
{kind=link}
{kind=link}
{kind=link}
北方水稻生育后期剑叶可溶性物质含量及植株生产力对CO2浓度增高的响应
[王惠贞1  , 赵洪亮
, 赵洪亮1 , 冯永祥2 , 姜乐1 , 宁大可1 , 谢立勇1, * , 林而达3, * ]
, 赵洪亮, 林而达]
|
|


以高产优质粳稻松粳9号和稻花香2号为试材, 利用中国北方FACE (free air CO2 enrichment)实验平台研究北方水稻生理代谢对CO2浓度增高的响应规律。在水稻抽穗期开始测定剑叶可溶性糖含量、蛋白质含量和总叶绿素含量, 收获后实测小区产量及产量构成因素, 比较处理间和品种间差异。结果表明, CO2浓度增高显著提高水稻抽穗期、乳熟期和完熟期剑叶可溶性糖含量, 松粳9号和稻花香2号最高增幅分别达11.7%和47.5%。CO2浓度增高显著降低抽穗期和完熟期剑叶可溶性蛋白含量, 松粳9号和稻花香2号最大降幅分别为16.2%和10.5%。CO2浓度增高使抽穗期和乳熟期剑叶总叶绿素含量显著增加, 松粳9号和稻花香2号最大增幅达18.9%和22.5%, 之后便逐渐降低。CO2浓度增高使松粳9号单株籽粒产量、生物学产量、经济系数分别提高6.82%、1.50%和12.64%, 稻花香2号平均升高2.56%、2.13%和26.05%。研究表明, CO2浓度增高最终提高了水稻植株生产力, 对可溶性物质含量的影响在不同生育期存在差异。这可能由于CO2浓度增高促进水稻生长发育, 导致水稻提早成熟, 叶片衰老促进了可溶性糖分解, 也加快了功能叶可溶性蛋白向籽粒运输速率。
, ZHAO Hong-Liang, LIN Er-Da
第一作者联系方式: E-mail:whz0301@163.com; Tel: 18311485048
The responses of high yield and good quality
工业革命以来, 大气中二氧化碳(CO2)含量明显增高。根据世界气象组织(WMO)的年度报告, 至2012年底全球大气CO2浓度已经增加到400.9 µmol mol-1, 比工业革命前的280 µmol mol-1增加了约42.86%。预计到21世纪中期将达450~550 µmol mol-1, 21世纪末将升至730~1020 µmol mol-1 [ 1, 2, 3]。作为作物光合作用的主要底物, CO2浓度增高必然对作物生理代谢与产量品质产生深刻影响[ 4, 5]。水稻属于C3作物, 对CO2浓度增加反应更为敏感[ 6]。水稻又是我国及东南亚等人口大国的主要粮食作物, 所以中国、日本以及菲律宾等东南亚国家都把水稻对高CO2浓度的响应研究作为气候变化对农业生产影响的研究重点。如南京土壤研究所利用江都(32°35′N, 119°42′E) FACE实验平台在亚热带对小麦-水稻轮作开展研究[ 7, 8, 9, 10, 11, 12], 日本东北农业试验站利用岩手(39°38´N, 140°57´E) FACE实验平台在温带开展了水稻生态系统研究[ 13, 14, 15]。研究表明CO2浓度增高促进水稻生育进程, 多数品种表现增产, 但增幅差异较大, 区域间(纬度间)的差异也广泛存在。北方水稻特别是东北粳稻历来以高产优质且口感佳著称, 近年来国内外需求量大, 在发展高产优质高效农业中具有特殊意义, 对保证居民的粮食供应和出口创汇也具有重要意义。本研究对北方水稻剑叶生育后期可溶性物质含量及植株生产力对CO2浓度增高的响应趋势开展试验研究, 探索其响应规律, 为全面揭示高CO2浓度对北方水稻产量生理影响规律及比较与南方水稻反应差异提供理论依据。
