



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
核盘菌诱导下甘蓝型油菜防御相关基因表达差异分析
引用本文
马田田, 彭琦, 陈松, 张洁夫. 核盘菌诱导下甘蓝型油菜防御相关基因表达差异分析. 作物学报, 34(3): 416-423
MA Tian-Tian, PENG Qi, CHEN Song, ZHANG Jie-Fu. Differential Expression of Defense Related Genes inBrassica napus Infected bySclerotinia sclerotiorum . 作物学报, 34(3): 416-423 
Permissions
MA Tian-Tian, PENG Qi, CHEN Song, ZHANG Jie-Fu. Differential Expression of Defense Related Genes in
Copyright©2014, 作物学报编辑部
作物学报编辑部
核盘菌诱导下甘蓝型油菜防御相关基因表达差异分析
摘要
菌核病是油菜主要病害, 至今尚未在油菜及相关植物中找到抗性基因。本研究利用qRT-PCR法比较了抗病品种宁RS-1和感病品种APL01在接种核盘菌后0~48 h内11个防御相关基因的表达差异, 以揭示抗病品种宁RS-1的抗病机制。结果表明, 4个基因(
关键词:
核盘菌; 甘蓝型油菜; 防御基因; 表达差异
Differential Expression of Defense Related Genes inBrassica napus Infected bySclerotinia sclerotiorum
Abstract
Sclerotinia stem rot is the main disease of rapeseed. Up to date, genes involved in defending
Keyword:
Sclerotinia sclerotiorum; Brassica napus; Defense gene; Differential expression
核盘菌[ Sclerotinia sclerotiorum(Lib) de Bary]是一种常见的腐生型病原真菌[ 1], 其寄主范围非常广泛, 可感染75个科, 278个属的408个种和42个亚种或品种的植物[ 2]。核盘菌的致病机制比较复杂, 其致病基因编码着侵染寄主所必需的一系列产物。研究表明草酸(oxalic acid, OA)和细胞壁降解酶(cell wall degrade enzymes, CWDEs)是核盘菌致病的重要因子[ 3, 4], 通过提高植物体内草酸氧化酶和细胞壁降解酶抑制蛋白的表达可以有效抑制核盘菌的侵染[ 5, 6]。
目前, 对油菜中与核盘菌侵染过程中植物组织细胞内的分子变化及抗性相关基因的表达情况还知之甚少[ 7], 抗病相关基因的研究还停留在功能基因的筛选与克隆阶段。Mei等[ 8]构建了抗、感菌核病甘蓝F2群体, 在C09染色体上获得2个主效QTL。Wen等[ 9]利用蛋白组双向电泳技术比较了抗、感菌核病甘蓝型油菜的蛋白质表达谱, 获得20个与抗病株系抗性相关的蛋白。刘仁虎等[ 10]以拟南芥cDNA芯片筛选核盘菌侵染时甘蓝型油菜的局部抗(耐)病相关基因, 共获得在病斑周围局部组织中受核盘菌胁迫后表达变化达2倍以上的基因61个。这些基因中, 36个上调表达, 25个下调表达。通过分子标记或者生物芯片技术分析得到的抗病相关基因很多, 但是其中真正有应用价值的很少, 大部分是借鉴其他植物中分离的基因再转化油菜, 研究其在油菜中的抗病作用。因此, 抗病基因尤其是抗菌核病功能基因的分离与应用仍将是近期内油菜抗病研究的重点[ 11]。
江苏省农业科学院经济作物研究所在育成抗菌核病种质宁RS-1的基础上[ 12], 构建了宁RS-1接种核盘菌前后的SSH文库, 获得了多个与植物抗病、防御反应、信号传递等相关的基因[ 13]。为了进一步研究这些基因在核盘菌侵染过程中的表达规律, 揭示宁RS-1在核盘菌侵染过程中所涉及的防卫反应和信号传递机制, 本研究筛选了多个防御相关基因(表1), 分析了抗病与感病油菜品种在接种核盘菌前后不同时间内各基因的表达动态差异, 以期获得与宁RS-1抗菌核病密切相关的基因。
| 表1 基因及其相关研究 Table 1 Gene and related research |
1 材料与方法
1.1 供试材料
抗菌核病品种宁RS-1[ 25, 26]、感病品种APL01[ 27], 均由江苏省农业科学院经济作物研究所提供, 苗期和花期菌核病接种鉴定表明, 这两品种间抗性差异达极显著水平[ 28]。
供试菌株的菌核采自江苏省农业科学院试验田油菜残茬, 经表面消毒, 并适量巴斯消毒液浸泡3~5 min, 转入75%乙醇漂洗数秒后迅速用无菌水洗净。然后接种于马铃薯培养基(PDA)上, 萌发菌丝后
于4℃保存用作菌源, 接种前3 d继代一次备用。
1.2 试验方法
