
{kind=link}
水稻穗部性状的QTL分析
[吴亚辉, 陶星星, 肖武名*  , 郭涛, 刘永柱, 王慧, 陈志强
, 郭涛, 刘永柱, 王慧, 陈志强* ]
, 郭涛, 刘永柱, 王慧, 陈志强]
|
|


水稻穗部性状与产量关系密切。以粳稻品种日本晴和一个大穗籼稻材料H71D为亲本构建F2群体, 于2011和2012年分别以172个单株和138个单株, 对穗长、一次枝梗数、二次枝梗数、总粒数、穗着粒密度等5个穗部性状进行QTL定位。以LOD≥3为阈值, 两年共检测到38个QTL, 其中2011年21个, 2012年17个, 2年重复检测到的QTL为4个, 占总数的10.5%。5个性状之间大都具有显著的表型相关性, 相关性较强的性状之间具有较多相同或紧密连锁的QTL, 效应值较大的QTL易于在不同群体和不同环境中被重复检测到。检测到的QTL为进一步进行元分析和精细定位打下了基础, 也为通过分子标记辅助选择提高产量提供了有用信息。
Rice yield potential is closely related to panicle traits. QTLs for five panicle traits including panicle length, number of primary branches, number of secondary branches, spikelets per panicle and seed setting density were identified by using 172 plants and 138 plants from a F2 population derived from a cross between Nipponbare and H71D in 2011 and 2012 respectively. A total of 38 QTLs were detected in the two trials, including 21 QTLs in 2011 and 17 QTLs in 2012, of them four QTLs (only 10.5%) were repeatedly detected in both years. Some QTLs controlling different traits shared the same maker interval on the chromosome with each other, which was consistent with their significant phenotypic correlations. QTLs with large effects are easily to be detected in kinds of populations and different environments. These QTLs provide useful information for meta-analysis and fine mapping, as well as MAS for high-yield rice breeding.
稻穗是水稻产量的最终体现部位, 穗部性状在产量结构中起着重要作用。水稻穗形相关性状包括穗长、一次枝梗数、二次枝梗数、总粒数、穗着粒密度等。穗着粒密度是总粒数和穗长之比。研究穗形相关性状对选育穗长适中、枝梗数分布合理、着粒密度适当增加的理想品种具有重要意义[ 1, 2, 3]。
水稻穗部相关性状QTL (quantitative trait locus)的定位研究已经有很多报道, 不同研究者利用各种群体定位的结果不尽相同。郭龙彪等[ 4]利用水稻汕优63 (珍汕97×明恢63)重组自交系群体241个株系, 对9个重要农艺性状进行年度间的QTL定位, 2年共检测到64个QTL, 其中2年均检测到的穗长、每穗总粒数、每穗实粒数QTL均只有1个。叶少平等[ 5]利用以培矮64S和Nipponbare为亲本构建的F2群体, 对单株穗数、每穗粒数、每穗实粒数、结实率、着粒密度、千粒重6个产量构成性状进行QTL定位, 共检测到22个QTL, 其中对每穗粒数、每穗实粒数、着粒密度3个性状分别检测到7、3和4个QTL。程桂平等[ 6]以广陆矮4号为母本及轮回亲本, 普通野生稻( Oryza rufipogon)为父本, 构建的BC2F2群体, 对14个产量相关性状进行QTL定位, 共检测到分别控制穗长、每穗总粒数、每穗实粒数、穗着粒密度的QTL为3、3、1和2个。由此可见, 控制水稻穗部性状的基因座具有较广泛的遗传多样性, 而且其遗传表达易受环境条件的影响。因此, 利用不同的定位群体在不同条件下对水稻穗部性状进行QTL分析, 有望鉴定出一些稳定表达的或者新的基因座位, 从而丰富水稻穗部性状研究的理论基础。
