


{kind=link}
{kind=link}
{kind=link}
新陆早棉花品种DNA指纹图谱的构建及遗传多样性分析
[聂新辉1, 2, * , 尤春源2, * , 李晓方3 , 秦江鸿2 , 黄聪1 , 郭欢乐1 , 王夏青1 , 赵文霞1 , 林忠旭1, *  ]
]
]
|
|


以新疆2013年前审定的51个新陆早常规棉花品种为材料, 从5000对SSR引物中筛选出多态性高、稳定性好、且定位在棉花26条染色体上(每条染色体上选择2~3对)的75对核心引物, 检测到多态性位点226个, 每个标记检测到的基因型位点数在2~12之间, 平均为3.01个; 引物多态信息量(PIC)值介于0.0799~0.8752之间, 平均值为0.6624。结果显示, 在51份新陆早棉花常规品种中, 可利用特征引物将21份品种一次性区分开。利用40对引物可以完全区分开新陆早51份常规品种, 并构建供试品种的指纹图谱。同时利用NTSYS-pcV2.10软件聚类分析表明, 51个新陆早棉花品种遗传相似系数变化范围为0.4269~0.9873, 平均值为0.7071, 说明新陆早棉花品种之间遗传多样性较狭窄; 遗传相似系数矩阵和聚类分析将51个新陆早品种分为4大类型, 与原品种选育系谱高度吻合。
Fifty-one Xinluzao conventional cotton varieties authorized before 2013 were detected with 75 pairs of primers with high polymorphism, good repeatability, and even distribution on 26 chromosomes (2-3 pairs from each chromosome) selected from 5000 pairs of SSR primers. A total of 226 polymorphic loci were obtained, and each marker detected 2-12 polymorphic loci with an average of 3.01; the polymorphism information content ranged from 0.0799 to 0.8752, with an average of 0.6624. The results showed that 21 varieties were differentiated by characteristic primers, and the rest 30 varieties could be identified by primer conbinations. In the end, 40 specific primers and combinated primers could completely differentiate the 51 Xinluzao cotton varieties from each other. DNA fingerprinting of the 51 Xinluzao conventional varieties were constructed with the 40 pairs of SSR markers. NTSYS-pcV2.10 