FACE实验平台位于北京市昌平南部中国农业科学院昌平实验基地(40°08´N, 116°08´E), 京昌公路的西侧。土壤类型属褐潮土, 耕作方式为冬小麦-水稻轮作, 土壤含有机质18.27 g kg-1、全氮 10.38 g kg-1、全磷1.14 g kg-1、全钾14.17 g kg-1、碱解氮为94.83 mg kg-1、速效磷49.1 mg kg-1、速效钾104.28 mg kg-1, pH 8.28。
选用粳稻品种松粳9号(典型特征为高产稳产, 平均产量10.5 t hm-2)和稻花香2号(典型特征为优质, 全部指标达到农业部优质米一级标准)。松粳9号从出苗到成熟需活动积温2650℃, 株高95~100 cm, 穗长20 cm左右; 稻花香2号需≥10℃活动积温2800℃左右, 株高100~105 cm左右, 穗长21.6 cm左右。
中国北方FACE实验平台的系统构成、系统控制等见郝兴宇等[ 16]报道。该试验设2个处理, CO2为主处理, 为试验地实测大气CO2浓度(389±40 μmol mol-1)和FACE圈增高浓度(600±60 μmol mol-1), 误差为10%, 各设3次重复(即3个FACE圈, 3个对照圈); 副处理为覆膜, 与对照相比, 土壤温度约提高0.5~1.0℃, 含水量约提高3%~6%。采用盆栽(盆长 55 cm, 宽40 cm, 深30 cm), 每圈种植60盆, 两品种各30盆, 每盆栽6穴, 每穴精选苗4株, 每品种半数盆覆地膜。
2012年5月16日育苗, 6月15日移栽, 同时开始对FACE圈内通CO2气体并覆膜处理至收获。对各盆施尿素4 g、磷酸二铵4 g、氯化钾2 g, 7月6日追加氯化钾3 g。FACE圈与对照圈管理方式相同; 覆膜与对照水分管理一致, 移栽前盆土浇透水, 水分管理与大田相同。
参照国家水稻研究中心水稻生育期的划分标准, 结合试验实际情况, 植株8月9日开始进入孕穗期(幼穗在叶鞘内伸长至叶鞘膨胀), 8月16日进入抽穗期(穗顶端从叶鞘中露出, 多数穗开花), 8月30日进入乳熟期(籽粒内容物呈白色乳浆状, 背部仍为绿色), 9月16日进入蜡熟期(籽粒内容物浓黏, 无乳状物出现, 手压穗中部籽粒有坚硬感, 米粒背部绿色逐渐消失, 谷壳微黄), 9月28日为完熟期(谷壳变黄, 不易破碎, 达收获期), 10月2日全部收获。
根据生育进程, 从水稻抽穗期开始共取样5次, 间隔7 d, 取样时间在上午8:00至9:00之间。选取各处理主茎剑叶10片, 分别放于自封袋, 标记后迅速放入冰箱中冷藏, 待测可溶性糖、可溶性蛋白含量及总叶绿素含量。收获期收割测产区植株测植株生产力相关指标。
1.5.1 可溶性糖含量测定方法 采用蒽酮法[ 17]。用牛血清蛋白制作标准曲线。称取剪碎混匀的新鲜叶片0.5 g, 放入盛有25 mL蒸馏水的三角瓶, 沸水浴煮30 min, 取出冷却, 过滤入100 mL容量瓶, 热水冲洗残渣数次, 定容至刻度。提取样品液0.5 mL加等量水, 加蒽酮试剂, 摇匀后用分光光度计于620 nm下比色测得光密度值, 计算可溶性糖含量。
