

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻抽穗期高温胁迫对不同品种受粉和受精作用的影响
[张祖建1 , 王晴晴2 , 郎有忠1 , 王春哥1 , 朱庆森1 , 杨建昌1, *  ]
]
]
|
|


水稻抽穗期高温障碍导致结实性降低已经成为长江中下游水稻的重要自然灾害之一。以现代常规粳稻、三系杂交籼稻和两系杂交籼稻为试料, 研究了抽穗期高温处理对现代水稻品种和组合受精率的影响。现代粳型品种表现出较好的抽穗期高温耐性; 杂交稻特别是两系杂交稻抽穗期高温耐性相对较弱, 抽穗期39℃的高温胁迫下, 受精率接近于零; 在杂交稻中, 汕优63在较轻高温胁迫下表现较好耐性, 但在较强的高温胁迫下高温耐性迅速下降。相同处理下抽穗期高温耐性在品种间有较大差异。2个两系超级杂交稻的单药花粉数均在1750粒以上, 数量最多, 宁粳1号数量最少, 为每花药979.8粒。总体趋势为杂交稻多于常规稻, 籼型品种多于粳型品种, 分析表明单药花粉数与高温耐性不相关。高温胁迫导致花药开裂系数显著下降, 宁粳1号在较强的高温胁迫条件下还能维持一定程度的纵裂率, 开花后柱头上花粉数量相对较多。可见水稻抽穗期高温耐性与花粉生产能力无关, 与花药开裂和柱头捕获的花粉量关系较大。
High temperature stress at heading stage is one of the important natural disasters causing the reduction of the seed setting rate of rice in the middle-lower reaches of Yangtze River. This study investigated the effects of high temperature stress at heading stage on fertilization rate of modern rice varieties and combinations, including modern inbred
水稻抽穗期高温障碍(也称为热害)是指水稻在抽穗开花期内35℃以上高温胁迫引起颖花授粉和受精不能正常进行, 受精率严重下降, 导致水稻大幅度减产的现象, 在国内外早已有诸多报道[ 1, 2, 3, 4, 5]。水稻抽穗期高温障碍具有发生突然、成灾面积大、常造成严重减产等特点[ 5, 6, 7]。全球热带地区水稻抽穗期受到高温潜在威胁的面积约有4.0×106 hm2, 我国长江流域也是水稻花期高温危害的重灾区, 湖南、湖北、江西、江苏、四川等地均有水稻花期高温灾害的报道, 受灾严重时不少品种大面积结实率降低到50%以下, 损失惨重[ 8, 9, 10, 11, 12, 13, 14, 15]。近年来, 全球气候变暖愈加明显, 水稻抽穗期高温危害的发生呈现出日益加剧的态势, 对我国特别是长江流域水稻的安全生产形成了严重威胁[ 6, 16]。日本的有关模拟结果显示, 日本西南暖地地区在CO2浓度倍增条件下, 水稻产量因高温不育将减产8%~38%。Matsui等[ 17, 18]认为, 气温升高和CO2浓度上升两者对水稻花期造成的不育危害还有互作加重效应。