software was used to analyze genetic diversity of the 51 conventional varieties, the results indicated that genetic similarity coefficient for the varieties ranged from 0.4269 to 0.9873, with an average of 0.7071, showing the narrow genetic diversity in Xinluzao cotton varieties. The 51 Xinluzao cotton varieties were divided into four types with the genetic similarity coefficient matrix and cluster analysis, which were strongly consistent with their pedigrees.
棉花是我国重要的经济作物, 优质、高产棉花品种的选育在棉花产业可持续稳定发展中发挥重要作用。随着棉花产业形势的发展, 新品种审定速度快、数量多, 而骨干亲本数量有限且反复利用, 导致品种间遗传差异变小; 随着转基因技术在棉花育种中的应用, 在原品种基础上改良少数或单个性状的衍生品种增多, 完全依据形态性状进行棉花品种的辨别越来越困难[ 1]。同时, 推向生产的新品种更新换代快、数量多, 导致种子市场“混、杂、乱”局面, 阻碍棉花产业的健康发展。传统的棉花品种真实性和纯度是通过田间种植的方法鉴定的, 耗时长、成本高、时效性差, 易受环境因素的影响[ 2]。
近年来, 随着分子生物学的不断发展, 分子标记技术因不受环境条件的影响且多态性标记可反映DNA水平上生物个体间的遗传差异而被广泛应用, 其中SSR标记被认为是品种鉴定较为理想的标记之一[ 3]。目前, 分子标记技术广泛应用在棉花品种指纹图谱构建和遗传多样性分析中, 例如, 郭旺珍等[ 4]最早在1996年利用RAPD技术对中国9个棉花主栽品种的指纹图谱进行了研究。Multani等[ 5]利用RAPD标记构建了澳大利亚13个陆地棉品种和1个海岛棉品种的指纹图谱。Iqbal等[ 6]对22个陆地棉品种和1个亚洲棉品种进行RAPD分析, 认为陆地棉品种的遗传基础很窄。武耀廷等[ 7]利用SSR标记检测了30个陆地棉栽培品种和4个杂交种亲本的多态性, 并建立了杂交种的指纹图谱, 用于它们的分子鉴定和纯度检测。马轩等[ 8]利用SSR技术建立18个彩色棉品系的DNA指纹图谱。姜伟等[ 9]利用ISSR标记, 对8个新疆品种(系) 构建指纹图谱。李武等[ 10]利用SRAP标记, 对中国引入海岛棉以来培育的36个国内品种及20个国外品种进行了遗传多样性分析。潘兆娥等[ 11]构建中棉所48的SSR数字指纹图谱。薛艳等[ 12]建立新疆早熟棉品种SSR分子标记体系。李成奇等[ 13]利用20对SSR核心引物构建了百棉系列棉花自交系品种(系)的DNA指纹图谱。孙宁等[ 14]以2009年度参加黄河流域国家棉花区试的57份参试品种为材料进行指纹图谱分析。匡猛等[ 15]采用SSR标记构建2008年中国3大棉区8个棉花主产省份32份棉花主栽品种的DNA指纹图谱并进行遗传多样性分析。赵亮等[ 16]利用26对SSR引物构建12个品种DNA条形码编制。与其他分子标记相比, 以微卫星序列为基础的SSR标记在DNA指纹鉴定上显示了独特的优越性, SSR 标记数量丰富, 覆盖整个基因组, 揭示的多态性高, 以孟德尔方式遗传, 呈共显性[ 2]。近年来, 基于SSR标记构建了水稻[ 17]、小麦[ 18]、玉米[ 19]、烟草[ 20]、大豆[ 21]等主要农作物的DNA指纹图谱数据库。