1.5.2 可溶性蛋白含量测定方法 采用考马斯亮蓝G-250法[ 17]。用分析纯葡萄糖制作标准曲线。称取剪碎混匀的新鲜叶片0.5 g, 用5 mL蒸馏水研磨成匀浆, 于3000转 min-1离心10 min, 提取上清液1 mL, 加5 mL考马斯亮蓝G-250溶液, 摇匀后放置2 min, 用分光光度计在595 nm波长下测定吸光度, 计算可溶性蛋白含量。
1.5.3 总叶绿素含量测定方法 采用丙酮乙醇混合液法[ 17](略有改进)。用电子天秤称取0.5 g叶片, 剪碎后放入盛有10 mL的1∶1的乙醇-丙酮混合浸提液的试管中, 室温下黑暗浸提48 h, 待材料完全变白后摇匀, 稀释10倍倒入干净比色皿, 用分光光度法分别测定其在663 nm、645 nm下的光密度值, 计算单位质量叶片总叶绿素含量。
1.5.4 植株生产力测定方法 在成熟期, 从各处理分别取4盆计算和分析植株生产力相关指标的差异。从各处理调查4盆植株的地上部干物质总量即生物学产量; 取代表性植株5株, 105℃杀青30 min,
80℃烘72 h称重, 根据单株平均穗粒数、结实率、千粒重计算单株籽粒产量。取测产区4盆植株, 机械收获称重得到实收经济产量, 经济产量与生物产量的比值即经济系数。
以SPSS统计软件进行数据处理和方差分析, 检验CO2处理和覆膜间的差异显著性, 用Microsoft Excel绘制图表。
2.1.1 可溶性糖含量 可溶性糖是植物碳素营养主要形式, 为作物提供能源物质, 并在维持植物蛋白质稳定方面起重要作用。可溶性糖含量的变化与水稻功能叶片光合作用能力密切相关[ 18]。松粳9号和稻花香2号剑叶可溶性糖含量在生育后期都呈先升高后降低的趋势, 在孕穗期, 二者剑叶可溶性糖含量分别升高13.9%和14.1%, 乳熟期达最大值(表1)。覆膜分别使其平均升高14.0%和18.2%, 增幅差异均达极显著水平, 但FACE条件下覆膜处理却分别降低3.4%和0.4%。
| 表1 FACE及覆膜处理对水稻生育后期剑叶可溶性糖含量的影响 Table 1 Effects of FACE and mulching on soluble sugar content in rice flag leaf during late growth stage |
从抽穗至完熟期整个阶段, FACE条件下松粳9号剑叶可溶性糖含量平均升高11.7%、11.0%、11.3%和11.2%, 而稻花香2号平均升高15.9%、7.3%、3.5%和47.5%, 除蜡熟期, 增幅均达显著或极显著水平。覆膜处理使松粳9号平均增加17.9%、9.1%、4.0%和0.1%, 抽穗期和乳熟期增幅达显著或极显著水平; 稻花香2号平均增加13.0%、5.5%、1.3%和8.3%, 抽穗期和完熟期增幅达极显著水平。松粳9号在孕穗和乳熟期, FACE处理与覆膜处理间分别存在极显著( P<0.01)和显著( P<0.05)交互效应; 稻花香2号在孕穗和抽穗期, 分别存在极显著( P<0.01)和显著( P<0.05)交互效应。
FACE处理对剑叶可溶性糖含量的影响存在品种间差异。在乳熟和蜡熟期, 松粳9号增幅大于稻花香2号, 而孕穗、抽穗和完熟期, 松粳9号增幅小于稻花香2号。覆膜条件下抽穗至蜡熟期, 松粳9号增幅均大于稻花香2号; 完熟期松粳9号增幅小于稻花香2号。
2.1.2 可溶性蛋白含量 可溶性蛋白是植物氮素存在的主要形式, 其含量常作为衡量叶片衰老程度重要指标, 尤其在灌浆期。剑叶可溶性蛋白含量升高有利于维持剑叶生长, 延长光合功能期, 为籽粒碳、氮化合物积累奠定物质基础, 同时反映叶片生