Prasad等[ 19]采用常温和常温+5℃对比试验的结果显示, 所有14个供试水稻品种的产量均显著降低。日本学者Satake等[ 20]根据不同类型品种对高温的反应, 将临界热害温度在32~34℃的定为敏感型品种, 35~37℃的定为中间型品种, 38~40℃的定为耐热型品种。我国早期研究认为籼稻开花长期高温伤害的临界温度为30℃, 短期高温伤害的临界温度为35℃[ 1]。国际水稻研究所的相关研究认为高温障碍指标为35℃[ 21]。徐云碧等[ 22]认为抽穗后3 d的平均最高温度超过35℃是早稻结实率明显下降的临界指标。其他还有不少水稻开花对高温敏感的时期及其障碍温度指标的报道, 多数结果认为32~37℃以上的持续高温即可引起受精的障碍[ 13, 23, 24]。已有的研究以籼稻为多, 有关我国粳稻类型的相关研究较少, 江苏省当前常用的水稻品种以粳稻为主, 其抗热性能并不清楚, 特别是近年来培育成功的超高产水稻材料的耐热性特征亟待判明。
有关花期热害的形成机制, 有研究指出, 品种间抽穗期耐热性的差异与柱头上的授粉数量呈显著正相关[ 25, 26], 同时还与花药结构、花粉产量、裂药特性等有关[ 25, 27, 28, 29]。张桂莲等[ 30]发现, 水稻抽穗开花期高温导致水稻开颖角度、花丝长度和花粉粒直径明显增大, 花药开裂、花粉可染率明显下降, 而柱头活力下降不明显。然而花期高温障碍机理的解析多局限于现象的观察, 少有直接因果关系的证明。
本文选用当前应用的常见籼、粳稻品种和杂交稻的代表组合, 从花粉行为入手解析当代籼、粳稻品种和三系、两系杂交稻抽穗期高温胁迫下的表现特点, 探讨水稻花粉行为的品种类型特征及其与抽穗开花期耐热性形成的关系。以期为解决两系杂交稻等新类型水稻受精率不稳定的难题提供新的思路, 为提高当代品种特别是超级稻品种的抗灾能力提供必要的理论积累, 也可为新型高产稳产超级水稻的选育提供一定的参考。
2005—2007年在扬州大学农学院盆栽实验场种植杂交籼稻汕优63和扬两优6号、常规籼稻扬稻6号、常规粳稻武香粳14、宁粳1号。5月5日播种, 大田育秧, 6月4日移栽至盆钵。盆钵高30 cm, 直径25 cm, 内装沙壤土。土壤含有机质24.5 g kg-1、全氮112.6 mg kg-1、速效磷58.0 mg kg-1和速效钾66.8 mg kg-1。每盆栽植3穴, 每穴1苗。移栽前每盆施1 g纯氮(以尿素溶液定量施入, 下同)、0.3 g P2O5 (以过磷酸钙施入)和0.5 g KCl做基肥; 在分蘖中期每盆施0.5 g纯氮, 穗分化期每盆施0.8 g纯氮。水层灌溉管理, 其余管理按常规栽培。
在控温自然光人工气候室中实施高温处理。以全盆20%茎蘖出穗为处理始期, 当天早晨搬入人工气候室, 连续处理4 d。搬入后每天早晨对当日开始开花的穗挂牌, 标记相应单茎的始花日。每天挂牌数量均在20穗以上, 以保证不同抽穗日成熟期受精率考种穗样在15个以上。
高温处理设置为37℃和39℃两个水平(气候室温控精度为±0.5℃)。每天高温处理时间为8 h (上午9:00至下午17:00), 高温处理时段, 气候室空气湿度控制在80%。其他时间为常温通风状态。
