{kind=link}
红麻光周期钝感材料的鉴定与遗传分析
[张立武, 黄枝秒, 万雪贝, 林荔辉, 徐建堂, 陶爱芬, 方平平, 祁建民*  ]
]
]
|
|


开花期是影响红麻纤维产量和品质的关键因素之一。本文通过分期播种调查6份新引育的红麻品系的光周期反应, 结果表明其光周期反应敏感度变化在36.0%~56.2%之间, 其中赞引1号最低(36.0%), 福红952B最高(56.2%)。将赞引1号与福红952B杂交, 在自然短日照条件下对其正反交F1分析表明, 开花期性状受核基因控制, 不存在细胞质效应, 光周期敏感对光周期钝感为显性。在自然短日照条件下对该组合的4个群体(P1、P2、F1和F2)联合分析发现, 赞引1号的光周期钝感特性受1对加性-显性主基因和加性-显性-上位性多基因模型(D-1)控制, 主效基因的加性效应值为8.2 d, 遗传率为80.2%。该研究有助于红麻光周期钝感种质改良及主效基因定位。
Flowering stage is one of the key factors affecting fiber yield and quality in kenaf. In this study, photoperiod response sensitivity of six newly bred kenaf lines was identified via different sowing time. The results showed that photoperiod response sensitivity of various lines ranged from 36.0% to 56.2%. Among them, Zanyin 1 was insensitive one to photoperiod response (36.0%) while Fuhong 952B was sensitive (56.2%) one. To uncover the genetic basis of flowering stage under the condition of natural short day, we further used the joint segregation analysis of four populations (P1, P2, F1, and F2) in a cross of Zanyin 1 × Fuhong 952B. The analysis of F1 and reciprocal F1 revealed that flowering stage was controlled by nuclear genes instead of cytoplasmic genes, and the photoperiod response sensitivity genotype was almost completely dominant. Flowering stage is best described by the D-1 genetic model, a case of one additive-dominance major gene as well as additive-dominance-epistasis polygenes. The additive effect of the major gene was 8.2 days. And heritability of the major gene was 80.2%. These findings will facilitate breeding strategies for the improvement of photoperiodic insensitive germplasm as well as the major gene mapping in kenaf.