理生化作用状态和叶片氮向籽粒转运状况[ 19]。松粳9号和稻花香2号剑叶可溶性蛋白含量在生育后期均呈下降趋势(表2)。与对照相比, 松粳9号从孕穗至完熟期剑叶可溶性蛋白含量平均降低5.6%、10.6%、9.8%、9.9%和16.2%, 增幅均达显著或极显著水平; 稻花香2号平均降低7.2%、10.5%、4.4%、9.8%和8.6%, 除乳熟期外增幅均达显著或极显著水平。覆膜处理使松粳9号平均提高7.8%、18.9%、15.3%、36.0%和49.7%, 而稻花香2号平均提高13.6%、18.8%、18.9%、38.7%和16.6%。除蜡熟期外, 松粳9号在FACE与覆膜处理间存在极显著( P<0.01)或显著( P<0.05)交互效应, 除乳熟和蜡熟期外, 稻花香2号在两处理间存在极显著( P<0.01)或显著( P<0.05)交互效应。
FACE处理下两品种剑叶可溶性蛋白含量均降低, 孕穗期松粳9号降幅小于稻花香2号, 在乳熟期和完熟期, 松粳9号降幅大于稻花香2号。覆膜条件下均升高, 孕穗、乳熟和蜡熟期, 松粳9号降幅小于稻花香2号, 完熟期松粳9号增幅大于稻花香2号。
2.1.3 总叶绿素含量 功能叶片叶绿素含量与光合作用密切相关, 结实期功能叶片叶绿素含量的变化反映叶片光合能力的变化和叶片衰老进程[ 20, 21], 生育后期保持剑叶较高的叶绿素含量为叶片吸收更多光能提供了良好生理条件[ 22], 为提高产量提供有力保障。松粳9号和稻花香2号剑叶总叶绿素含量在生育后期均呈降低的趋势(表3)。孕穗至乳熟期, FACE处理使松粳9号剑叶总叶绿素含量平均升高24.9%、18.9%和17.7%, 增幅均达显著或极显著水平; 稻花香2号平均升高21.8%、22.5%和20.8%, 除孕穗期外增幅均达极显著水平。覆膜使松粳9号平均降低8.1%、12.4%、15.8%, 除孕穗期增幅均达显著水平; 使稻花香2号平均降低16.1%、10.7%和14.4%, 除孕穗期外增幅均达显著或极显著水平。蜡熟期FACE处理使松粳9号和稻花香2号剑叶总叶绿素含量降低6.0%和12.0%。覆膜使二者降低5.6%和5.8%, 但FACE条件下覆膜使二者剑叶总叶绿素含量升高11.2%和25.0%。松粳9号在FACE与覆膜处理间不存在显著交互效应; 稻花香2号在抽穗期两处理间存在显著( P<0.05)交互效应, 乳熟期两处理间存在极显著( P<0.01)交互效应。
| 表2 FACE及覆膜处理对水稻生育后期剑叶可溶性蛋白含量的影响 Table 2 Effects of FACE and mulching on soluble protein content in rice flag leaf during late growth stage |
孕穗至乳熟期, FACE处理使两品种剑叶的总叶绿素含量都明显提高, 但孕穗期松粳9号增幅大于稻花香2号, 抽穗至乳熟期松粳9号增幅小于稻花香2号。蜡熟期FACE处理松粳9号降幅小于稻花香2号, 覆膜却使松粳9号增幅小于稻花香2号。覆膜降低两者总叶绿素含量, 品种间差异不显著; 但FACE条件下覆膜却使松粳9号增幅小于稻花香2号。
| 表3 FACE及覆膜处理对水稻生育后期剑叶总叶绿素含量的影响 Table 3 Effects of FACE and mulching on chlorophyll content in rice flag leaf during late growth stage |