1.3.1 花药中花粉数的测定 对自然生长状态下的稻株取样观察。早晨选取预计当日开始开花的穗, 每材料取6穗, 用镊子轻取固定粒位(上3个一次枝梗的基部3粒)的9个颖花, 立即固定于50%酒精中待测。
按照染色→去除药壁→排列→计数的步骤[ 31]。选取测定颖花内颖侧最里侧的花药, 用镊子小心夹断花丝, 把花药置载玻片上, 在显微测微尺下量出花药的长度和宽度。加一滴1% I-IK溶液, 染色5 min至花药转为紫黑色。为防止操作过程中过快干涸, 在液滴中加入少许甘油。在解剖镜下用解剖针破开染色的花药, 去除药壁组织。待载玻片上的溶液蒸发至适宜时, 用解剖针将其上花粉粒排成“S”型, 于显微镜下计数。浓染花粉为充实花粉粒, 染色浅的和未染色的花粉为充实不良或未充实花粉粒。
1.3.2 花药开裂率 取当日即将开放的小花5朵(从稻穗上3个一次枝梗的基部3粒中选取), 共取2个穗子上的10个颖花, 在上午8:00取籼稻和杂交稻、10:00取粳稻样。用镊子将颖壳剥掉, 将花药置垫有潮湿滤纸的培养皿中, 于恒温箱培养5 h (25℃, 相对湿度为70℅), 然后在解剖镜下观察花药的开裂情况。将其区分为孔裂、纵裂两类。
1.3.3 柱头花粉数的观察 在观察花药开裂情况的同一天, 于开花后3~4 h (对籼稻和杂交稻为14:00—15:00, 对粳稻为16:00—17:00), 取分布于2~4个稻穗上的10朵当日开的颖花固定于FAA固定液。用苯胺蓝染色, 借助荧光显微镜观察柱头上花粉粒的数目。
高温处理的4 d期间, 按抽穗日期分别考测不同抽穗日稻穗强势粒的受精率。按顾世梁等[ 32]方法选择强势粒: 除顶部第2粒外直接着生于穗顶部4个一次枝梗上的颖果为强势粒。用透光法(借助垩白粒仪)鉴定受精与否, 子房有明显伸长者即为受精。
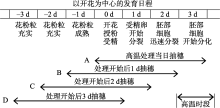
本试验以高温处理4 d期间不同日期开始开花稻穗的强势颖花为调查对象, 其受高温胁迫的发育阶段如图1所示, 通过这一方法评价高温胁迫对颖花花粉充实、授粉、发芽和受精过程等开花前后不同发育阶段的影响。抽穗开花越迟者, 其强势颖花受高温胁迫作用的发育阶段越早, 由此形成了4个在颖花开花前后的不同阶段的高温胁迫处理A、B、C、D, 高温胁迫的生育阶段依次向前推移。由此得到对颖花开花前后各发育阶段高温胁迫反应的初步比较。
 | 图1 高温处理期不同抽穗日形成的开花前后不同阶段高温胁迫处理Fig. 1 High temperature stresses around flowering stage based on different days of heading in treatment durations of high temperature |
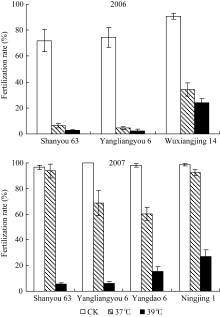