1.2.1 病菌接种和取样 选取终花期的油菜为被接种植株, 接种前将牙签置盛有PDA培养液的烧瓶中, 浸泡24 h, 再经灭菌备用。将菌丝块接种到PDA平板中央, 把牙签呈放射状均匀摆放在菌丝块周围, 每个培养皿20根, 活化菌丝培养2 d后用于接种, 用消毒电钻在离地面50 cm的茎杆处打孔, 插入带菌牙签。每个品种各接种长势一致的3株植株。分别在接种前和接种后12、24和48 h取主轴及上部2个分枝同一部位开花后5 d的幼嫩角果。装入离心管后迅速冷冻于液氮中, -80℃保存。
1.2.2 总RNA提取及cDNA制备 采用Promega公司生产的SV total RNA提取试剂盒(Z3105), 按其说明书提取幼嫩角果样品的RNA, 样品起始量为100 mg。以琼脂糖凝胶电泳检测RNA质量, 紫外分光光度计检测RNA总浓度, 并根据结果用DEPC水调整RNA终浓度至250 ng μL-1。采用TaKaRa公司生产的RT Reagents反转录试剂盒合成cDNA, RT反应液含5 × Prime Script buffer (for Real-time PCR) 2 μL和总RNA 2 μL, 用RNase Free ddH2O补充至总体积10 μL, 将反应后得到的cDNA保存在-20℃冰箱备用。
1.2.3 引物设计 根据在NCBI网站检索得到的各基因序列, 利用Primer Premier 5.0软件设计并筛选, 获得荧光定量PCR扩增所用的特异性引物, 并由华大基因科技有限公司合成引物, 引物序列见表2。
| 表2 荧光定量PCR引物序列 Table 2 Primers used for qRT-PCR |
1.2.4 荧光定量PCR 采用ABI公司的7500型荧光定量PCR仪, 参照TaKaRa公司的Prime Script RT Master Mix试剂盒(DRR036A)介绍的方法进行qRT-PCR。以油菜 Actin基因(AF1l1812)作为内标基因, 采用2-ΔΔCT法分析数据, 荧光染料为SYBR Green I, 分别检测抗病相关基因在油菜接种核盘菌前后的相对表达量的动态变化, 进一步研究菌核病诱导后不同抗性品种中基因的表达差异。实时荧光定量PCR反应体系含SYBR Premix Ex Taq 10 μL、10 μmol L-1上下游引物各0.4 μL、cDNA模板2 μL, 以ddH2O补充体积至20 μL。PCR反应程序为预变性95℃, 30 s; 95℃ 5 s, 60℃ 34 s。根据不同引物调整退火温度, 每个样品设3次技术重复, 试验共设3次生物学重复。
2 结果与分析
2.1 油菜总RNA检测
取接种核盘菌前后不同时间的宁RS-1和APL01样品, 提取总RNA, 经琼脂糖凝胶电泳检测, 样品的总RNA条带清晰(图1), 质量较好; 总RNA紫外分光光度计检测, A260nm/ A280nm比值为2.0, 表明
 | 图1 油菜角果总RNA琼脂糖凝胶电泳图Fig. 1 Electrophoretogram of total RNA from silique of Brassica napus L. |
总RNA纯度较高, 可用于荧光定量PCR分析。
2.2 油菜接种核盘菌后抗氧化相关酶基因的表达差异
Cu/ZnSOD基因的表达量在抗、感品种内均呈先降后升的变化趋势, 且在24~48 h出现峰值。但抗病品种宁RS-1在12~48 h内一直呈上升的趋势, 48 h的表达量超过接种前; 而感病品种APL01在48 h后又回到了接种前水平(图2)。接种前后 Cu/ZnSOD基因在宁RS-1中的表达量均高于APL01, 接种后48 h, Cu/ZnSOD在抗病品种宁RS-1的表达量为感病品种APL01的4.7倍(表3), 但同一品种接种前后表达量变化不大。
FeSOD在抗病品种宁RS-1中的表达一直处于较低水平且变化幅度不大, 只在24 h后略有上升, 但48 h后又恢复到接种前水平。 FeSOD的表达在感病品种APL01中同样处于较低水平, 但显著高于在抗病品种宁RS-1中, 且在感染后呈现上升趋势(图2)。
 | 图2 不同油菜品种接种核盘菌后 Cu/ZnSOD和 FeSOD的表达差异Fig. 2 Expression of Cu/ZnSODand FeSODin different rapeseed varieties infected by S. sclerotiorum |
| 表3 核盘菌诱导下抗、感品种相关基因的表达差异比较 Table 3 Comparison of gene expression between resistant and susceptible varieties infected by S. sclerotiorum |
RMGEI: ratio of max gene expression between before and after inoculations; RMGEV: ratio of maximum gene expression between varieties.