红麻( Hibiscus cannabinus L.)是锦葵科木槿属一年生韧皮纤维作物, 具有适应性广、耐盐碱、耐涝、耐贫瘠、抗干旱能力较强、纤维产量高、品质优良及生长快速等特性。其纤维可作为纺织原料、工业原料和生物质能源等, 倍受业界对新产品新材料开发的重视[ 1, 2]。红麻是短日照植物, 短日照促进红麻的生殖发育, 而长日照的作用则相反[ 3, 4]。由于光周期作用, 持续下雨或在低纬度地区种植等因素, 会导致红麻生育期缩短, 提早开花, 降低纤维的产量和品质[ 3]。对红麻材料进行光周期反应的遗传研究, 有助于了解不同材料光周期反应的遗传差异, 对红麻光周期钝感新品种选育与推广具有指导意义。
在红麻光周期反应的研究中, 研究人员主要从红麻的光周期钝感材料鉴定、遗传规律及生理生化指标测定等方面进行了分析[ 3, 4, 5, 6, 7]。李德芳等[ 3]和徐建堂等[ 6]鉴定不同红麻品种光周期钝感材料并进行遗传分析, 认为光周期钝感反应中存在主基因控制。彭定祥等[ 5]分析了红麻品种光周期反应特性, 认为裂叶型叶片按生长发育不同时期发生叶周期变化。徐建堂等[ 7]研究不同光周期反应的叶片生理指标, 认为叶片是感受光周期诱导的主要部位, 叶片内的生理指标变化与光周期反应敏感性类型有重要的关系。然而, 不同来源的品种光周期反应有一定差别[ 4]。在育种实践中, 福建农林大学麻类遗传育种研究室从红麻可能的起源地赞比亚[ 8]引进并选育了一份优良品系赞引1号, 该品系具有无刺、短柱头等特点, 其光钝感特性表现突出, 但其光周期反应的遗传特点不明。
本研究通过对引育的红麻品系赞引1号进行光周期反应敏感度分析, 并探讨其遗传特点, 可以为红麻的光周期钝感种质改良提供理论依据。
来源不同的6份红麻品系均来自福建农林大学麻类遗传育种研究室, 分别为赞引1号、福红R-1、KG121、福红航1B 、福红B3和福红952B。其中, 赞引1号是从赞比亚引进并选育的一份优良品系, 茎红色, 圆叶型, 植株高大; 而其他材料是课题组选育的高世代品系, 茎绿色, 裂叶型, 植株较高。
2012年于福建农林大学试验田分期播种6份红麻品系。采用4个播期, 分别为5月1日、6月1日、7月1日与8月1日。采用人工去雄, 按极端光周期敏感与光周期钝感交配设计方式配制正反交F1, 亲本套袋自交, 2012年12月将亲本和正反交F1同时播种于福建农林大学海南南繁试验田, 2行区, 行长2 m, 株行距1.20 m×0.20 m, 采用单因素随机区组设计, 田间管理同大田。由于亲本和正反交F1是不分离世代, 以小区为单位记载开花期, 即以试验小区全部麻株为观测对象, 50%植株开花的日期为开花期。
2013年8月, 于福建农林大学试验田播种以赞引1号和福红952B为亲本杂交获得的4个群体(P1, F1, P2和F2), 株行距和田间管理同2012年。不分离世代P1、F1和P2开花期按试验小区记录, 而分离世代F2群体按单株记录, 开放第一朵花的日期为开花期。2013年12月于福建农林大学海南南繁试验田播种该组合30份F2:3株系(由于其他株系开花偏迟, 12月还未收获种子), 株行距和田间管理同于2012年, 每株系约20个单株。各F2:3株系按单株记录, 开放第一朵花的日期为开花期。