2.1.1 不同高温条件下各品种的受精率 为了消除植株生育时期和颖花发育阶段不同对高温耐性的影响, 必须严格选择生育进程一致的单茎作为高温胁迫效应的调查对象。本试验以高温处理开始后第2天始花的生长较一致的分蘖作为统计对象, 高温处理持续4 d, 以成熟期标记单茎的固定粒位的籽粒受精率作为检测各品种对抽穗开花期高温处理反应的指标。从图2可见, 不同品种对开花期高温胁迫的反应显著不同。在第一年试验中(2006年), 粳型品种武香粳14的高温耐性显著高于2个杂交稻组合汕优63和扬两优6号。在37℃和39℃的2个高温处理条件下, 汕优63和扬两优6号的受精率均已降至接近于0, 而武香粳14尚能维持30%的受精率, 39℃的高温胁迫效应强于37℃, 两温度处理间表现出近10%的差异。第2年的结果(2007年), 所有供试材料在39℃的高温处理下的受精率, 远远低于37℃的高温处理, 显示39℃的胁迫效应远大于37℃的高温胁迫。在39℃高温胁迫下受精率的表现趋势与前一年度相似, 粳型品种宁粳1号显著高于其他材料, 而汕优63和扬两优6号接近于0, 籼型材料扬稻6号居中。在37℃处理条件下, 各供试材料的受精率显著高于上一年度相同处理, 宁粳1号和汕优63表现出较好的耐性, 受精率接近于正常水平。扬稻6号和扬两优6号的受精率显著下降。
综合两年试验结果, 可见供试的粳型品种表现出较好的开花期高温耐性, 杂交稻特别是两系杂交稻开花期高温耐性相对较弱, 开花期39℃高温胁迫下, 受精率几乎接近于0。37℃高温胁迫在两年间结果差异较大, 表明在相对较轻的高温胁迫条件下, 即使是相同品种在不同年际间也有较大差异, 这也表明水稻的生长状况可能对开花期高温耐性有很大影响。汕优63的高温耐性表现较为特殊, 在较轻高温胁迫下, 耐性较好, 但超过一定范围后, 高温耐性迅速下降, 甚至低于一般品种的水平。
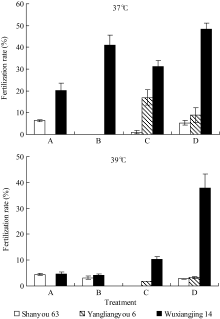
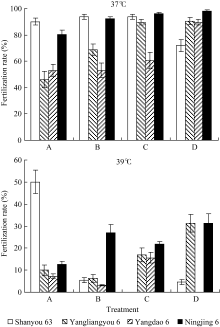
2.1.2 抽穗期高温胁迫时期对受精率的影响 如图3和图4所示, 在颖花开花前后的不同发育阶段, 虽然受高温胁迫时期的差别只有3 d, 最终受精率却有明显的差异, 且其反应在品种间有很大不同。这种反应特征在2007年试验中更为明显。2006年的结果中, 因高温胁迫效应较强, 汕优63的受精率表现很低水平, 处理间差异不明显, 但在武香粳14、扬两优6号中有所表现, 在4 d高温处理期间, 迟抽穗开花者呈受精率较高的趋势。在2007年的结果中, 扬两优6号、扬稻6号和宁粳1号均可见在4 d高温处理期间, 随开花日期推迟受精率上升的明显趋势, 但汕优63则相反, 随开花日期推迟而受精率下降。这种趋势无论是在37℃条件下还是在39℃条件下均可见一致表现。由此表明, 供试品种中颖花开花前后高温胁迫反应的敏感性, 汕优63开花前的花粉充实过程高于开花后的受精过程, 而其他3个品种则表现为开花后的受精过程高于开花前的花粉充实过程。
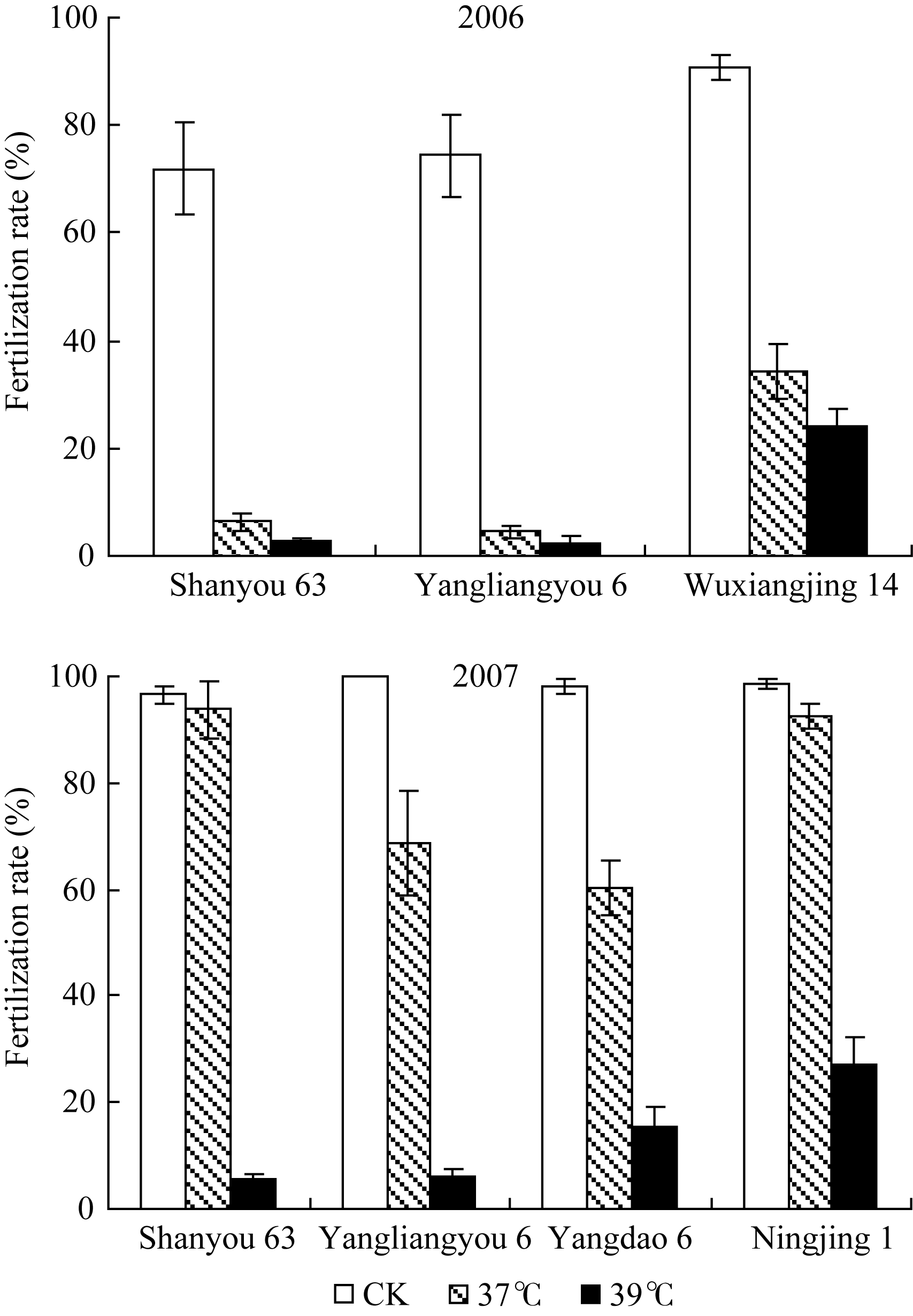 | 图2 高温胁迫对各品种受精率的影响Fig. 2 Effect of high temperature stress on fertilization rate |