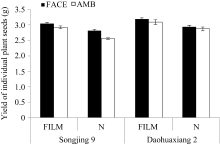
2.2.1 单株籽粒产量 如图1所示, 松粳9号单株籽粒产量低于稻花香2号, 品种间差异达极显著( P<0.01)水平。FACE使松粳9号比对照平均增加6.82%, 达极显著( P<0.01)水平。而稻花香2号比对照平均增加2.56%, 未达显著( P<0.05)水平。覆膜使松粳9号和稻花香2号两品种单株籽粒产量都极显著( P<0.01)提高了, 分别提高11.03%和8.07%。统计分析表明,两品种FACE×FILM之互作效应对水稻单株籽粒产量的影响均未达显著( P<0.05)水平。
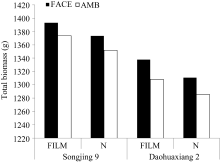
2.2.2 生物学产量 松粳9号生物学产量高于稻花香2号, 品种间差异达极显著( P<0.01)水平(图2)。两品种FACE处理都极显著( P<0.01)高于对照, 松粳9号和稻花香2号分别比对照平均增加1.50%和2.13%。覆膜同样使两品种生物学产量极显著( P<0.01)提高, 松粳9号和稻花香2号分别提高1.51%和1.94%。统计分析表明, 两品种FACE×FILM互作效应对水稻生物学产量的影响均未达显著水平。说明水稻生物学产量品种间差异显著, 并为FACE和FILM处理显著提高, 且增幅表现为稻花香2号大于松粳9号。
 | 图1 FACE及覆膜处理对水稻单株籽粒产量的影响FACE: 开放式空气CO2浓度增加; AMB: 正常大气CO2浓度; FILM: 对土壤做覆地膜处理; N: 土壤正常状况; Songjing 9: 松粳9号; Daohuaxiang 2: 稻花香2号。Fig. 1 Effects of FACE and mulching on seed yield of individual plant in riceFACE: free air CO2 enrichment; AMB: ambient CO2; FILM: plastic-film mulching; N: natural soil. |
 | 图2 FACE及覆膜处理对水稻生物学产量的影响FACE: 开放式空气CO2浓度增加; AMB: 正常大气CO2浓度; FILM: 对土壤做覆地膜处理; N: 土壤正常状况; Songjing 9: 松粳9号; Daohuaxiang 2: 稻花香2号。Fig. 2 Effects of FACE and mulching on total biomass of riceFACE: free air CO2 enrichment; AMB: ambient CO2; FILM: plastic-film mulching; N: natural soil. |
2.2.3 经济系数 松粳9号经济系数高于稻花香2号, 品种间差异极显著( P<0.01)水平(图3)。两品种FACE处理均高于对照, 松粳9号和稻花香2号分别比对照平均增加12.64%和26.05%, 均达极显著( P<0.01)水平。覆膜使松粳9号极显著( P<0.01)降低, 平均降幅为2.54%; 而稻花香2号比对照平均增加12.84%, 达极显著( P<0.01)水平。