采用韩天富等[ 9]介绍的方法计算各品系的光周期反应敏感度PRS (photoperiod response sensitivity) (相当于短日开花促进率), PRS(%)=(首期播期的开花持续时间-末期播期的开花持续时间)/首期播期的开花持续时间×100。
参照Gai等[ 10]和章元明等[ 11]介绍的方法对杂交组合的4个群体进行开花期的遗传分析。共使用了24个遗传模型, 其中A模型是1对主基因遗传模型, B模型为2对主基因遗传模型, C模型是无主基因模型, D模型为1对主基因+多基因遗传模型, E模型为2对主基因+多基因遗传模型, 各类模型再根据基因的加性、显性和上位性效应分成若干个模型进行分析。分析主要包括4个步骤: 先通过极大似然法和IECM (iterated expectation and condition maximization, 迭代期望及条件极大值)算法对混合分布中的有关成分分布参数作出估计; 然后根据AIC值选择候选模型; 再对候选模型进行适合性测验; 在最适遗传模型下, 计算主基因和多基因效应值、方差等各种遗传参数。分析软件由南京农业大学章元明教授提供(http://jpkc.njau.edu.cn/swtj/)。
由 表1可以看出, 不同类型品系的光周期反应有明显差别, 表现为自然长日照条件下(如5月1日播期), 不同品系开花期存在一定差异, 而在短日照条件下(如8月1日播期), 不同品系开花期差异非常明显。在5月1日播期中, 各品系的开花持续时间变化在147~175 d之间, 最大相差28 d; 而在8月1日播期中, 中红麻16和福红952B的开花持续时间分别为62 d和66 d, 赞引1号的开花持续时间为112 d, 后者是前者的近2倍。
利用分期播种试验计算不同的红麻品系光周期反应敏感度(PRS)( 表1)。福红952B光周期反应敏感度高达56.2%, 而赞引1号光周期反应敏感度最低(36.0%), 表现为光周期反应钝感。这应与赞引1号是从低纬度国家赞比亚引进选育而成的有关, 而福红952B是在亚热带春夏播条件下育成。
| 表1 不同播期红麻品系开花期及光周期反应敏感度 Table 1 Flowering stage and photoperiod response sensitivity of lines under different sowing time in kenaf |
为了剖析红麻光周期钝感品系赞引1号的遗传特点, 利用来源不同的光周期敏感品系福红952B与其配制正反交F1。在自然短日照条件下(2012年12月10日在海南三亚播种), 比较各亲本及正反交F1的开花期( 表2), 正反交之间在该性状上差异不显著, 说明开花期性状受核基因控制, 不存在细胞质效应。正反交F1开花期表现光周期敏感, 表明在该组合中光周期敏感对光周期钝感在杂交后代中以显性的方式表现出来。
| 表2 自然短日照条件下红麻光周期钝感基因等位性分析 Table 2 Photoperiod insensitive allelic analysis in kenaf under the condition of natural short day |
从 图1可以看出, 杂交组合福红952B×赞引1号中F2群体开花期在自然短日照条件下(2013年8月1日在福州播种)的频率分布情况。亲本福红952B的开花持续时间为78 d, 赞引1号的开花持续时间为111 d, 属2个极端值的亲本类型; F1的开花持续时间为80 d, 接近于亲本福红952B; F2群体包含117个单株, 各单株的开花持续时间变异幅度大, 变化在71~115 d之间, 存在一定的超亲遗传现象。而且开花期呈明显的双峰分布, 偏离正态分布, 表明有主效基因控制该组合开花期的表现。