本研究利用一个来源于日本晴/H71D的F2群体, 分别于2011年和2012年对172个单株和138个单株的穗长、一次枝梗数、二次枝梗数、总粒数、穗着粒密度等5个性状进行QTL分析, 并与前人的研究结果进行比较, 这将有助于加深我们对水稻穗部性状遗传多样性的认识, 并为进一步的元分析以及利用分子标记辅助选择聚合水稻穗部性状基因提供一定的理论依据。
以粳稻日本晴(Nipponbare)为母本、大穗籼稻品种H71D为父本进行杂交, 于2010年早造配制Nipponbare/H71D杂交组合, F1自交后分别于2011年晚造和2012年晚造种植成F2群体。
分别于2011年和2012年晚造于华南农业大学校内农场按宽窄行(窄行20 cm、宽行40 cm)种植日本晴、H71D以及F2群体, 单本插。田间水肥管理同常规大田。
成熟后取主穗按申宗坦[ 7]的方法考种。考察5~10株亲本的主穗, 取平均值, 2011晚造和2012晚造分别随机取172个和138个F2单株的主穗, 考察穗长(panicle length, PL)、一次枝梗数(number of primary branches, PB)、二次枝梗数(number of secondary branches, SB)、每穗总粒数(spikelets per panicle, SPP)、穗着粒密度(seed setting density, SD)等, 其中穗着粒密度是每穗总粒数与穗长的比值, 单位为粒 cm-1。遵循McCouch等[ 8]的方法命名QTL。
从已发表的408对SSR引物中选取123对多态性较好的引物, 参考Cornell SSR 2001及Gramene上(http://www.gramene.org/)公布的其他遗传图谱调整部分引物的顺序(籼粳杂交标记偏分离比较严重, 而偏分离会影响标记的排序[ 9, 10, 11]), 利用IciMapping 3.2[ 12]构建覆盖水稻基因组1927 cM的遗传图谱, 标记间平均距离为15.7 cM。采用ICIM-ADD方法对5个穗部性状进行QTL定位分析, 取LOD=3.0为阈值检测QTL存在与否。若LOD≥3.0, 则认为该区间LOD值最高处所对应的位点即为该性状的1个QTL, 同时计算出每个QTL的加性效应和对性状的贡献率。
2011年和2012年F2群体及其双亲5个穗部性状均存在显著或极显著性差异(表1), 表明2个亲本的遗传差异较大, 有利于QTL定位。除一次枝梗数外, F2其他4个性状均出现不同程度超亲分离; F2群体各性状的偏度和峰度的绝对值都小于或约等于1, 说明这些性状都呈正态分布, 且均为多基因控制的数量性状。
2011年晚造F2群体的总粒数与穗长、一次枝梗数的相关性不显著, 穗长与穗着粒密度呈负相关, 其他性状间则呈极显著正相关; 2012年晚造各性状间均为极显著正相关(表2)。这说明穗部各性状间的关系极复杂, 同时容易受环境的影响。
2.3.1 2011年穗部性状QTL的定位结果 2011年共检测到影响5个性状表现的21个QTL (表3), 分布于除了第4和第9染色体外的其他10条染色体上, LOD值介于3.04~6.94。这些QTL解释的表型变异贡献率变幅为7.17%~25.67%, 其中表型变异贡献率大于10%的QTL有18个。共检测到2个穗长QTL, 均位于第12染色体上, qPL-12-1和 qPL-12-2两个位点的LOD值分别为3.19和3.04, 贡献率分别为15.85%和15.99%。两个穗长QTL的加性效应均为负值, 增效等位基因均来自H71D。共检测到6个一次枝梗数QTL, LOD值范围为3.04~6.17, 其中 qPB-3-1和 qPB-10-4两个QTL的增效等位基因来自于Nipponbare, 其他4个QTL的增效等位基因均来自H71D。共检测到4个二次枝梗数QTL, 增效等位基因均来自H71D, 总的表型变异贡献率高达76.15%。共检测到4个总粒数QTL, 增效等位基因也都来自H71D, 其中 qSPP-10-1的表型变异贡献率高达25.67%, qSPP-11-1的加性效应高达-45.6粒。共检测到5个穗着粒密度QTL, 其增效等位基因也都来自H71D, 表型变异贡献率变幅为9.30%~24.58%, 加性效应变幅为-0.56 ~ -1.51粒 cm-1。
