{kind=link}
不同生态区冬前低温下白菜型冬油菜不同抗寒品种(系)的比较
[刘自刚1  , 张长生
, 张长生2 , 孙万仓1, * , 杨宁宁1 , 王月1 , 何丽1 , 赵彩霞1 , 武军艳1 , 方彦1 , 曾秀存1 ]
, 张长生, 杨宁宁|
|


以白菜型冬油菜不同抗寒品种(系)为材料, 分别在原种植区(甘肃天水)及北移区(甘肃临洮、兰州和永登)设置田间试验, 采用田间记载、光合参数测定和显微观察相结合的方法, 调查参试品种(系)在4个生态区的苗期形态、光合参数及气孔形态。结果表明, 与原种植区(天水)相比, 冬油菜北移后苗期生长习性由半直立逐渐变为匍匐生长; 冬前低温阶段叶片
, ZHANG Chang-Sheng, YANG Ning-Ning第一作者联系方式: E-mail:Lzgworking@163.com
We conducted field experiments in Tianshui (original planting area), Lintao, Lanzhou, and Yongdeng (extending northern areas), Gansu province, to investigate the seedling morphology, photosynthetic parameters, and stomatal morphological characteristics of
我国油菜生产分为冬油菜和春油菜两大产区, 以山东南部、河北南部、山西南部、陕西关中到甘肃东、南部为界, 向南为冬油菜区, 向北则为春油菜区; 我国北方寒旱区为传统的春油菜种植区[ 1]。在全球气候变化背景下, 我国北方地区最冷月气温明显升高, 低温持续时间明显缩短, 在此情况下, 冬油菜种植区北移成为油菜研究的热点问题, 也是油菜生产应对全球气候变化挑战的重要技术措施之一[ 2]。冬油菜北移至北方寒旱区后越冬期持续低温成为影响油菜生长发育和产量的重要环境因子, 品种的抗寒性是保证冬油菜安全越冬和获得高产稳产性的先决条件。叶片气孔是植物有机体与外界环境进行物质交换的重要通道[ 3, 4], 气孔可改变自身状态敏感地响应低温等逆境[ 5], 通过控制气体交换和水分蒸腾影响光合等生理过程[ 6, 7]。
冬油菜可利用冬前低温阶段有限的光热资源进行光合积累, 冬前充足的有机物积累是越冬期根部抵御冬季严寒而安全越冬, 以及翌年返青进行地上部营养体迅速重建的物质基础。进行冬前低温下北移种植区(多个生态点)与原种植区冬油菜及不同抗寒品种间光合作用及产物分配特征比较, 不同冬油菜品种气孔形态特征变化的分析, 可明确其气孔形态差异及光合作用和产物分配变化规律。孙万仓等[ 1]研究了北方寒旱区冬油菜北移的可行性, 提出我国北方沿长城一线以及西藏、新疆中南部、东北南部存在一个有巨大发展潜力的冬油菜生产带; 并明确了品种抗寒性是冬油菜北移的技术关键[ 1]。此后, 许多研究对白菜型冬油菜品种越冬抗寒的形态特征、生育期进程特点以及生理生化基础等方面进行了分析, 指出低温诱导下白菜型冬油菜抗氧化酶系活性升高, 可溶性糖、游离脯氨酸等渗透调节物积累, 且抗寒性不同的材料在低温下其生理特性变化存在差异[ 8]; 还在白菜型冬油菜中克隆了 BnMKK2[ 9]、 BnMKK4[ 10]、 BnMPK6[ 11]、 BnHMGB2[ 12]等基因cDNA全长, 并分析了基因结构、功能及低温诱导下的表达情况, 结果表明这些基因在响应低温、盐胁迫中发挥重要作用。孙万仓等[ 2]研究表明, 北移区(张掖)冬油菜生长发育特性明显不同于原生产区(天水); 在低温下抗寒性强的冬油菜品种冬前叶片贴地生长、深绿色、生长点低于地表等形态特征明显不同于常温条件, 也与弱抗寒品种间存在较大差异[ 1]。作物通过光合作用形成有机物质, 光合作用也是作物受低温影响最明显的生理过程之一[ 13]。与原种植区相比, 北移区降温早, 冬前生长期中低温阶段相对较长, 白菜型冬油菜北移后, 如何有效利用冬前低温阶段光热资源进行光合有机物合成和积累, 对冬油菜在环境更为严酷的北移区安全越冬和翌年返青后营养体重建具有重要意义。本研究对冬前低温阶段, 北移后不同白菜型冬油菜在不同生态区与原种植区的光合特征变化、北移种植区不同抗寒性品种(系)气孔形态特征差异进行了分析, 以期明确白菜型冬油菜不同抗寒品种(系)光合变化和产物分配规律、抗寒品种(系)低温下气孔形态特征, 为白菜型冬油菜抗寒性选择育种及品种(系)抗寒性鉴定提供理论依据。