 | 图3 开花前后高温胁迫时段对受精率的影响(2006年)Fig. 3 Effect of stress stage of high temperature around the flowering stage on fertilization rate in 2006 |
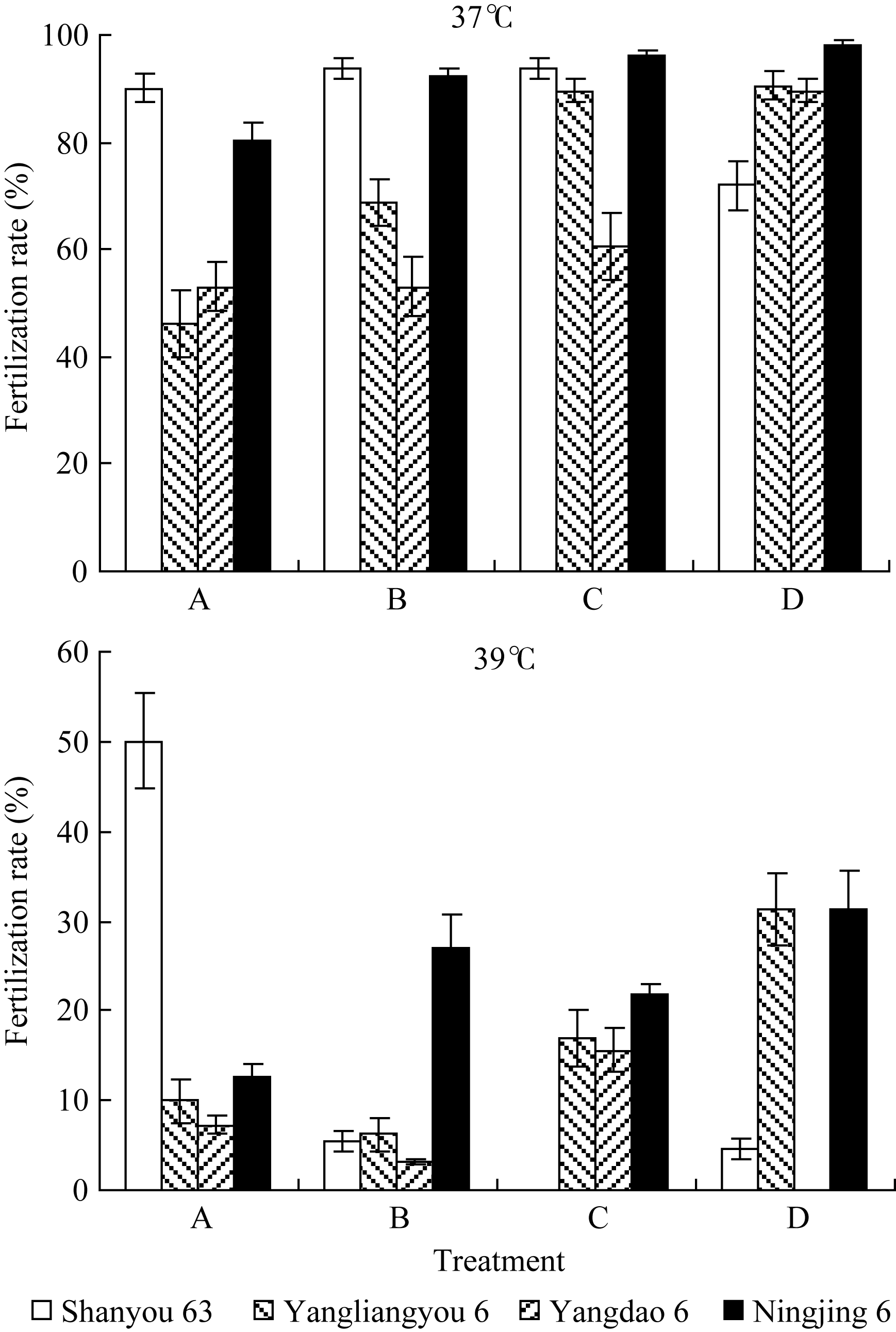 | 图4 开花期高温胁迫时段对受精率的影响(2007年)A、B、C和D为颖花开花前后不同阶段高温胁迫处理, 具体定义如图1。Fig. 4 Effect of stress stage of high temperature around the flowering stage on fertilization rate in 2007A, B, C, and D is the treatment of high temperature stresses around the flowering stage based on different days of heading, showed in Fig. 1. |
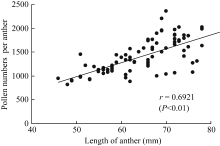
2.2.1 花药形态及其单药花粉数 两年共观测7个品种的颖花形态、花药形态及其单药花粉数状况(表1)。供试品种有籼有粳, 各品种颖花长宽差异明显, 颖长最长的扬稻6号和最短的武香粳14相差2.7 mm, 差幅达27.4%。颖宽最大的宁粳1号比扬稻6号宽1.14 mm, 差幅达28.7%。花药形态也存在类似的差异, 药长最长的为扬稻6号, 达2.44 mm, 最短的武香粳14为1.72 mm, 两者差幅达29.5%, 药宽(花药宽度)最大者为扬稻6号, 最小者为武香粳14, 差幅为31.9%。各品种的单药花粉数差异更大, 最大差异达44.6%, 单药花粉数最多的品种为两优培九, 达每花药1768.5粒, 且供试的2个超级两系杂交稻表现一致, 花粉生产力均表现很高的水平。最少的品种宁粳1号为每花药979.8粒。总体趋势为杂交稻多于常规稻, 籼型品种多于粳型品种。测定结果还表明, 在供试品种中, 花药长度与充实花粉数之间存在极显著的正相关, 花药越长, 充实花粉越多(图5)。
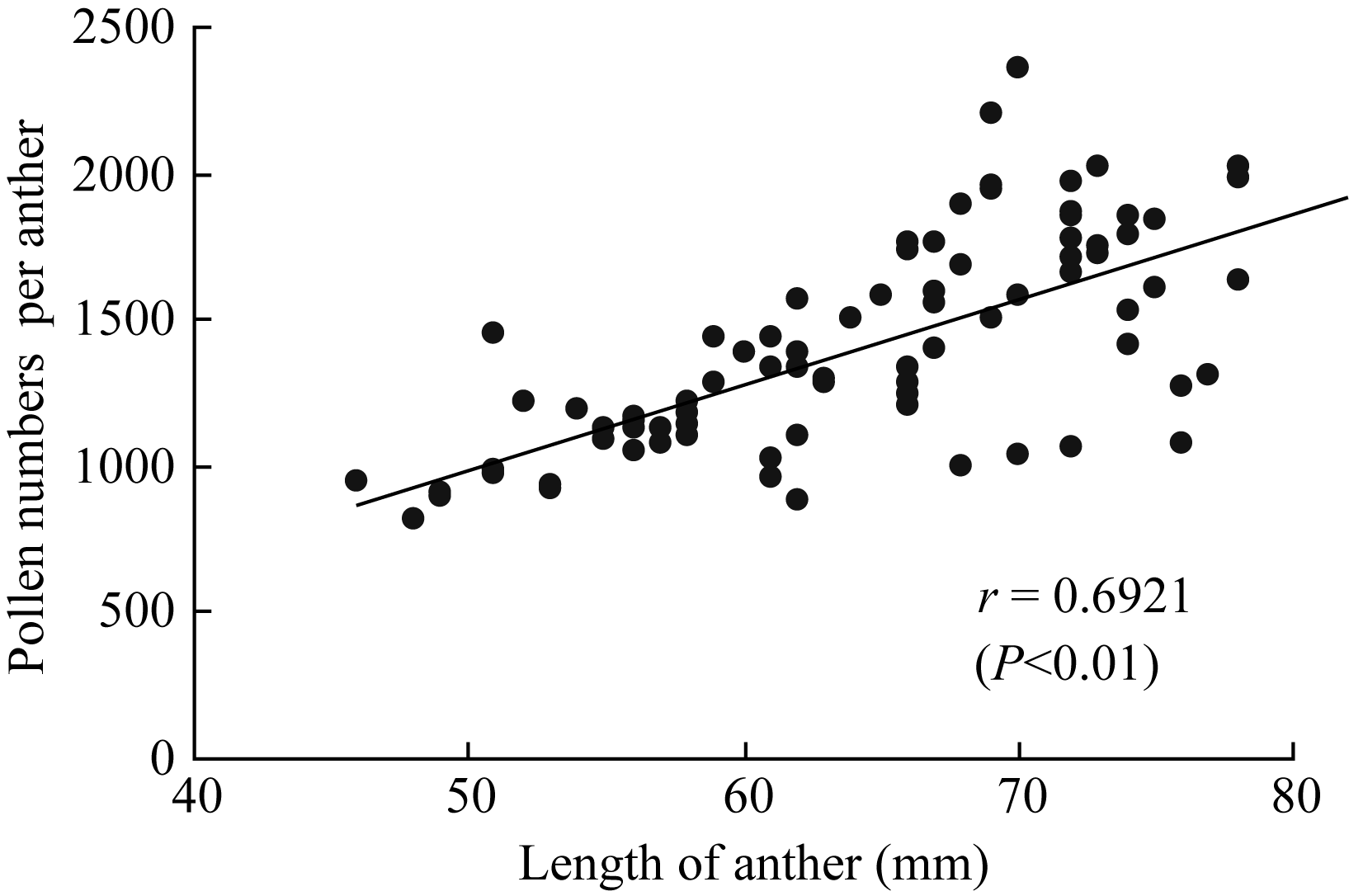 | 图5 充实花粉数与药长的关系Fig. 5 Relationship of filled pollens number and length of anther |
| 表1 供试品种的雄蕊形态和充实花粉数 Table 1 Morphology of stamen and filled pollens number in tested cultivars |
2.2.2 花药开裂率 花药开裂的情况可大致分为3种, 花药两端向中间均开裂为纵裂(完全开裂), 花药只有尖端开裂为孔裂(部分开裂), 两端均未开裂为未开裂, 据此定义, 花药开裂系数 K = (完全开裂花药数+部分开裂花药数)/镜检花药总数。 K值的生物学意义表示花药散落花粉的效率, K值大, 花药开裂能力强, 散落花粉的效率高, 柱头比较容易捕捉到花粉, K值小, 花药散落花粉的效率则低, 柱头上不易着粉。