本研究拟通过对51份新陆早常规棉花品种指纹图谱的建立及品种间遗传关系分析, 为种子管理经营单位提供棉花品种快速、准确、科学的鉴定方法, 保护育种者品种的知识产权和育种家的权益, 也为育种者提供亲本有效选配、品种合理种植的参考依据, 加快优质丰产新陆早棉花新品种的选育。
2013年以前新疆审定的新陆早棉花常规品种51份, 统一由原品种选育单位提供(见附表)。SSR引物由华中农业大学作物遗传改良国家重点实验室提供, 引物合成、PCR试剂dNTPs、 Taq酶及电泳试剂等购自上海Sangon公司。
参照白静的棉花总DNA提取方法、SSR-PCR反应体系、PCR扩增程序、PAGE凝胶电泳及显色方法[ 22]。
根据PCR扩增产物在电泳凝胶上的相对位置, 对每对引物生成的不同基因型直接编号, 构建51份新陆早棉花品种的DNA分子指纹图谱。其中电泳结果采用0,1系统记录谱带位置, 某一扩增条带有带记为1, 无带记为0, 将每对引物在品种间扩增得到0,1 (二进制)数据转化成十进制数据, 该十进制数据代表每个引物扩增的结果, 则每40个引物组合的十进制数字串作为每一个品种的数字指纹。
各引物对应不同基因型扩增, 计算位点多态性信息含量(polymorphism information content, PIC), 按公式PIC = 1-Σ Pi2 计算, 其中 Pi指第 i个等位变异出现的频率。采用UPGMA (unweighted pair-group method with arithmetic mean, 未加权平均数)法进行聚类分析, 并绘制树状聚类图。利用NTSYS-pcV 2.10软件中的Qualitative data计算品种间的遗传相似系数(Jaccard系数)。
全面综合评价实验室已有的5000对引物, 选择多态性高、稳定性好、定位在棉花26条染色体上且分布均匀的标记引物91对, 在参试品种中进行筛选, 最终确定核心引物75对, 占检测引物的82.42%。
如 表1所示, 75对SSR核心引物扩增检测到多态性基因型位点226个, 每个标记检测到的基因型位点数介于2~12个之间, 平均为3.01个。其中基因型数目大于平均数的标记47个, 引物编号分别为2、3、4、5、7、10、11、12、13、14、16、17、20、21、22、23、24、25、26、27、30、36、37、38、39、40、41、42、43、 46、48、50、52、54、56、57、60、61、64、65、66、68、69、70、71、73和74。 图1所示, NAU3995标记在51份材料中扩增出等位基因数为4个。
表1所示, 标记间的多样性指数介于0.0799~ 0.8752之间, 平均多样性指数为0.6624, 其中多样性指数大于平均数的标记51个, 引物编号分别为1、2、3、4、5、7、8、9、10、11、12、13、14、15、16、20、21、22、23、24、25、26、27、30、31、32、34、36、37、38、39、41、42、43、44、45、46、48、50、53、54、56、57、60、61、62、64、65、66、70和71。表明这些标记具有较强鉴别品种的能力。