统计分析表明, 松粳9号FACE×FILM的互作效应对两品种经济系数影响达极显著水平, 而稻花香2号仅达显著水平(图4)。说明不同品种水稻经济系数对FACE和FILM处理的响应存在很大差异, 两处理对松粳9号影响的差异更明显, 但总体而言, 稻花香2号变幅大于松粳9号。
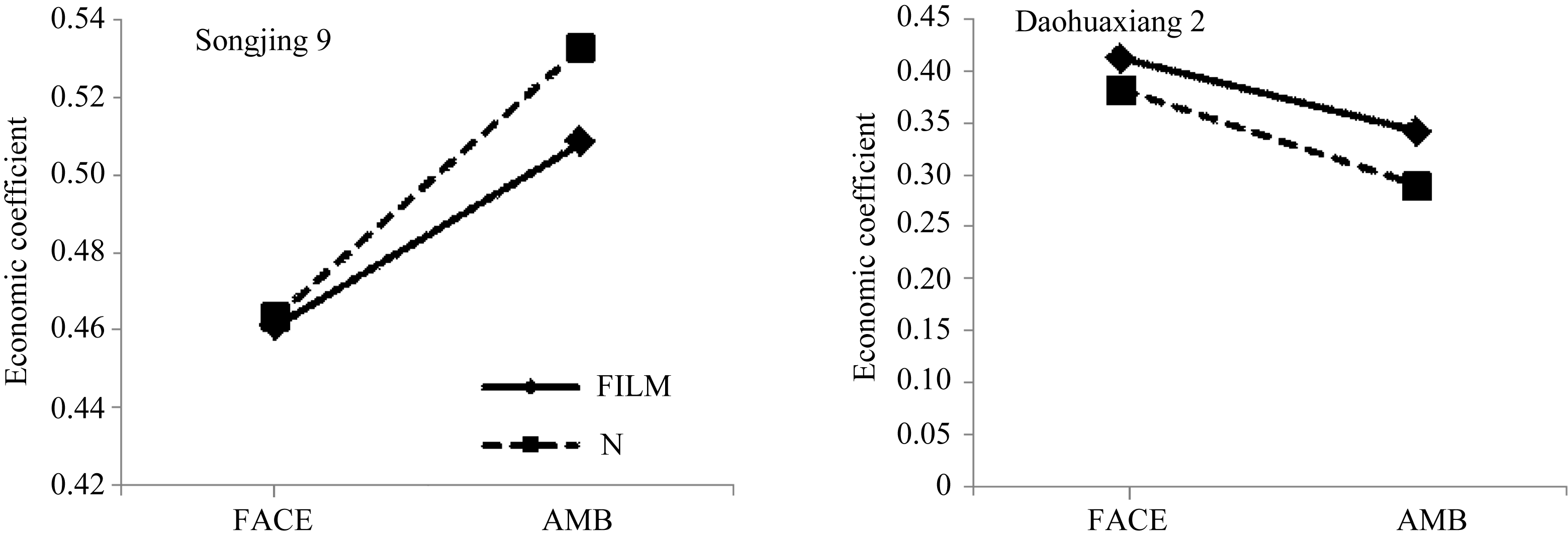 | 图3 FACE及覆膜处理对水稻经济系数的影响 Fig. 3 Effects of FACE and mulching on economic coefficient of riceFACE: 开放式空气CO2浓度增加; AMB: 正常大气CO2浓度; FILM: 对土壤做覆地膜处理; N: 土壤正常状况; Songjing 9: 松粳9号; Daohuaxiang 2: 稻花香2号。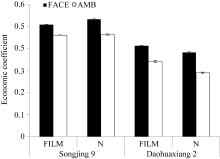 |
一般认为, 从水稻孕穗期开始, 剑叶中可溶性糖开始积累, 并不断向籽粒运输, 乳熟期加速向籽粒输送, 因此生育后期剑叶可溶性糖含量呈先升高后降低的趋势, 乳熟期最高[ 23]。生育后期功能叶片可溶性蛋白是籽粒重要氮源, 籽粒充实阶段剑叶向籽粒源源不断地输氮化合物, 所以剑叶可溶性蛋白含量呈下降趋势[ 24]。土壤水分含量增加使抗旱性品种剑叶可溶性糖含量增加幅度较大, CO2浓度增加使多数品种孕穗期到完熟期剑叶可溶性蛋白含量下降[ 25, 26]。
孕穗期是碳氮代谢旺盛的时期, 可溶性糖是光合作用暗反应主要产物。本研究显示高浓度CO2促进可溶性糖的积累; 而覆膜增加土壤温度, 超过幼穗分化最适温度则不利于糖的形成, 且增加呼吸对糖的消耗。CO2浓度升高和覆膜都可能加快生育进程, 导致功能叶早衰, 增加糖积累量。抽穗至完熟期是决定结实率和粒重的关键时期, 土壤温度及叶片寿命等都可能影响最终产量[ 26]。本研究显示高浓度CO2和覆膜可能导致功能叶早衰, 衰老分解产生部分可溶性糖, 另外覆膜增加土壤含水量, 有利于根吸收水分运输到叶片中, 增强叶片蒸腾, 同时气孔开放增加CO2的摄入, 抑制呼吸对部分糖的分解。蜡熟期对松粳9号影响大于稻花香2号, 原因可能是抗旱性品种剑叶可溶性糖含量水分含量增加时增加幅度较大。
呼吸代谢与氨基酸和蛋白质的生物合成、转化密切相关, 早在19世纪就有人发现呼吸作用随CO2浓度升高而下降, 以后不少研究都证实了这一点[ 27]。本研究显示高浓度CO2和覆膜处理条件下, 水稻生育后期可溶性蛋白含量变化表现出一定规律性, 高浓度CO2使剑叶中可溶性蛋白含量下降, 原因可能是环境中高浓度CO2使叶片早衰, 加快功能叶片合成的蛋白向籽粒中运输, 且CO2浓度升高有加快生育进程的可能, 导致叶片中蛋白提前运输到籽粒中。而覆膜却使两品种整个生育后期剑叶中可溶性蛋白含量都提高, 覆膜增加的土壤水分在一定程度上促进根系吸收更多水分, 延迟叶片衰老, 降低可溶性蛋白质的分解速率。有研究表明, 水分含量较高情况下, 植物体内正常的蛋白质合成受到抑制, 会被诱导出新蛋白, 使蛋白质的含量增加[ 24]。本研究表明孕穗期对松粳9号的影响小于稻花香2号, 完熟期对松粳9号的影响大于稻花香2号, 说明不同时期两品种对高浓度CO2和覆膜的响应程度有差异。