于2011—2013年在甘肃省的天水渭南、临洮玉井、兰州安宁、永登秦川4个试点进行试验。天水为冬油菜原种植区试点, 其他3个试点为北方旱寒区冬油菜北移区试点, 其中, 天水试点位于甘肃东南部的黄土梁峁沟壑区, 属秦巴山区西秦岭北部, 大陆性半湿润气候, 油菜区划上属于北方冬春油菜过度区(或北方冬油菜边缘区), 境内四季分明, 光照充足, 在河谷川水区种植甘蓝型冬油菜, 山区种植白菜型冬油菜。北移种植区主要气象因子见表1。8月25日至9月10日依据各试点具体气候特点安排播种, 播前结合翻耕整地施尿素150 kg hm-2、磷酸二铵150 kg hm-2作基肥, 其他管理同大田生产。试验小区6 m2 (2 m×3 m), 重复3次, 随机区组排列。试材为北方寒旱区不同生态区大面积推广应用的主要白菜型冬油菜品种和抗寒性不同的重要育种材料(品系), 品种(系)特性及来源见表2。
| 表1 冬油菜生育期间原种植区与北移区主要气象因子 Table 1 Major climatic and environmental factors in the original and the extending northern regions for winter rapeseed |
| 表2 试验材料抗寒特性及来源 Table 2 Name and source of material tested |
冬前于各试验点观察记载不同白菜型冬油菜的基叶叶型、叶色、叶脉色、叶柄长度、叶尖形状、叶缘形状、叶裂刻、叶柄基色泽、侧裂叶对数、真叶刺毛、苗期生长习性等形态特征。
冬前低温阶段, 于晴天上午10:00至11:30, 在各试验点选取完全展开的新叶片, 在田间条件下采用LI-6400便携式光合仪测定各参试品种(系)叶片净光合速率( Pn)、气孔导度( Gs)、胞间CO2浓度( Ci)、蒸腾速率( Tr)等; 测定时设定叶室(2 cm×3 cm)内温度为20℃, 光强为600 μmol m-2 s-1; 同时采用叶绿素仪测定叶片叶绿素含量、氮素含量、水分含量。每个处理选5株, 每株读数3次。
冬前低温阶段后期(11月5日), 于各试验点田间随机选取各参试品种(系)发育良好健壮株5株, 选完全展开的新叶测定光合参数后整株采挖, 带回实验室冲洗干净, 用吸水纸吸干, 室温晾10 min后测定根长、根粗、侧根数、叶片长度、宽度、叶数。冬前低温阶段指白天温度在0℃以上, 夜间温度在-2℃以下, 日平均温度低于10℃, 土壤表面成昼消夜冻状态的时段。
冬前低温阶段后期(11月5日), 在兰州试点取抗寒性不同的白菜型油菜叶片为材料, 采用水封片法观察叶片表皮及气孔形态。取新展开叶片, 用镊子轻轻撕下叶片表皮, 置于加1-2滴I2-KI染液的载玻片上, 使表皮展开, 盖上盖玻片, 显微观察并照相。
利用SPSS软件统计分析实验数据。
白菜型冬油菜不同品种(系)苗期叶柄较短、黄绿色、叶脉白色、叶片深裂, 在原种植区及3个北移种植区无明显变化。基叶叶型、叶色、叶尖形状、叶缘形状4个性状在天水、临洮、兰州3个生态区无变化, 而北移至永登种植时部分品种(系)性状改变, 如天油2号基叶叶型由裂叶变为完整叶型, 叶缘由锯齿变为波状; 天油5号、天油8号、200119叶色变浅、叶尖性状变尖。白菜型冬油菜从东从南向西向北西扩北移的过程中, 侧裂叶对数总体上呈增加趋势, 苗期基本从半直立向匍匐方向变化, 品种(系)间差异较为明显, 如超强抗寒品种(系)陇油6号、陇油7号在各试点匍匐生长, 而其他品种(系)在天水试点为半直立生长, 北移至永登后变为匍匐生长(表3)。
| 表3 白菜型冬油菜不同品种(系)在不同生态区形态特征差异 Table 3 Morphological comparison of winter rapeseed varieties (lines) in different eco-regions |
