 Minsoy
Minsoy ×
× Essex
Essex and
and{kind=link}
大豆产量及主要农艺性状QTL的上位性互作和环境互作分析
梁慧珍1  , 余永亮
, 余永亮1 , 杨红旗1 , 张海洋1 , 董薇1 , 李彩云1 , 杜华1 , 巩鹏涛2 , 刘学义3 , 方宣钧4
, 余永亮|
|


第一作者联系方式: E-mail:lhzh66666@163.com, Tel: 0371-65751589
以栽培大豆晋豆23为母本, 半野生大豆灰布支黑豆ZDD2315为父本杂交衍生的F2:15和F2:16的447个RIL家系为遗传群体, 绘制SSR遗传图谱, 采用混合线性模型方法, 对2年大豆小区产量及主要农艺性状进行加性QTL、加性×加性上位互作及环境互作分析。结果检测到9个与小区产量、茎粗、有效分枝、主茎节数、株高、结荚高度相关的QTL, 分别位于J_2、I、M连锁群上, 其中小区产量、茎粗、株高、有效分枝和主茎节数QTL的加性效应为正值, 说明增加这些性状的等位基因来源于母本晋豆23。同时, 检测到7对影响小区产量、茎粗、株高和结荚高度的加性×加性上位互作效应及环境互作效应的QTL, 共发现14个与环境存在互作的QTL。上位效应和QE互作效应对大豆小区产量及主要农艺性状的遗传影响较大。大豆分子标记辅助育种中, 既要考虑起主要作用的QTL, 又要注重上位性QTL, 才有利于性状的稳定表达和遗传。
, YU Yong-LiangImproving seed yield is an important goal of soybean breeding programs. In this investigation, a soybean SSR genetic linkage map constructed by a total of 447 recombinant inbred lines (RILs) derived from a cross between Jindou 23 (cultivar, female parent) and ZDD2315 (semi-wild, male parent) and the mixed linear model was used to identify the QTLs of yield and other QTLs for major agronomic traits in a two-year experiment. Nine QTLs bearing additive effects for pod position, plant height, node number on main stem, branch number, stem thickness and yield per plot were mapped in the linkage groups J_2, I, and M. The QTLs of yield per plot, stem thickness, plant height, branch number and node number on main stem showed positive additive effects donated by Jindou 23. Seven pairs of epistatic effects QTLs for pod position, plant height and stem thickness were detected, which had an interaction with environments. The results indicated that the epistatic effects and the environmental factors played an important role in yield per plot and agronomic traits in soybean. It will be very important to pay attention to not only QTLs with major effects but also those with epistatic effects in soybean molecular marker-assisted breeding in considering the stability expression and inheritance of the agronomic traits.