从孕穗期开始, 叶片开始进入功能衰退期, 叶绿素加速分解。本研究显示大气CO2浓度增高使孕穗期至乳熟期剑叶叶绿素含量高于对照, 原因可能是CO2浓度增加条件下, 植物充分利用环境资源, 增加对CO2的同化作用, 需要通过增加叶片叶绿素的含量来提高对光能的捕获能力, 以满足碳同化时能量的需求; 同时CO2浓度升高一定程度上加快生育进程, 减少叶绿素的分解。CO2浓度升高使蜡熟期剑叶总叶绿素含量下降, 可能因为叶片早衰加快叶绿素分解速率, CO2浓度升高条件下覆膜可提高剑叶总叶绿素含量, 抑制早衰。此外, 覆膜使土壤含水量增加, 叶片加速变黄, 影响叶绿素的合成, 与前人研究结果一致[ 28]; 如果覆膜和高浓度CO2同时作用则改善了这种状况, 提高剑叶总叶绿素含量, 有助于植物捕获更多光能供给光合作用。叶片失绿被认为是叶片衰老的最主要和最初症状, 本研究发现存在品种间差异, 松粳9号的抗性大于稻花香2号。蜡熟期至完熟期, 覆膜条件下CO2浓度升高使两品种剑叶总叶绿素含量都升高了, 说明此时期CO2影响更大。
水稻产量是反应植株生产力重要参数, 关于CO2浓度增加对水稻产量影响的报道较多, 但研究条件各异, 多数结果表明, FACE使大部分品种产量极显著提高[ 13]。生物学产量是作物茎叶光合产物贮存累积的结果, 体现该品种的总生产力。日本关于FACE对水稻生物学产量的影响有研究显示平均增幅为12%, FACE使武香粳14的生物学产量平均比对照增产16.2%[ 29, 30, 31], 经济系数比对照减少2.7%[ 16]。有关CO2浓度增加(FACE)对水稻产量及其构成因素影响的研究[ 13, 17, 19]表明多数品种有增产趋势。而本研究显示高浓度CO2使两水稻品种单株籽粒产量和总生物量大幅度提高, FACE处理和覆膜对两品种经济系数的影响存在互作效应, 说明FACE比对照显著增产的重要原因, 经济系数对FACE水稻产量的提高可能有明显影响。
总之, 大气CO2浓度增高促进水稻生长发育, 抽穗后加速可溶性糖及叶绿素的积累, 孕穗开始促进可溶性蛋白向籽粒中运输, 且品种间响应程度在不同生育期有差异; 覆膜延长水稻功能叶片光合功能期, 促进可溶性糖和蛋白积累, 但加快生育后期叶绿素降解。在生育后期, 覆膜可一定程度抑制CO2浓度增高对叶片早衰的影响。CO2浓度增高和覆膜不同程度提高水稻品种植株生产力。由于FACE系统实验地点条件所限, 本试验对北方优质高产水稻的研究并非在北方水稻主产区, 以后的研究中将逐步加以改进。
本研究发现, FACE和覆膜处理均显著提高两个品种抽穗至乳熟期剑叶的可溶性糖含量, FACE处理下松粳9号增幅小于稻花香2号, 覆膜条件下松粳9号增幅大于稻花香2号。FACE使松粳9号剑叶在整个生育后期的可溶性蛋白含量明显降低, 而稻花香2号除乳熟期外均显著增加。覆膜使两品种整个生育后期剑叶可溶性蛋白含量显著升高, 但孕穗期松粳9号变幅小于稻花香2号, 完熟期松粳9号变幅大于稻花香2号。孕穗至乳熟期, FACE使两品种剑叶总叶绿素含量显著升高, 两品种变幅各阶段有差异。FACE和覆膜都不同程度提高水稻单株籽粒产量, 稻花香2号增幅小于松粳9号, 总生物量稻花香2号增幅大于松粳9号; FACE和覆膜对水稻经济系数的影响有差异, 但总体上稻花香2号变幅大于松粳9号。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|
| 19 |
|
| 20 |
|
| 21 |
|
| 22 |
|
| 23 |
|
| 24 |
|
| 25 |
|
| 26 |
|
| 27 |
|
| 28 |
|
| 29 |
|
| 30 |
|
| 31 |
|