冬前低温阶段, 白菜型冬油菜由原种植区(天水)移至北移种植区(兰州和永登)后, 不同品种(系)叶片气孔导度、胞间CO2浓度明显降低, 蒸腾速率显著升高; 而光合速率在品种(系)间存在明显不同, 抗寒性弱的天油系列品种(系)、宁油2号和200119明显下降, 但抗寒性强的陇油系列品种和延油2号明显升高(表4)。在原种植区天油系列品种叶片光合速率明显高于陇油系列品种, 而在北移种植区则低于陇油系列品种, 表现出对来源地光温等气候条件的适应, 而对北移种植区低温等严酷生态环境适应能力较差; 相比而言, 陇油系列品种则具有高效利用北方寒旱区冬前低温阶段光热资源的能力。
原种植区白菜型冬油菜叶片气孔导度大于北移种植区, 而水分蒸腾速率明显降低, 可能与原种植区气候较湿润、大气湿度较高, 降低了气孔下腔与外界环境的蒸气压力差, 水分蒸腾阻力增大有关。
| 表4 不同生态区白菜型冬油菜品种(系)光合参数 Table 4 Photosynthetic parameters of winter rapeseed varieties (lines) in different eco-regions |
白菜型冬油菜北移后, 冬前苗期叶片和根部性状发生了明显变化。与原种植区相比, 在北移种植区的临洮和兰州两试点叶片明显变长, 叶片宽度在兰州和永登两试点有所增加, 而临洮试点变化不显著; 3个北移种植区日均出叶数明显减少, 但不同品种(系)减少幅度有所不同, 陇油系列品种日均出叶数在原种植区与北移种植区间差异达显著水平, 天油5号北移至兰州和永登试点后日均出叶数变化不显著。在冬油菜原种植区, 陇油品种和天油品种日均出叶数差异不大, 而在兰州和永登试点陇油系列品种明显低于天油系列品种。
方差分析结果表明, 同一冬油菜品种(系)的根长、根直径和侧根数在不同试点间存在极显著差异, 冬油菜北移后总体表现根长增长、根直径增加、侧根数变少的基本规律。从表5可看出, 在原种植区不同冬油菜品种(系)的根直径在同一水平, 而在3个北移试点陇油系列品种根直径明显大于天油系列品种。表明在北移种植区陇油系列品种冬前光合有机物被优先分配至地下部, 以增加根粗、根长, 而地上部叶片的生长明显延缓, 根部较多有机物贮存可提高其安全越冬几率, 同时也避免了地上部冬前徒长(表6)。
| 表5 不同生态区白菜型冬油菜营养生长状况 Table 5 Vegetative growth statues of winter rapeseed varieties (lines) in different eco-regions |
冬前低温阶段, 兰州试点白菜型冬油菜不同抗寒品种(系)的气孔密度有显著差异(表6), 其中陇油7号最大为245个 mm-2; 陇油9号最小, 为232个 mm-2, 显著低于其他品种。
| 表6 白菜型冬油菜不同抗寒品种的气孔特征 Table 6 Stomatal characteristics of winter rapeseed varieties (lines) with different cold resistance levels |
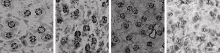
冬前低温阶段油菜不同抗寒品种间保卫细胞叶绿体数目、气孔开张比、气孔开张度差异显著。超强抗寒品种(陇油7号)和强抗寒品种(陇油8号)保卫细胞叶绿体数目在11.8~12.2个之间, 显著高于其他品种。随着品种抗寒性的降低, 气孔开张比、开张度有逐渐下降趋势; 超强抗寒品种叶片气孔有85.6%~87.3%处于完全开放状态, 基本未受驯化低温的影响(表6和图1-A); 抗寒品种陇油8号气孔开张比已降为52.3%, 开张度也明显下降, 其中多数处于半开放状态(表6和图1-B)。
 | 图1 不同品种自然低温驯化后白菜型冬油菜叶片气孔形态比较1: 陇油7号; 2: 陇油8号; 3: 陇油9号; 4: 天油2号。1: Longyou 7; 2: Longyou 8; 3: Longyou 9; 4: Tianyou 2.Fig. 1 Comparison of stomatal morphological characteristics in leaf from different varieties (lines) in cold adaptation |