等位基因数和多样性指数均大于平均数的标记有40个, 如标记NAU3736, 等位基因数和基因型个数分别为5和12, 引物多态性信息值为0.8752。进一步证实挑选的核心标记引物有效性高, 标记的等位变异数多, 其多样性指数也高。
| 表1 75对SSR标记在51份新陆早棉花品种中的引物多态性信息 Table 1 Polymorphism information of 75 SSR loci in 51 Xinluzao cotton varieties |
 | 图1 标记NAU3995在51份新陆早棉花中电泳扩增等位基因特征Fig. 1 Alleles feature of marker NAU3995 in 51 Xinluzao cotton varieties |
采用75对引物对51个新陆早棉花品种进行指纹分析, 其中21个新陆早棉花品种具有特征引物。在一定材料范围内, 特征引物是指某个品种具有明显区别与其他品种特有指纹信息的引物, 采用这对引物就可以直接区分该品种与其他品种。 表2所示, 新陆早4号、新陆早10号、新陆早13、新陆早15、新陆早16、新陆早21、新陆早22、新陆早23、新陆早26、新陆早33、新陆早34、新陆早37、新陆早38、新陆早39、新陆早41、新陆早45和新陆早54分别具有1个特征引物, 新陆早3号、新陆早6号、新陆早40和新陆早47分别有3、3、6和2个特征引物。 图2所示, BNL3257标记是新陆早41的特征引物。而NAU3736标记可以一批次分别鉴别出新陆早13、新陆早16、新陆早22、新陆早38、新陆早39和新陆早47。
表3所示, 其余30份材料需要引物组合来区别该品种与其他品种。例如, 在51份新陆早品种中区分新陆早1号, 第一步加标记HAU2022, 利用扩增出的特征带型(指纹信息)从51份材料中首先区分出来新陆早1号和新陆早3号, 第二步加新陆早3号特征引物NAU3995, 可明显区分开新陆早1号和新陆早3号, 即选用2对引物HAU2022和NAU3995组合就鉴别出新陆早1号。
依据 表2和 表3, 综合如 表4所示, 最少选用40对特征引物和组合引物就可以完全区分开30份新陆早常规品种。因此, 可用这40对引物的指纹信息构建新陆早品种的指纹图谱。
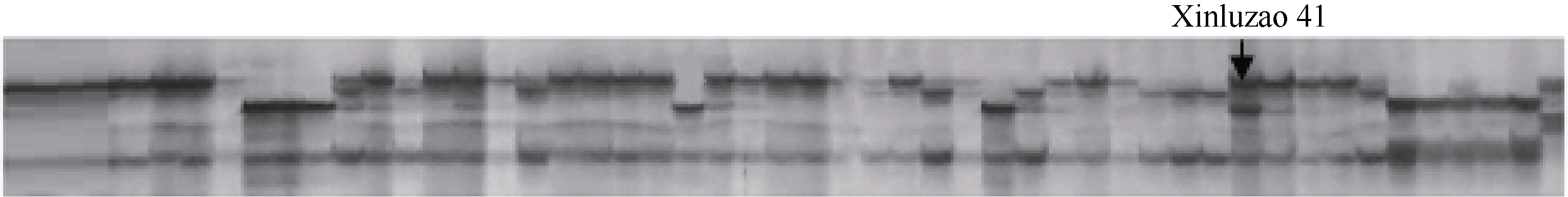 | 图2 新陆早41的特征引物BNL3257电泳扩增图Fig. 2 Electrophoresis profiles of specific marker NAU1362 for Xinluzao 41 |
| 表2 新陆早棉花品种的特征引物 Table 2 Specific amplification primers for Xinluzao cotton varieties |
| 表3 30份新陆早品种区分的组合引物 Table 3 Primer combinations for 30 Xinluzao cotton varieties |
| 表4 利用40对引物构建51份新陆早棉花品种的指纹信息 Table 4 DNA fingerprinting of 51 Xinluzao cotton varieties using 40 primers |