因为高温处理时间的关系, 高温处理当天抽穗者没有取样, 故表2所列为高温处理后1~3 d的结果, 即相当于图1所示的B、C、D处理的样本结果。可见高温胁迫导致花药开裂系数明显下降。在常规温度条件下, 供试各品种花药开裂系数均在85%以上。在37℃高温胁迫条件下, 供试各品种均有较大幅度的下降, 39℃高温处理下降更为显著。且随着高温处理时间的延长, 花药开裂系数下降幅度明显增加, 这种趋势在39℃高温胁迫条件下更为显著。虽然总的花药开裂率没有显示明显的品种间差异, 但衡量花药完全开裂比例的纵裂率却在品种间有明显的不同, 2个杂交稻组合在处理1 d的条件下下降较少, 但在2 d和3 d的高温处理下大幅下降, 而孔裂率相对下降较少。表明在较强的高温胁迫条件下, 杂交稻的花药开裂可能更容易受到局部的抑制, 而以孔裂为主维持一定程度的散粉。而宁粳1号在较长时间的高温胁迫条件下还能维持一定程度的纵裂率, 保持较多花药能够较好地开裂和散粉。
2.2.3 高温胁迫对柱头捕获花粉量的影响 图6表明, 高温胁迫条件下水稻柱头上的花粉粒数量明显低于适温对照, 且随高温处理强度的增加明显减少, 39℃高温下柱头上的花粉粒多的仅见数粒, 少的甚至未能接受到一粒花粉, 能够观察到花粉的数量也极为稀少。品种间比较, 常温下可见扬两优6号、宁粳1号柱头上花粉数相对较多, 汕优63则相对较少。在37℃的高温胁迫下, 宁粳1号和汕优63柱头上花粉数量相对较多, 这与两者在37℃处理条件下仍可维持较高的受精率表现一致。
| 表2 高温胁迫对花药开裂的影响(2007年) Table 2 Effect of high temperature stress on anther dehiscence in 2007 |
 | 图6 不同温度条件下柱头捕获的花粉量A: 汕优63; B: 扬两优6号; C: 扬稻6号; D: 宁粳1号。Fig. 6 Pollen amount on stigma under different temperaturesA: Shanyou 63; B: Yangliangyou 6; C: Yangdao 6; D: Ningjing 1. |
从图7可见, 单药的充实花粉数与高温胁迫下的受精率并不存在明显的相关。供试的杂交稻组合, 特别是两系杂交稻, 显示出优越的花粉生产能力, 但高温条件下受精率的下降更为明显, 宁粳1号花粉最少, 但高温胁迫下受精率却最好。显示水稻开花期的高温耐性与其花粉生产能力并不存在因果关系。
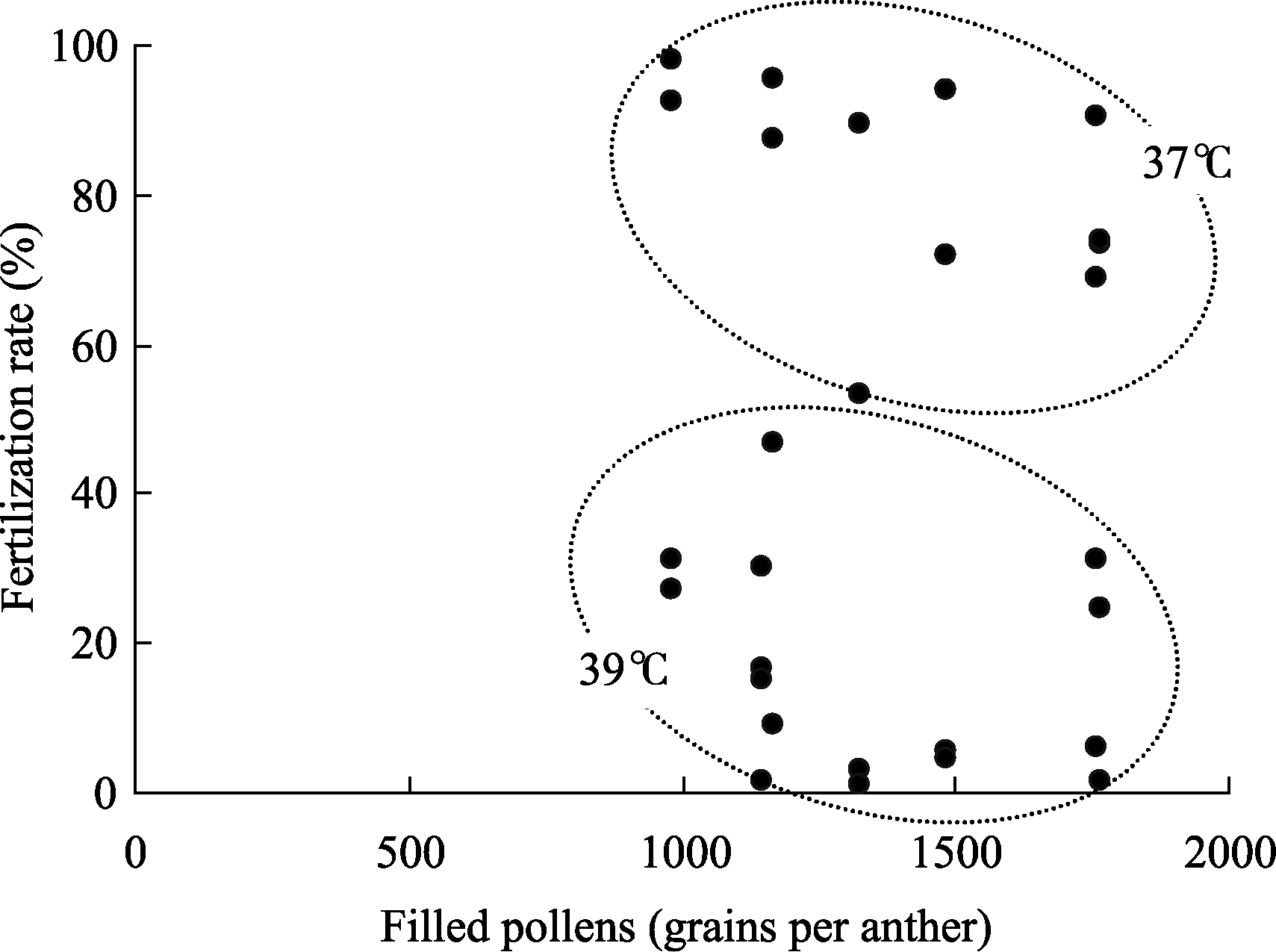 | 图7 供试品种充实花粉数与高温胁迫下受精率的关系Fig. 7 Relationship of filled pollens number and fertilization rate under high temperature |
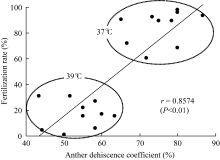