为了确认该结果, 有必要以其峰低作为界限来划分早花组和迟花组。结果表明, F2群体早花组和迟花组分别为85株和32株, 经卡方测验, 分离比例符合3∶1 (χ2 = 0.413, P>0.05)。表明该组合开花期主要受一对主效基因的控制, 光周期敏感对光周期钝感为显性。
为了确定F2群体各单株主效基因的基因型, 以福红952B为分界点, 即开花持续时间小于或等于75 d, 将该组合部分F2:3株系(30份)在自然短日照条件下(2013年12月10日在海南播种)进行后代验证。除了2份F2:3株系出现开花期性状分离现象, 28份F2:3株系未出现分离, 表现为早花。说明这28份F2:3株系应该属于纯合光周期敏感基因型, 而出现开花期性状分离的2份F2:3株系属于杂合光周期敏感基因型。
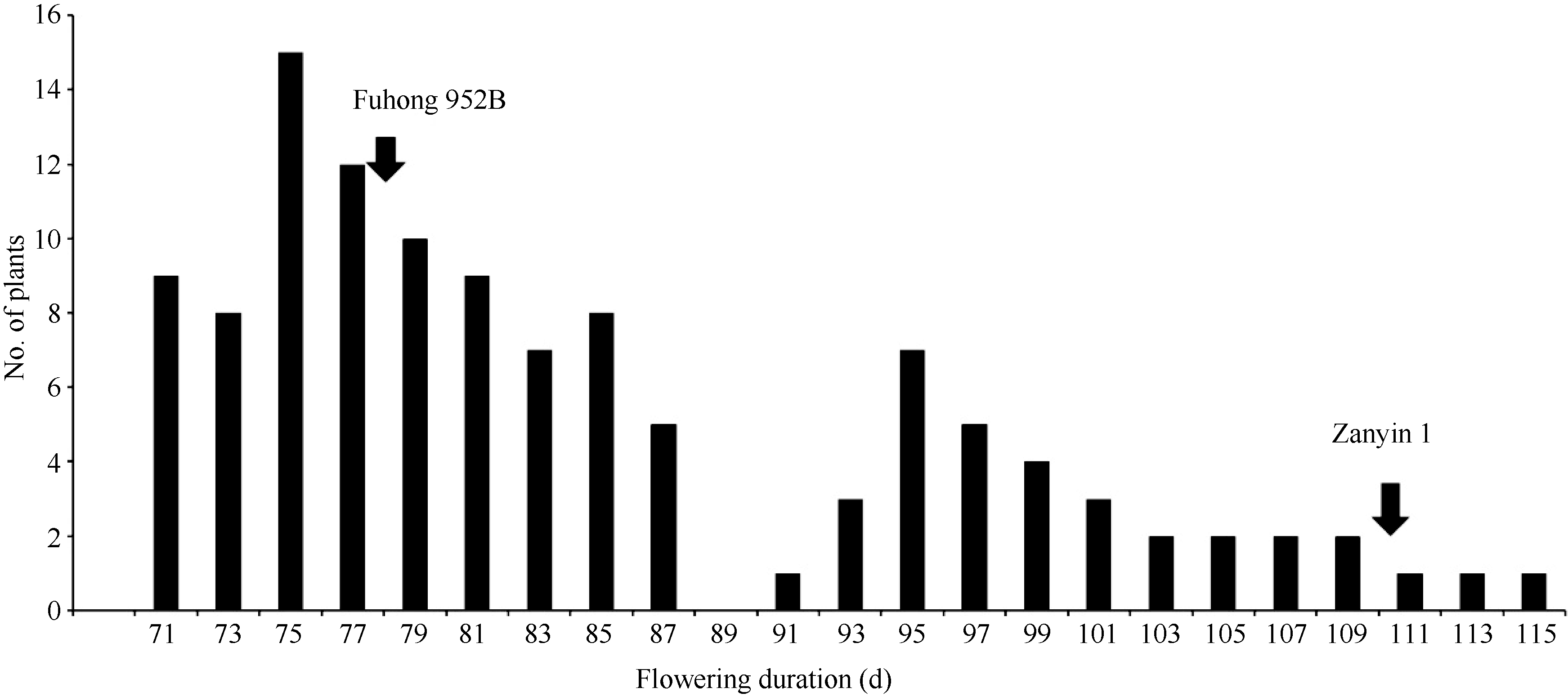 | 图1 在自然短日照条件下杂交组合福红952B×赞引1号F2群体开花持续时间的频率分布Fig. 1 Frequency distribution of flowering duration in the F2 population of Fuhong 952B × Zanyin 1 under the natural short day |
为了进一步分析主效基因的遗传信息, 用主基因+多基因遗传模型的多世代联合分析法对该组合不同群体的开花期进行分析, 得到 表3中24种遗传模型的AIC值。
| 表3 自然短日照条件下红麻杂交组合福红952B×赞引1号开花持续时间不同遗传模型的AIC值 Table 3 AIC values under various genetic models for flowering duration in the cross of Fuhong 952B × Zanyin 1 under natural short day |
比较AIC值大小, 发现D-0和D-1的AIC值最小, 确定为候选的遗传模型。为确定最优的遗传模型, 进行了适合性检验( 表4)。达到显著的统计量在D-0和D-1模型下均为0个。由于D-1模型AIC值最小, 确定D-1模型为最优模型。即杂交组合福红952B×赞引1号开花期受1对加性-显性主基因和加性-显性-上位性多基因模型控制。
| 表4 自然短日照条件下红麻杂交组合福红952B ×赞引1号4个群体在候选模型下的适合性检验 Table 4 Test of goodness-of-fit in candidate models in four populations of the cross of Fuhong 952B × Zanyin 1 under natural short day |
根据成分分布计算杂交组合福红952B×赞引1号4个基本世代在D-1模型下的一阶遗传参数和二阶遗传参数( 表5)。估计的主基因加性效应值(8.2)为正值, 说明赞引1号对开花期发挥着增效作用。主基因的显性效应值为-8.1, 其显性度为-0.99, 说明光周期敏感对光周期钝感为完全显性。主基因的遗传率为80.2%, 多基因遗传率为2.2%, 主基因的遗传效应占总效应的百分比为97.3%, 说明开花期主基因遗传率较大。
| 表5 自然短日照条件下红麻杂交组合福红952B×赞引1号4个群体在D-1模型下开花持续时间各遗传参数估计值 Table 5 Estimates of genetic parameters of flowering duration in four populations of the cross of Fuhong 952B × Zanyin 1 under natural short day |