根据SSR分子标记统计不同基因型数据结果, 利用NTSYS-pcV2.10软件, 采用Jaccard系数计算51个新陆早常规品种遗传相似系数, 得到相似系数矩阵。结果表明, 51个新陆早品种遗传相似系数变化范围是0.4269~0.9873, 平均值为0.7071, 遗传相似系数较高, 表明新陆早棉花品种之间遗传多样性较狭窄。遗传相似系数最大的品种是新陆早30和新陆早31, 表明二者的遗传差异较小; 遗传相似系数最小是新陆早15和新陆早32, 表明两者遗传差异较大, 亲缘关系较远。
根据品种遗传相似系数矩阵, 对品种聚类( 图3), 在相似系数为0.4619处可以将51个新陆早棉花品种分为4大类, A类包括新陆早13个棉花品种, B类包括29个, C类包括2个, D类包括7个, 其中B类在遗传相似系数0.4778处又可分为2个亚类, 即B1与B2, B1包含16个品种, B2包含13个品种。对比各新陆早品种系谱可明显看出, A类以石河子棉花研究所(有新陆早2号、新陆早5号、新陆早7号、新陆早8号、新陆早10号、新陆早19、新陆早36、新陆早46)及农七师农业科学研究所(有新陆早3号、新陆早4号、新陆早6号)选育的品种为主, 其亲本多含有前苏联早熟陆地面血缘, 其中新陆早1号、新陆早6号、新陆早7号、新陆早8号、新陆早36在北疆早熟棉区作为主载品种推广面积较大。
B类以农七师农业科学研究所(新陆早9号、新陆早13、新陆早15、新陆早16、新陆早23、新陆早25、新陆早31、新陆早35、新陆早37、新陆早38、新陆早39、新陆早47和新陆早49)及农五师地区(新陆早11、新陆早12、新陆早37、新陆早54)所选育品种为主, 新陆早系列品种中, 优质中长绒或抗病性较好的品种主要集中在B类中。其亲本多含有贝尔斯诺、爱子棉、中棉所12、中棉所17及辽棉系列的资源, 且通过审定的中长绒品种品种主要都聚类在B2亚类中(有新陆早24、新陆早25、新陆早28、新陆早29、新陆早30、新陆早31、新陆早35、新陆早38、新陆早39、新陆早40、新陆早47、新陆早49)。其中新陆早12和新陆早13在北疆早熟棉区作为主载品种推广。
C类中的包含新陆早26、新陆早27, 其中新陆早26在北疆早熟棉区推广面积较大。
D类新陆早品种主要由新疆农垦科学院棉花研究所(有新陆早22、新陆早32、新陆早33、新陆早42、新陆早48、新陆早51、新陆早53)选育, 其中新陆早33和新陆早48在北疆早熟棉区为主载品种。新疆农垦科学院棉花研究所选育的新陆早45的亲本之一来源于新陆早13, 在分子标记多样性聚类分析中, 与新陆早13聚为一类和系谱描述高度吻合。
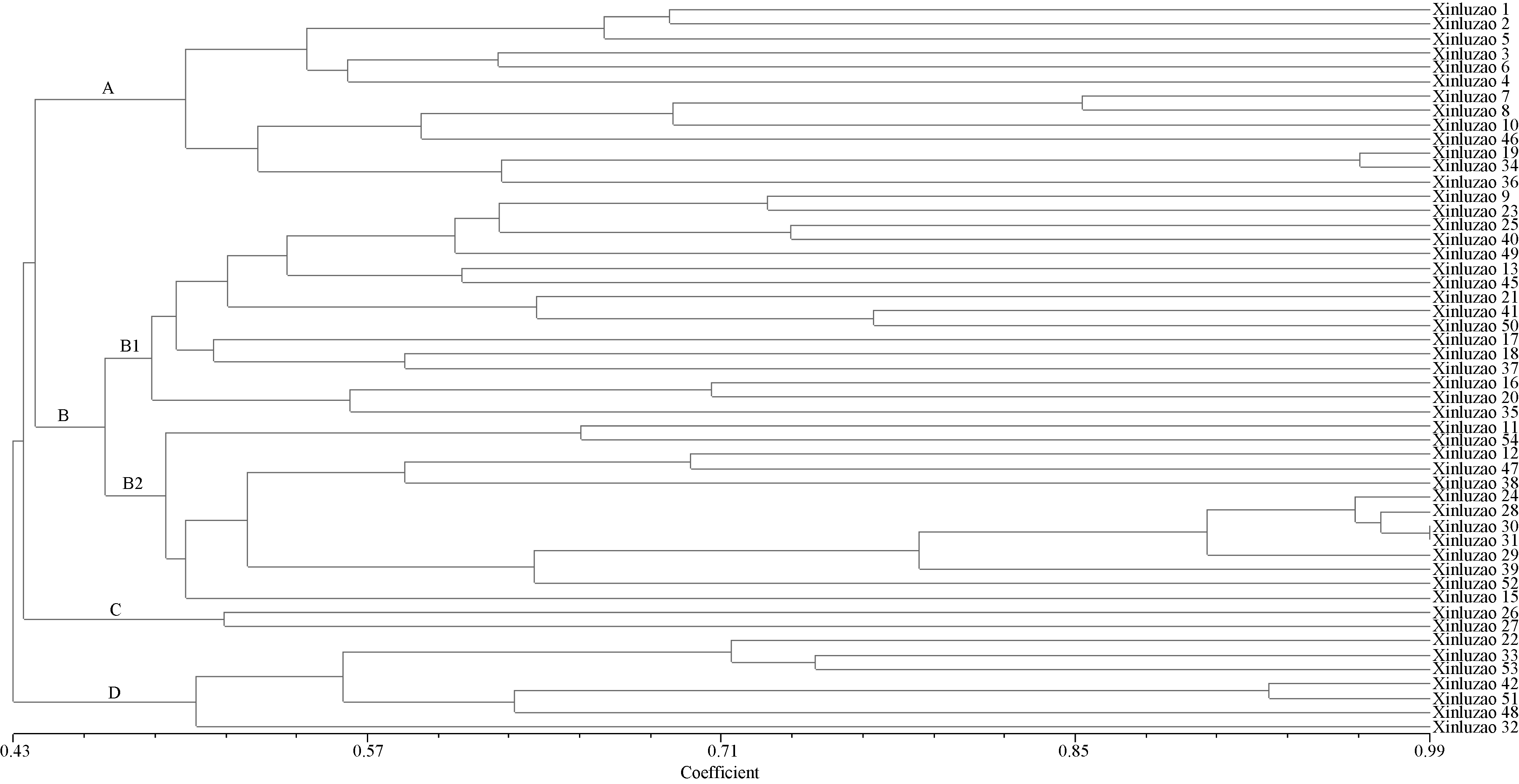 | 图3 51个新陆早棉花品种的聚类分析图Fig. 3 Clustering analysis of 51 Xinluzao cotton varieties |