| 表1 亲本及F2代群体穗部性状的表现 Table 1 Phenotypic performance of panicle traits between parents and the F2 population from the cross of Nipponbare/H71D |
| 表2 各性状间的相关性 Table 2 Correlation coefficient between observed traits in the F2 population |
2.3.2 2012年穗部性状QTL的定位结果 2012年共检测到5个性状的QTL为17个(表3), 分布于6条染色体上, LOD值介于3.06~5.99, 解释的表型变异贡献率为9.29%~52.62%, 其中表型变异贡献率大于10%的QTL 16个。检测到影响穗长的QTL只有位于第6染色体上的 qPL-6-1, 其LOD值为3.23, 贡献率为15.22%。2012年没有检测到影响一次枝梗数的QTL。共检测到4个二次枝梗数QTL, 其增效等位基因均来自H71D, 其中 qSB-1-2和 qSB-10-1的表型变异贡献率分别为25.02%和21.35%。共检测到5个总粒数QTL, 其增效等位基因都来自H71D, 其中 qSPP-10-1的表型变异贡献率高达52.62%, 加性效应高达-64.29粒。共检测到7个穗着粒密度QTL, 除 qSD-7-1外增效等位基因全部来自H71D, 表型变异贡献率变幅为10.57%~45.33%, 加性效应变幅为-2.06~0.68粒 cm-1。
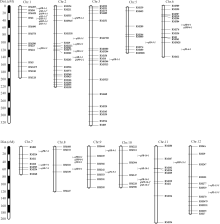
2.3.3 2年定位结果的比较 2年共检测到38个QTL, 其中2011年21个, 2012年17个, 2年间重复检测到的QTL有4个(表3和图1), 占总数的10.53%。
2011年共检测到2个穗长QTL, 2012年只检测到1个穗长QTL, 2年中没有重复检测到的穗长QTL; 2011年共检测到6个一次枝梗数QTL, 但在2012年没有检测到一次枝梗数QTL, 可能与LOD阈值的设定有关, 同时表明一次枝梗数受环境影响较大; 2年均检测到4个二次枝梗数的QTL, 其中 qSB-1.1被重复检测到, 且2年的贡献率均大于10%; 2年共检测到控制穗粒数的QTL有8个, 其中重复检测到的 qSPP-10-1在2011年和2012年的贡献率分别高达25.67%和52.62%; 2年共检测到控制穗着粒密度的QTL 12个, 重复检测到的QTL有2个( qSD-1-1和 qSD-10-1), 两者的贡献率均较大, 2012年检测到的 qSD-7-1和 qSD-7-2的增效等位基因来自于Nipponbare。第1染色体上的 qPB-1-1、 qSB-1-1、 qSPP-1-1和 qSD-1-1均在分子标记RM495~RM84区间内, 该区间可能存在一个同时控制一次枝梗数、二次枝梗数、总粒数和穗着粒密度的QTL; 第10染色体上的 qPB-10-2、 qSPP- 10-1、 qSD-10-1等3个QTL均位于区间RM147~ RM228, 表明该位点可能存在一个效应值较大的多效性主效QTL。
| 表3 2年间5个穗部性状的QTL定位结果及遗传参数估算 Table 3 Identification of QTLs for five panicle traits in F2 population and their genetic parameters estimated in 2011 and 2012 |
 | 图1 2年中检测到的穗部性状QTL在染色体上的位置*表示2年重复检测到的QTL。Fig. 1 Location of the QTLs for panicle traits detected in 2011 and 2012 in the genetic map* Indicates the QTL detected in two years. |
前人的很多研究表明, 数量性状极易受环境的影响, 控制同一性状的QTL在不同定位群体中有不同的表现, 同一群体在不同环境、不同年份的检测结果往往也会有所不同[ 4, 5, 6]。本研究也发现同一群体不同年份间检测到的QTL差异是很明显的。2年共检测到5个性状的38个QTL, 其中重复检测的只有
4个, 重复率分别为19.0%和23.5%, 其2年间的贡献均大于10%, 表明这4个QTL在不同年份和条件下变化较小, 这些对环境影响不敏感且效应值较高的QTL对分子标记辅助选择育种具有较大价值。