北方冬油菜一般8月下旬播种, 9月下旬至10月上旬即遇明显降温过程, 到11月中下旬进入枯叶期, 冬油菜冬前生长期大部分时间处于低温环境下, 有效利用冬前低温阶段有限的光热资源积累同化物对冬油菜安全越冬至关重要。冬油菜北移区与原北方冬油菜区是两种完全不同的生态类型, 冬前同期北移区月均平均温度低1.60~7.84℃。与原种植区相比, 白菜型冬油菜北移后冬前低温阶段其气孔导度、胞间CO2浓度下降, 蒸腾速率升高; 弱抗寒品种(天油系列) Pn降低, 而强抗寒品种(陇油系列) Pn升高。原种植区天水试点年降雨量在500~650 mm, 而北移区仅有300 mm左右, 大气湿度也明显不同; 北移区较干燥的气候和较低的大气湿度, 增加了气孔下腔与外界环境的蒸气压力差, 降低了水分蒸腾阻力, 气孔下腔内水分更容易通过气孔进入大气, 引起蒸腾速率的升高。弱抗寒的天油系列品种北移后, 由于 Gs、 Ci、 Pn明显降低, CO2供应不足导致叶片 Pn下降, 气孔限制是北移区天油系列品种冬前低温阶段光合速率下降的原因之一。同一试点天油品种与陇油品种间 Ci无明显差异, 但在原种植区天油系列品种 Pn高于陇油系列, 而在北移区低于陇油系列; 说明北移后冬前低温阶段天油品种光合同化能力弱于强抗寒的陇油品种。在北移区强抗寒的陇油品种Rubisco酶对CO2亲和能力可能高于天油品种, 使RuBP再生能力大为增加, 另外, 强抗寒的陇油品种对北移区低温环境有较强的适应能力, 其光合机构关键组分稳定性较高, 这增加了光合机构同化利用CO2的能力, 使陇油品种 Pn高于天油品种; 同时由于强抗寒的陇油品种在一定范围的低温下气孔仍能保持开放状态, 保证了CO2的供应和补充, 可避免胞间CO2含量因光合消耗而迅速降低。但北移后强抗寒的陇油品种 Pn是否被较低 Ci限制, 即通过增加CO2供给提高 Ci, 能否进一步提升 Pn尚需试验验证。
有关低温对叶片气孔形态的影响, 前人的研究结果有所不同, 烟草等受低温胁迫后气孔关闭[ 14], 而黄瓜幼苗亚低温弱光7 d气孔开放, 低温弱光3 d气孔关闭[ 4]; 在北移区兰州试点, 不同抗寒冬油菜品种气孔形态的观察结果表明, 冬前低温阶段陇油7号多数气孔未受影响而处于完全开放状态, 有利于CO2通过气孔进入胞间, 以增加底物供给从而促进光合的高效进行; 而抗寒性弱的天油品种遇低温后多数气孔逐渐关闭或半关闭, 很容易因气孔限制而引起 Pn下降。这些研究表明叶片气孔形态变化与胁迫强度、处理时间和品种适应特性相关。另外, 冬油菜北移后叶片 Ci降低, 对天油品种而言可能是由部分气孔关闭引起, 而对陇油品种可能是由 Pn升高CO2同化消耗速率增大引起。
许多研究认为耐寒作物如冬黑麦、冬小麦等经历一定时间低温适应后光合能力可恢复至更强的水平[ 15, 16, 17]。抗寒性强的陇油系列品种北移后在同时期冬前较低温度下, 经历一定时间冷驯化适应后使叶片获得高于原种植区的光合能力。白菜型冬油菜是一种耐寒作物, 在低温下细胞冷诱导基因被大量表达, 从而改变着胞内生理状态和物质代谢进程, 这种变化即可能增强叶片光合能力。光合产物的分配和积累对白菜型冬油菜品种抗寒性的形成有重要作用[ 17, 18]。北方寒旱区白菜型冬油菜入冬后地上叶片全
部枯萎, 冬油菜以地下根部越冬, 翌年春季返青后进行地上部营养体的重新构建; 安全越冬和翌年返青进行地上部营养体重建, 都需要根部冬前积累充足的贮藏物。白菜型冬油菜抗寒性强的品种根部直径、长度明显高于弱抗寒品种, 强抗寒品种光合产物较多被分配到地下, 供根部生长并贮存积累, 使其安全越冬以及翌年迅速返青和营养体重建得到保障。根粗是根重的重要构成因子, 在白菜型冬油菜生产实践中也观察到根直径在1.0 cm以上是其在甘肃河西地区安全越冬重要形态特征。
| 1 |
|
| 2 |
|
| 3 |
|
| 4 |
|
| 5 |
|
| 6 |
|
| 7 |
|
| 8 |
|
| 9 |
|
| 10 |
|
| 11 |
|
| 12 |
|
| 13 |
|
| 14 |
|
| 15 |
|
| 16 |
|
| 17 |
|
| 18 |
|