2.3 油菜接种核盘菌后防御相关酶基因的表达差异
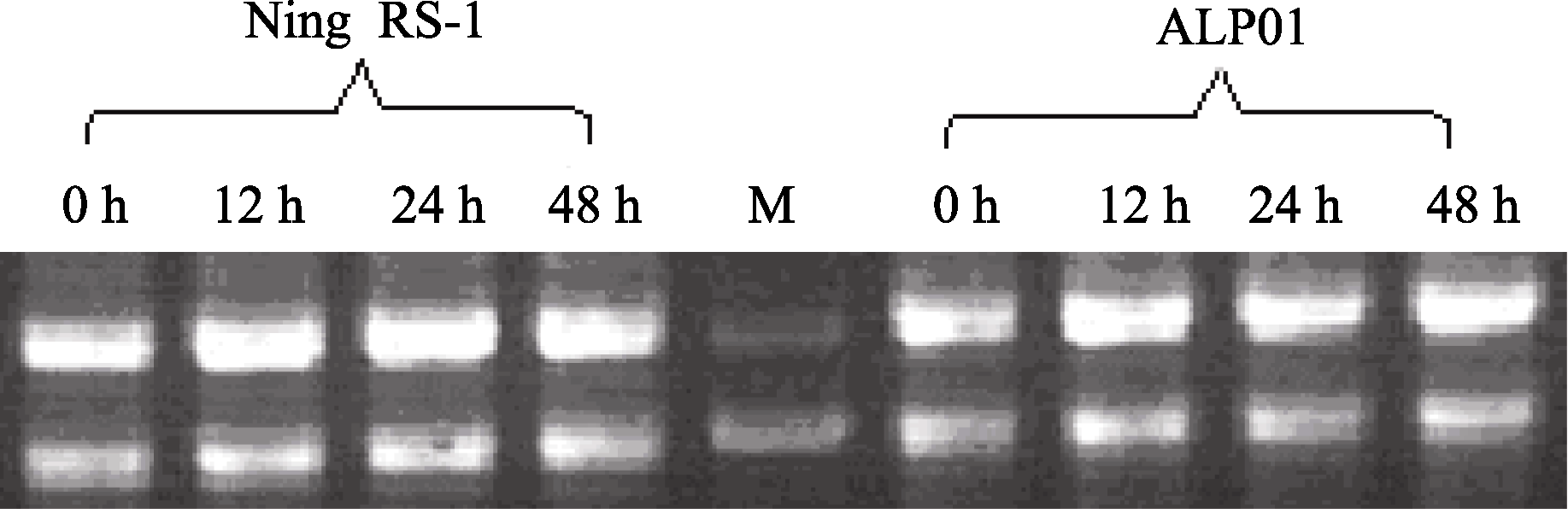
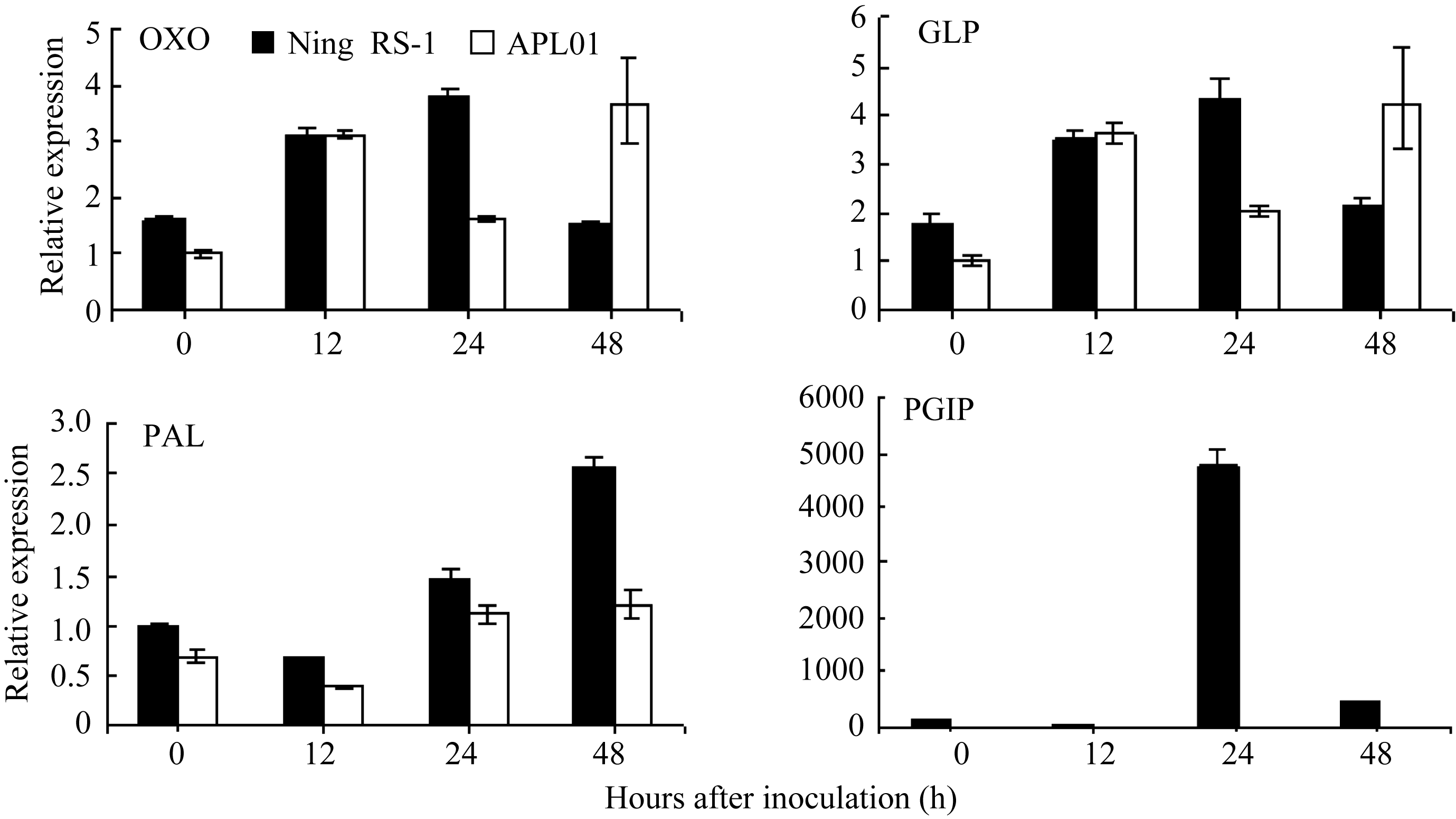
抗病品种宁RS-1在接种24 h内 OXO和 GLP表达量均持续上升, 24 h的表达量分别为接种前的2.3倍和2.4倍, 之后表达量逐渐下降, 感病品种APL01接种核盘菌后, OXO和 GLP表达量均呈现先升后降再升的趋势, 48 h表达量最高, 分别为接种前的3.67倍和3.91倍(图3和表3)。抗、感品种间 OXO和 GLP表达量在时间上存在差异, 而表达量没有显著差异。这说明在抗、感品种间 OXO和GLP诱导防御反应的快慢存在差异。 OXO与 GLP的表达变化趋势在抗、感品种间的表现基本一致, OXO与 GLP基因同源, 同属 GLP家族成员, 这也从侧面证明了2种抗病基因经核盘菌诱导表达的同步性。
PAL基因在同一个品种接种前后表达量差异显著, 抗病品种宁RS-1接种48 h内持续升高, 达接种前的2.6倍, 而感病品种APL01在接种24 h后表达量即不再升高。抗病品种宁RS-1和感病品种APL01在接种诱导48 h内, PGIP的表达均呈先降后升再降的变化趋势, 24 h达最高峰值, 接种前后 PGIP在抗病品种宁RS-1中的表达差异极显著, 24 h表达量为接种前的170.4倍, 而 PGIP在感病品种APL01中表达量在接种前后差异较小, 24 h表达量仅为接种前的3.5倍, 并且 PGIP基因在抗病品种宁RS-1的表达量远高于感病品种APL01, 同时期最高差异达1299.4倍(图3和表3)。