大豆具有较高的营养价值, 广泛应用于工业与食品。长期以来, 大豆产量偏低的问题, 一直困扰着大豆产业发展。大豆产量是受多基因控制的综合数量性状, 容易受遗传、生理、环境、形态以及农艺性状的影响[ 1]。近年来, 分子遗传学和分子标记技术快速发展, 构建出了高密度遗传连锁图谱, 推动了数量性状的QTL基因定位研究进程[ 2]。根据SoyBase大豆数据库(http://soybeanbreederstoolbox.org/)最新公布的数据, 对大豆产量及相关性状的QTL定位已经超过350个, 并且几乎覆盖所有的连锁群。以往对大豆产量和相关农艺性状的研究, 主要集中于QTL的定位及各座位间的互作, 而较少涉及QTL间的上位性和环境互作效应, 这主要是受到了遗传分析模式研究和分析软件研发的限制[ 3, 4, 5, 6, 7, 8]。朱军[ 9]采用混合线性模型, 扩展了以往的广义遗传模型分析方法, 使基因和环境的互作效应无偏分析变成了现实。Wang等[ 10]据此开发出了QTL和QE互作分析软件。研究人员对玉米[ 11, 12]、水稻[ 13, 14]、小麦[ 15, 16]等作物进行了QTL与环境互作分析, 对大豆蛋白质、脂肪和异黄酮[ 17, 18, 19]等已有研究报道, 但对产量特别是小区产量诸性状QTL间的上位性和环境互作效应研究较少。受遗传背景和环境条件的影响, 前人对大豆产量和相关性状定位出的QTL, 在用于分子标记时, 经常出现屏蔽。同时, QTL间的上位性效应对小区产量诸性状表型值的组成具有重要作用。本研究对2011—2012年种植的大豆晋豆23×半野生大豆灰布支黑豆重组自交系进行小区产量及主要农艺性状QTL定位和加性效应、加性×加性上位互作效应及环境互作效应分析, 以期用于指导实践中的分子标记辅助育种工作。
以栽培大豆晋豆23为母本, 半野生大豆灰布支黑豆(ZDD2315)为父本所衍生的447个株系的F2:15和F2:16重组自交系作为性状遗传分析的供试群体。
以随机区组设计, 3次重复, 行长4 m, 行距0.4 m, 株距0.13 m。周边设6行保护行, 消除边行优势。2011和2012年连续2年在河南省农业科学院现代农业科技试验示范基地(河南省原阳县)种植, 一播全苗, 按照大田模式进行田间管理。成熟后, 将4行区全部收获计产。按照《中国大豆品种志》的标准调查小区产量(yield per plot, YLD)、结荚高度(pod position, PP)、株高(plant height, PH)、主茎节数(node number on main stem, NNMS)、分枝数(branch number, BN)、百粒重(100-seed weight, SW)、茎粗(stem thickness, ST) 7个性状。
分别取2011和2012两年测量数据的平均值, 利用DPS软件[ 20]对各性状考察并作相关性分析。
通过对群体亲本两地表型数据统计(表1), 发现2个亲本在7个性状间差异较大, 为QTL分析提供了较好的遗传背景。RIL群体的最小值和最大值之间差异明显, 表型数据的随机误差项服从正态分布, 满足QTL分析要求。
| 表1 RIL群体亲本和群体家系性状描述统计分析 Table 1 Phenotypic variation of all traits in RIL and the parents |
表2表明, 不同环境条件下的相关分析结果基本一致。小区产量与结荚高度、株高、主茎节数、分枝数、百粒重和茎粗均呈现显著或极显著正相关, 茎粗在2012年表现出与株高、主茎节数、分枝数、结荚高度、百粒重极显著正相关, 2011年表现出与株高、主茎节数、分枝数极显著正相关。其他各性状间均显著或极显著正相关。说明小区产量诸性状之间存在着既相互影响又相互制约的关联关系, 在实际育种工作中为表型相关选择提供了参考。
| 表2 7个性状之间的相关性系数 Table 2 Correlation coefficients between seven traits |
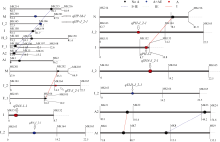