开花期仍是植物遗传育种研究的热点。红麻品种(系)光周期反应程度的遗传差异不仅影响红麻的种子产量, 还与纤维产量和品质密切相关。掌握其遗传规律是红麻光周期钝感品种(系)改良的前提。本研究分期播种试验表明, 不同来源的品系在自然短日照条件下开花期差异明显。这与李德芳等[ 3]报道的较一致。但本研究与之不同的是, 采用分期播种方法定量检测各品系的光周期反应。本研究采用4个播期(5月1日、6月1日、7月1日与8月1日)来检测不同品系的光周期反应敏感度, 变化在36.0%~56.2%之间。末一个播期(8月1日)可以划分为短日照条件, 采用5月1日与8月1日2个播期,可以计算出各品系的光周期反应敏感度。按照邓丽卿[ 4]介绍的花蕾促进率来划分, 红麻光周期反应敏感度小于50%为光周期钝感。赞引1号的光周期反应敏感度为36.0%, 可划为光周期钝感材料。
用合适的遗传交配设计所获得的遗传特点能为育种方案的制定提供较多的信息, 开花期性状可通过遗传育种手段加以改良。就本试验而言, 开花期主要受加性基因作用方式控制, 并且具有较高的遗传率。这表明在早代选择开花期性状可收到较好成效, 类似结果在其他作物也有所体现[ 12, 13, 14, 15]。主基因+多基因模型分析表明, 在自然短日照条件下赞引1号与福红952B组合中开花期表现主要受一对主效基因控制, 并且光周期敏感对光周期钝感为显性。虽然使用的红麻材料不一样, 但结果与李德芳等[ 3]和徐建堂等[ 6]报道类似。这说明, 对于特定的红麻杂交组合, 存在主效基因控制光周期反应。当然, 本研究虽然对该组合开花持续时间小于或等于75 d的30份F2:3株系进行了后代验证, 但未能进行其他F2:3株系主效基因的基因型鉴定。为了确定纯合光周期钝感基因型的F2:3株系, 尚需进一步在自然短日照条件下进行后代验证。由于光周期钝感表现为隐性遗传, 为了利用红麻杂种优势[ 16], 应选择双亲都是光周期钝感材料, 这对选配优良的杂交组合有着特殊的意义。因此, 在育种中必须重视控制开花期性状基因的加性、显性效应, 采用适当的育种手段聚合更多的加性基因和有利基因。当然, 选育优良红麻品种还需兼顾丰产性、纤维品质、抗逆性与适应性等性状。
随着研究的不断深入, 应用遗传连锁图谱可以将控制开花期性状的QTL分解开来, 逐个研究其效应和作用方式[ 17]。而亲本的选择直接影响到构建连锁图谱的难易和所建图谱的适用范围。一般来说, 应该选择亲缘关系较远而差异大的品种(系)作为构建作图群体的亲本。赞引1号是从低纬度国家赞比亚引进选育的一份优良品系, 而福红952B是课题组通过杂交育种方法在亚热带春夏播条件下育成的品系, 两者来源差异远。且分期播种发现, 赞引1号光周期反应敏感度最低(36.0%), 福红952B光周期反应敏感度最高(56.2%)。因此, 可以用这两份材料作为构建分离群体的亲本, 用于基因定位。
不同的红麻品系在不同播期条件下, 短日照加快其发育速度, 导致开花提前。其光周期反应有一定差别, 赞引1号表现光周期反应钝感。在自然短日照条件下, 赞引1号与光周期反应敏感品系福红952B杂交组合中, 开花期性状受核基因控制, 光周期敏感对光周期钝感在杂交后代中以显性的方式表现出来, 存在1对主效基因控制, 主效基因的遗传率高。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|