图8表明, 裂药特征对水稻开花期的高温胁迫耐性有较大的影响, 花药开裂系数越大, 高温条件下受精率越高, 高温耐性越强。这种相关性更大程度表现在不同的高温胁迫温度条件上, 37℃条件下花药开裂系数大, 受精率较高, 39℃条件下花药开裂系数低, 受精率低。但分析同一温度条件下各品种间的情况, 则不能看到这种显著的相关, 表明在相同强度的高温胁迫条件下, 影响受精率的决定因素, 除了花药开裂状况之外, 可能还有其他重要因子。
 | 图8 供试品种高温胁迫下花药开裂系数与受精率的关系Fig. 8 Relationship of anther dehiscence coefficient and fertilization rate under high temperature stress |
水稻抽穗开花期热害直接导致籽粒受精障碍而形成空粒, 容易产生严重的产量损失, 这在实际生产中并不少见。抽穗期耐热性较高的品种, 安全生产系数相对较高。本试验结果显示, 在供试品种和组合中, 粳稻品种对抽穗开花期高温表现出较好的耐性, 而两系法杂交稻表现相对较差。两系法杂交稻结实率的生态稳定性较差, 这在以往的研究和实际生产中已有不少直接的观察结果[ 35, 36], 本试验结果也与此基本一致。但供试的粳稻品种抽穗期高温耐性表现较好, 这和以往概念似乎有所不同。在长期的品种分化中, 水稻从低纬度向高纬度地区扩散, 逐渐产生了较为抗寒的粳稻亚种。相对于基本型的籼亚种, 粳稻的一般概念是抗寒不抗热。而且在粳稻生产的较南方地域如江苏南部等地区也常有粳稻热害的现象。然而在本试验中, 武香粳14和宁粳1号高温耐性的表现超过杂交稻, 甚至超过常规籼稻扬稻6号。这和本试验供试品种的选育地点可能有一定关系。这2个品种均在江苏南部选育成功, 在选育过程中, 选育地较高的夏季温度环境, 可能无意中形成品种抽穗开花期高温耐性的选择压力, 因而选择成功的品种也就自然形成对高温的较好的耐性。由此表明, 品种高温耐性的形成或品种特性, 和籼粳品种的生态分化并不一致, 可能并不具备很系统的类型特征, 而是与品种选育的地点关系较大, 从而形成独特的品种个体特征。关于粳稻品种高温耐性的类型特征的描述还有待对更多品种的深入分析。
两系杂交稻扬两优6号在较低的高温胁迫条件下即表现出较弱的高温耐性, 受精率与其他材料相比降到很低的水平, 显示这一在生产上表现优异的超级杂交稻对抽穗开花期的高温天气存在一定的风险, 这在生产中应引起一定的重视, 适当调节播栽期, 使其抽穗开花期避开当地的特高温季节, 可能是保证其稳产高产应该注意的措施。但仅从本试验结果还不能认为两系杂交稻高温耐性较差, 还有必要扩大相应组合材料, 才能得到对两系杂交稻高温耐性的系统评价。另一方面, 适应性很广的籼型三系杂交稻汕优63在较弱的开花期高温胁迫条件下表现较强的耐性, 但在较强的高温胁迫下其耐性迅速下降, 也表现出较为独特的反应特征, 这也从另一层面说明水稻品种的开花期高温耐性, 具有较为复杂的特点, 有时可能要有多个级别的温度胁迫反应数据方能科学评价其强弱。