联合分析2年的结荚高度、株高、主茎节数、分枝数、百粒重、茎粗、小区产量性状数据, 共检测到9个QTL (表3和图1)。检测到7个性状的加性效应为-0.6633~5.3910, 对性状贡献率为1.14%~13.01%, 其中, 小区产量、茎粗、株高、有效分枝和主茎节数均表现遗传正效应, 即增加其性状的等位基因来源于母本晋豆23。除百粒重外, 其他性状与环境互作效应贡献率为0.02%~9.74%。所有性状( qPP_M_1和 qYLD-J_2-1除外)与环境互作的贡献率均小于其自身的加性效应贡献率。
| 表3 各个性状的QTL及QTL与环境互作 Table 3 QTL and QE interaction for all traits |
 | 图1 检测到的QTL及加性效应QTL在连锁群上的分布Fig. 1 Location of QTLs and epistatic effects QTLs on linkage groups |
小区产量、茎粗和有效分枝的QTL均位于J_2染色体, 加性效应值分别为0.4894、3.61和7.59, 贡献率分别为3.06%、3.61%和7.59%; 3个性状均与环境之间存在互作, 互作贡献率分别为9.74%、1.62%和4.95%。小区产量的加性贡献率小于互作贡献率, 说明环境对产量的影响较大。
共检测到2个主茎节数QTL, 分布在J_2和N上, 加性效应值分别为1.1541和1.0368, 贡献率分别为13.01%和9.21%; 2个QTL均与环境存在互作效应, 互作贡献率分别为1.43%和1.12%。
共检测到2个株高QTL, 分布在J_2和I上, 加性效应值分别为5.3910和4.5808, 贡献率分别为7.78%和6.64%; 2个QTL均与环境存在互作效应, 互作贡献率分别为0.45%和0.02%, qPH-I-1与环境的互作贡献率较小。共检测到2个结荚高度QTL, 分布在J_2和M上, 加性效应值分别为0.7097和-0.6633, 贡献率分别为2.77%和1.14%; 2个QTL均与环境存在互作效应, 互作贡献率分别为1.58%和1.48%。主茎节数、株高和结荚高度分析结果表明加性贡献率大于互作贡献率, 说明基因比环境对这3个因子的影响显著。
共检测到7对影响小区产量、结荚高度、株高和茎粗的加性×加性上位互作效应的QTL (表4)。2对小区产量QTL上位性效应的效应值分别为-0.1858和-0.1944, 贡献率分别为2.00%和3.27%, 效应值均为负值, 表现出重组型大于亲本型。2对QTL均存在上位互作效应, 互作贡献率分别为2.30%和4.42%。1对茎粗QTL上位性效应的效应值
为-0.0286, 贡献率为3.32, 效应值为负值, 为重组型大于亲本型。这对QTL存在上位互作效应, 互作贡献率为1.38%。3对结荚高度QTL上位性效应的效应值分别为0.7538、0.5989和0.5878, 贡献率分别为1.11%、0.99%和1.51%, 效应值均为正值, 为亲本型大于重组型。3对QTL均存在上位互作效应, 互作贡献率分别为1.16、1.08%和0.82%。1对株高QTL上位性效应的效应值为4.8742, 贡献率为5.45, 效应值为正值, 为亲本型大于重组型。这对QTL存在上位互作效应, 互作贡献率为1.14%。
| 表4 各个性状加性×加性上位互作效应QTL Table 4 Epistatic effects QTL of additive × additive for all traits |
小区产量可以理解为构成性状, 产量是复合形状, 本研究选择“小区产量”作为个体值进行QTL定位, 研究结果可能更为有效。首先, 复合性状比构成性状受到更多的QTL控制, QTL的连锁关系和遗传效应更加复杂。因而, QTL作图功效明显下降, 使用复合性状作图的意义不大[ 25]。其次, 遗传力 h2= VA/ VP, VA和 VP均为个体值的方差。产量是复合性状, 往往是不准确的。即使采用, 也由于竞争的作用而意义不大。第三, 构成性状和复合性状在遗传研究和实际育种中的使用, 应考虑到二者的研究目标不尽相同。遗传研究的目的是探索控制目标性状基因的遗传规律, 相对“产量”(复合性状)而言, “小区产量”(构成性状)遗传模型较为简单, 受较少的QTL控制, 更易于进行单个QTL遗传研究; 而作物育种的目的, 是要尽可能多地选择优良基因和有利基因组合。和多性状选择指数[ 26]类似, “产量”作为复合性状, 在育种工作中可以同时选择相互之间有影响的多个有利等位基因, 提高实际育种工作效率。因此, 选用“小区产量”更适合于QTL作图研究, 在育种中“产量”则更为常用。