随着棉花全基因组测序迅速进展, 构建高通量、准确性高, 易操作的全基因组覆盖DNA指纹库将越来越受到重视。一种理想的分子标记应具有以下特点, 即标记引物的多态性高、重复性和稳定性好; 带型清晰, 易统计; 染色体上均匀分布; 共显性; 开发和使用成本低。DNA指纹技术的发展经历3个阶段, 第1代是以Southern杂交为基础的RFLP, 第2代是以PCR为基础的DNA指纹标记(如RAPD、AFLP、SSR、SCAR、ISSR等), 第3代是以单核苷酸多态性为基础的SNP[ 23]。近些年, 分子标记开始应用于各物种的鉴定, 国际植物品种权保护组织(UPOV)在BMT测试指南草案中已将构建DNA指纹数据库的标记确定为SSR和SNP[ 24]。随着第2代测序技术的发展, 使品种基因组DNA测序和SNP全面分析成为可能, 从而大大提升了品种识别和鉴定精度, 但检测成本较高, 有时会因检测尺度的过度精细, 把品种内的差异误判成品种间的差异[ 25]。因此, SNP高额的投入使其未能得到广泛应用[ 26], 其中SSR标记技术比较成熟, 成为当前各个作物构建指纹数据库的首选技术。本研究采用SSR技术构建了新陆早常规品种指纹图谱, 并分析和研究了品种的遗传多样性和亲缘关系。
本试验的聚丙烯凝胶电泳采用变性胶, 而薛艳等[ 12]研究42份新疆陆地早熟棉指纹图谱构建与品种鉴别时采用9%非变性聚丙烯酰胺凝胶电泳分离方法, 曲鲁江等[ 27]报道的非变性凝胶中条带过于复杂, 出现的非特异性条带极大地影响了分析带形的准确性, 使试验结果的误差加大, 变性胶在利用银染检测微卫星产物时条带清晰, 带型分析准确性更高, 结果更加可靠。
本研究以75对核心引物SSR标记扩增检测到多态性基因型位点226个, 每个标记检测到的基因型位点数在2~12之间, 平均为3.01个, 其中等位基因型数目大于平均数的标记47个; 标记间的多样性指数介于0.0799~0.8752之间, 平均数为0.6624, 多样性指数大于平均数的标记51个。张玉翠等[ 2]用32个主栽品种, 40对引物, 共扩增出161种多态性基因型, 引物多态信息量(PIC)为0.2989~0.7585, 平均为0.5407; 艾先涛等[ 28]用94份新疆陆地棉品种, 54对具有稳定多态性的引物, 共检测出153个多态性位点, 每对引物的等位变异为2~6个, 平均为2.93个; 引物多态信息含量(PIC)为0.0430~0.6640, 平均为0.3831。与其相比, 本研究筛选的引物标记是高效的。
51个新陆早棉花品种采用75对引物进行指纹分析, 其中21个品种具有特征引物, 获得特征引物为23个, 其中NAU3736标记可以一批次分别鉴别出6个品种, 而引物NAU3736在 表1中, 等位基因数、基因型个数和PIC值分别为5、12、0.8752, 结果表明引物多态性越丰富且特征谱带数量越多的标记引物其鉴别品种能力越强, 可以首选作为构建新陆早棉花品种指纹图谱的标记。其余30份材料没有获得特征引物, 采用引物组合来区别该品种与其他品种, 共选用组合引物34个。最终统计, 至少确定40对特征引物和组合引物就可以完全区分开新陆早常规品种。以上研究表明, 特征引物在一定材料范围内鉴定是最快捷、易见的, 但是只有部分品种具有特征引物, 随着鉴定材料数量的扩大, 需要采用组合引物鉴定, 可大幅度提高鉴别能力。本研究中全面、系统构建的新陆早51个品种指纹图谱, 每一个品种的指纹都是唯一的, 具有很强的特异性, 此数据库可以用于新陆早棉花品种初级数据库。与赵亮[ 16]等的研究具有明显差异, 本试验选用新疆2013年前审定的早熟陆地棉品种全套完整材料, 75对核心标记均匀分布于棉花26条染色体且较高多态性, 结合特征引物和组合引物方法构建品种指纹。