在对水稻冷害的研究中已经充分证明, 花粉生产能力和水稻孕穗期耐冷性有着十分密切的关系。日本的水稻冷害专家西山岩男研究表明, 柱头上能够捕获一定数量的花粉, 是保证较高受精率的前提条件, 日本型粳稻要达到95%以上的受精率, 柱头上的花粉数量大约要在20粒以上[ 32], 并认为花粉数的决定机制应该是冷害研究的中心路线[ 33], 佐竹等也认为花粉生产能力可以作为水稻耐冷能力的重要指标[ 34]。水稻开花期高温障碍虽然与水稻孕穗期冷害有着本质的区别, 但在颖花受精障碍这一点上, 两者又是相通的。只有在柱头上能够得到一定数量的花粉, 才能有足够的机率保证受精过程的完成。本试验结果表明, 高温胁迫条件下, 开花后柱头上花粉数显著减少, 与受精率的关系也很明显。那么, 按照推测, 花药花粉数多, 同等条件下散落在柱头上的花粉粒也就可能较多, 这似乎与水稻冷害中的现象应该一致。然而观测结果显示完全相反的趋势。开花期高温耐性表现最好的宁粳1号, 其单药花粉数反而最少, 而两系杂交稻扬两优6号的单药花粉数几乎是宁粳1号的两倍, 开花期高温耐性显示最差。这一现象表明单药花粉数和开花期高温耐性没有关系, 似乎与花药本身的开裂机构有关, 开花期的高温有可能直接对花药药壁的组织结构产生物理影响, 使花药的开裂不畅[ 36]。那么, 是什么条件决定了开花期高温胁迫条件下花药开裂的能力?看来有必要从花药本身结构, 例如药壁结构, 花药的结构特征等方面解析, 在今后的研究中这方面的工作值得重视。另一方面, 有研究显示, 花粉活力与高温耐性也存在一定的关系[ 5, 24, 37], 表明综合评价品种抽穗开花期的高温耐性, 还需要从花粉量和质的两方面进行, 相关研究还有待充实。
本试验通过在高温处理开始后标记不同开花日期的方法, 获得了高温胁迫合计4 d, 但受高温胁迫时期有少许差异的稻穗的受精情况的观察结果, 具体而言, 高温胁迫实施在开花前0 d至开花后3 d、开花前1 d至开花后2 d、开花前2 d至开花后1 d和开花前3 d至开花后0 d的稻穗, 形成高温胁迫处理时间在开花日前后有所差异的不同稻穗, 从而能够评价开花前3 d到开花后3 d稻穗的高温耐性。据此分析可见两种类型的表现, 一种为开花前的花粉充实过程对高温胁迫反应敏感程度高于开花后的受精过程, 如汕优63; 一种则为开花后的受精过程对高温胁迫的反应敏感程度高于开花前的花粉完成过程, 如扬两优6号、扬稻6号和宁粳1号, 这种类型的品种, 在开花期和开花后的短时间内受到高温胁迫, 发生大幅度减产的可能性最大。当然, 评价开花前、开花中、开花后的高温耐性, 也就是评价花粉完成期、散粉、花药萌发、受精等不同生理过程对高温胁迫的耐性, 还需要更为细致的实验验证, 但从本试验结果可以看出, 在这一系列过程中, 各阶段对高温胁迫的耐性存在明显的品种特征和品种间差异。为何形成这种对高温耐性的不同, 目前还很难给出合理的推测, 有待更深层次的试验和观察。
供试粳型品种表现出较好的抽穗期高温耐性; 杂交稻特别是两系杂交稻抽穗期高温耐性相对较弱; 汕优63在较轻高温胁迫下表现较好耐性, 但在较强的高温胁迫下高温耐性迅速下降。水稻抽穗期高温耐性与单药花粉数没有相关关系, 与花药开裂和柱头捕获的花粉量关系较大。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|
| 32 |
|
| 33 |
|
| 34 |
|
| 35 |
|
| 36 |
|
| 37 |
|