QTL的多效性指的是同一区间的QTL同时对多个性状产生作用, 这种情况在动植物中普遍存在[ 13, 14, 15, 16]。本研究的QTL定位结果也证实QTL多效性的存在。如第1染色体上的 qPB-1-1、 qSB-1-1、
qSPP-1-1和 qSD-1-1均在分子标记RM495~RM84区间内, 该区间可能存在一个同时控制一次枝梗数、二次枝梗数、总粒数和穗着粒密度的QTL; 还有第10染色体上的 qPB-10-2、 qSPP-10-1、 qSD-10-1等3个QTL均位于区间RM147-RM228, 表明该位点可能存在一个效应值较大的多效性主效QTL。这些集中分布的QTL与性状间的相关性是一致的。这种QTL成簇分布的现象在遗传上通常表现为一因多效或基因的连锁或基因重叠, 可能是一些具有多效性的未知关键因子同时控制着多种代谢途径, 从而使相关性状在自然选择过程中形成相互适应的有利性状[ 17], 要探究其真正原因还需要通过构建近等基因系进行精细定位和克隆来进一步分析。
目前穗部性状的研究已经有很多。邢永忠等[ 18]通过珍汕97/明恢63的重组自交系群体对5个穗部性状进行QTL分析, 以LOD≥2.3为阈值分别检测到10、3、6、8、7个每穗颖花数、每穗实粒数、结实率、穗长和穗着粒密度QTL。李绍波等[ 19]在大田直播条件下, 利用来源于“Lemont/特青”的重组自交系群体, 对6个穗部性状进行QTL分析, 以LOD≥2.0为阈值检测到19个QTL, 其中控制穗长、一次枝梗数、二次枝梗数、每穗颖花数、着粒密度的QTL分别为4、2、4、2和4个。沈希宏等[ 20]通过对281个株系组成的超级杂交稻协优9308重组自交系群体2年的数据对7个穗部性状进行QTL检测, 以LOD≥2.5为阈值检测到52个QTL, 其中包括7个控制穗长的QTL, 8个控制一次枝梗数的QTL, 9个控制二次枝梗数的QTL, 6个控制着粒密度的QTL, 7个控制每穗总粒数的QTL。王智权等[ 21]以Asominori/IR24构建的染色体片段代换系于2007和2008年对穗部性状进行定位, 以LOD≥2.0为阈值共检测到5个性状的33个QTL, 其中2年同时检测到的稳定表达的QTL有10个。通过BioMercator V3.1整合部分已发表的研究结果(目前收集到79份图谱, 共600个QTL, 详细数据未列出), 发现本研究的结果与前人的研究结果有很多相同之处。如第1染色体上RM490附近的QTL簇在刘头明[ 22]的研究中有过报道, RM428附近的 qSPP-1-2在叶少平等[ 5]的研究中也有过报道; 在第6染色体RM584和RM136附近的QTL簇在沈希宏等[ 20]、刘头明[ 22]以及Marathi等[ 23]的研究中均有过报道; 第7染色体上的 qPB-7-1在黄成[ 24]的研究中有过报道。
本研究2年间重复检测到的4个QTL表型变异贡献率均高于10% (表3)。其中2个QTL qSPP-10-1和 qSD-10-2已经被报道, qSPP-10-1在叶少平等[ 5]、Mei等[ 25]、Jing等[ 26]和Bai等[ 27]的研究中表型变异贡献率分别为11%、11%、31%和14%, qSD-10-2在叶少平等[ 5]和Jing等[ 26]的研究中的表型变异贡献率分别为13%和22%。这2个效应较大的QTL位于同一位点, 该位点可能存在一个同时控制每穗粒数和穗着粒密度的多效性QTL, 可以考虑构建次级作图群体对这些效应较大的QTL进行精细定位。其他2个重复检测到的QTL ( qSB-1-1和 qSD-1-1)暂时未见于收集到的文献资料, 这些位点是否是本研究检测到的新位点还有待进一步整合更多已报道的位点并进行元分析。
本研究构建了覆盖水稻基因组1927 cM的遗传图谱, 标记间平均距离为15.7 cM。对穗长、一次枝梗数、二次枝梗数、总粒数、穗着粒密度等5个穗部性状进行QTL定位分析, 2年共检测到38个QTL, 包括2011年的21个, 2012年的17个。2年重复检测到的QTL有4个, 占总数的10.5%, 表型变异贡献率较大的QTL受环境影响较小, 较容易被重复检测。5个性状之间大都具有显著的表型相关性, 相关性较强的性状之间具有较多相同或紧密连锁的QTL。与前人的研究结果相比, 本研究中检测到的部分穗部性状QTL已经有过报道, 其中不乏2年均检测到且效应非常大的QTL (如第10染色体上的 qSPP-10-1和 qSD-10-2), 可以考虑构建次级作图群体对这些QTL进行精细定位。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|