51个新陆早品种遗传相似系数矩阵和品种聚类表明, 新陆早棉花品种之间遗传多样性较狭窄, 这与艾先涛等[ 28]的研究结果相似, 说明新疆陆地棉品种间遗传关系相对简单, 品种的遗传基础相对狭窄, 品种遗传组分差异较小, 总体上遗传多样性不够丰富。在相似系数0.4619处将51个新陆早品种分为4大类, 其中每类包括的新陆早品种基本是来源于一个地区育种单位, 例如, A类包括石河子棉花研究所和农七师农业科学研究所选育品种, B类包括农七师农业科学研究所和农五师农业科学研究所选育品种, C类是2个公司选育品种, D类包括新疆农垦科学院棉花研究所选育品种, 具有相同来源的品种最先聚为一起; 遗传相似系数最大的品种是新陆早30和新陆早31, 表明其遗传差异较小, 结合 表3显示, 新陆早30和新陆早31分别用标记BNL119、BNL1421组合和标记DPL238、BNL2486、NAU3995、CIR170、BNL1317组合引物鉴别, 因此遗传差异较小的品种使用组合引物标记鉴别才更为准确。以上研究结果与Liu等[ 29]的观点一致, 即SSR聚类结果与地理分布一致, 遗传相似系数揭示种群之间的正相关关系; 从附表看出, 新陆早9号、新陆早16、新陆早25、新陆早31和新陆早39的亲本均来源于贝尔斯诺系统, 因此B类品种主要亲本来源有贝尔斯诺、爱字棉系统、中棉所12、中棉所17、辽棉系列, 此系列选育品种数量最多、棉花纤维品质较优, 说明聚类结果和品种根据实际系谱来源分类是一致的, 这与张玉翠等[ 2]的研究结果一致, 即遗传相似系数聚类分析结果基本上反映了品种之间的亲缘关系。新疆北疆早熟陆地棉品种间分子聚类结果与品种本身遗传系谱背景高度吻合, 符合品种本身的真实特性。具有相近表型性状、相同的遗传背景和相同育种单位选育的品种聚在同一类群中。说明北疆不同育种单位在早熟棉选育过程中积累形成了各自的特点及优势, 表现在各育种单位在基础种质资源上确立了其特定的选育方向。
今后应将形态信息与分子数据结合一起形成指纹图谱, 使品种保护信息更加全面、准确。目前已在油菜[ 30]、大豆[ 31]构建分子身份证, 在甜瓜[ 32]上建立指纹图谱QR编码, 在水稻[ 33]上提出基于SSR分子指纹和商品信息构建品种身份证的新思路。因此, 棉花DNA指纹鉴定技术发展将不断完善, 将棉花的指纹信息编码结合品种的基本商品信息和特异基因信息构建棉花品种身份证。
利用SSR分子标记技术, 通过40对特征引物和组合引物构建了新陆早51份棉花品种指纹图谱, 将51个新陆早品种聚为4大类型, 遗传多样性较狭窄。构建的指纹图谱可以作为新陆早棉花品种初级分子标记指纹数据库。
致谢: 本研究的试验材料来自各品种育种单位提供的原原种, 主要由新疆农垦科学院李保成和余渝, 石河子棉花研究所孔宪良、黄顶元及秦江鸿, 农七师农科所李家胜和赵富强, 农五师农科所曹阳提供; 分子标记实验由作物遗传改良国家重点实验室林忠旭教授精心设计指导, 同时得到黄聪、郭欢乐、王夏青等研究生的帮助, 在此表示真诚的感谢。
| 附表 新陆早棉花品种的信息 Supplementary table Informations of Xinluzao cotton varieties |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|