在衰老基因报道, pls2是一个新的水稻叶片早衰基因。测序分析仅发现 pls2在 LOC_Os03g15840第9个外显子第41位的C被替换为T, 导致精氨酸被半胱氨酸所替换。通过在NCBI上的比对分析发现, 该位点是一个进化上非常保守的位点, 在高等植物和藻类中没有改变。 pls2的候选基因 LOC_Os3g15840编码水稻中的一个糖基转移酶(glycosyltransferases, GTs)。它是植物次生代谢产物, 参与生物合成、保持内稳态, 并调控信号分子和防御化合物的活性, 依赖UDP的糖基转移酶(UDP-dependent glycosyl-transferases, UGTs)催化糖从活性糖供体转移到小分子受体上, 形成糖苷键。已有的研究表明, 糖基转移酶与植株抗性以及植株衰老等过程有一定的关系。在拟南芥中, 过表达PsUGT1的拟南芥叶片衰老加速[ 32], 干扰AtUGT85A7的拟南芥叶片衰老延迟、植株生长速率降低、生命周期延长。因此, 本研究中突变基因 LOC_Os3g15840可能是造成叶片早衰的原因。
水稻衰老突变体PLS2是一个新的衰老突变体, 由于突变体叶片早衰, 光合能力较野生型降低, 导致株高变矮、节间和穗长缩短, 分蘖数和有效分蘖数减少, 每穗粒数和结实率下降明显, 千粒重降低, 主穗发育不良, 籽粒灌浆不饱满。黑暗处理能加速衰老, 在抽穗期前, 突变体叶尖或叶片基部的叶绿体结构均比野生型的要差, 而且叶绿体中淀粉和嗜锇颗粒增多。 pls2基因被定位在水稻第3染色体上标记RM14704 (8 674 283 bp)与SL-I-5 (8 758 394 bp)之间84.11 kb范围内, 预测编码含有糖基转移酶(glycosyltransferases, GTs)结构域蛋白的 LOC_Os3g 15840是候选基因。这为进一步基因克隆和水稻早衰机制的研究奠定了基础。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|