本研究检验出的QTL贡献率值在各年份间大小不同, 联合数据分析结果往往出现QTL贡献率值低于不考虑QTL与环境互作的结果。2年间大豆小区产量诸性状的QTL与环境互作, 共检测到9个加性效应QTL, 相对数量较少, 且每个性状均有1个QTL定位在J染色体上, 呈现出很大的集中性, 这与前人的研究结果有所不同。同时, 有1个结荚高度QTL定位在M连锁群上, 各有1个株高和主茎节数QTL定位在I连锁群上, 与已有研究结果类似[ 27], 又呈现出较好的稳定性和真实性。加性效应的9个QTL贡献率总体表现较低, 分布在1.44%~13.01%之间, 其中4个QTL的贡献率低于4%, 只有一个大于10%。为了验证结果的可靠性, 选用WinQTL Cart 2.5复合区间作图法中Zmapqtl方法的Model 6进行了QTL分析。设定QTL的显著性水平为0.05, 当LOD>2.5时, 即认定该位置存在一个QTL。结果共定位出5个结荚高度QTL, 分别位于A2、C2、J_1和J_2上; 7个株高QTL, 分别位于A2、B1、C1、C2、I和M上; 3个茎粗QTL, 分别位于A2、C2和J_2上; 6个小区产量QTL, 分别位于A2、C1、C2、J_2、N和M上; 分别于与 QTLNetwork 2.0 定位结果中的结荚高度J_2、株高I、茎粗J_2和小区产量J_2相吻合; 同时, 用WinQTLCart 2.5定位出2个主茎节数QTL, 分别位于C2和H_1上、3个有效分枝QTL, 分别位于C1、C2和O上、2个粒厚QTL, 分别位于A2和C2上, 与 QTLNetwork 2.0 定位结果不一致(表5)。
| 表5 用WinQTLCart检测出的各个性状QTL位置及遗传效应 Table 5 Putative QTL and their genetic effects for all traits detected by WinQTLCart |
综上所述, 我们认为QTLNetwork 2.0分析结果应该是可靠的。分析结果中定位结果较少、贡献率偏低产生的原因可能是分析软件综合考虑了加性、上位性以及与环境互作效应, 以往的研究中考虑与环境的互作效应较少。
环境对大豆产量有重要影响。同一性状在不同的环境下的QTL常常表现出较大差异[ 28]。遗传力越低, 受环境影响越大; QTL效应越小, 越容易易受环境条件影响[ 29]。品种与环境互作的存在是品种稳定性产生的根源, 对其效应进行准确的估计是合理评估作物品种稳定性的基础[ 30]。本研究共检测到9个有加性效应的QTL, 除 qPH-I-1没有互作外, 其他QTL都与环境存在互作效应, 其中 qYLD-J_2-1与环境互作贡献率明显高于加性效应的贡献率, 说明环境比基因对大豆产量的影响更大。该研究结果从理论上进一步解释了大豆产量对纬度、光照比较敏感和“百里不引豆”的主要原因。
经典数量遗传学认为, 性状相关是由于基因多效性或者基因紧密连锁引起的。本研究检测到控制小区产量、茎粗、有效分枝、主茎节数、株高、结荚高度6个性状有加性效应的9个QTL中, 每个性状均有1个QTL定位在J染色体上, 呈现出很大的集中性。株高、主茎节数、分枝数、结荚高度、百粒重均呈现极显著正相关, 说明控制这些性状的基因存在着紧密连锁。同时, 尽管控制这些性状的QTL存在紧密连锁关系, 但并不排除控制这些性状的基因独立存在。有1个结荚高度QTL定位在M连锁群上, 各有1个株高和主茎节数QTL定位在I连锁群上, 与已有研究结果类似[ 29], 说明它们在遗传上也可能是相互独立的, 具有较好的稳定性。
检测到9个大豆小区产量诸性状的QTL, 全部表现出与环境的互作效应; 同时检测出7对上位互作位点, 说明加性效应、加性×加性上位互作效应及环境互作效应是小区产量、茎粗、有效分枝、主茎节数、株高和结荚高度等性状的重要遗传基础, 在进行分子标记辅助育种时, 既要考虑起主要作用的QTL, 又要关注其上位性QTL, 这样更有利于性状的稳定表达和遗传。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|