2.4 油菜接种核盘菌后水杨酸(SA)介导的抗病信号相关基因表达差异
接种前 EDS1基因在抗、感品种间的表达量差异不显著; 接种后 EDS1基因在抗、感品种中都能被
 | 图3 不同油菜品种接种核盘菌后 OXO、 GLP、PAL与 PGIP的表达差异Fig. 3 Expression of OXO, GLP, PAL, and PGIP in different rapeseed varieties infected by S. sclerotiorum |
 | 图4 不同油菜品种接种核盘菌后 EDS1和 PR1的表达差异Fig. 4 Expression of EDS1 and PR1 in different rapeseed varieties infected by S. sclerotiorum |
核盘菌诱导表达, 且品种间表达量差异达到显著水平, 其中抗病品种宁RS-1在接种后24 h表达量达最大值, 然后迅速降低, 而感病品种APL01在接种后48 h表达量达最大值, 表明 EDS1作为一种信号分子, 在抗、感品种中可能存在不同的诱导表达途径。接种后48 h内, PR1在抗、感品种间的表达虽在某一时间段略有起伏, 但总体上均处于下调状态, 说明 PR1在核盘菌诱导后表达受到了抑制(图4和表3)。
2.5 油菜接种核盘菌后JA/ET介导的抗病信号相关基因表达差异
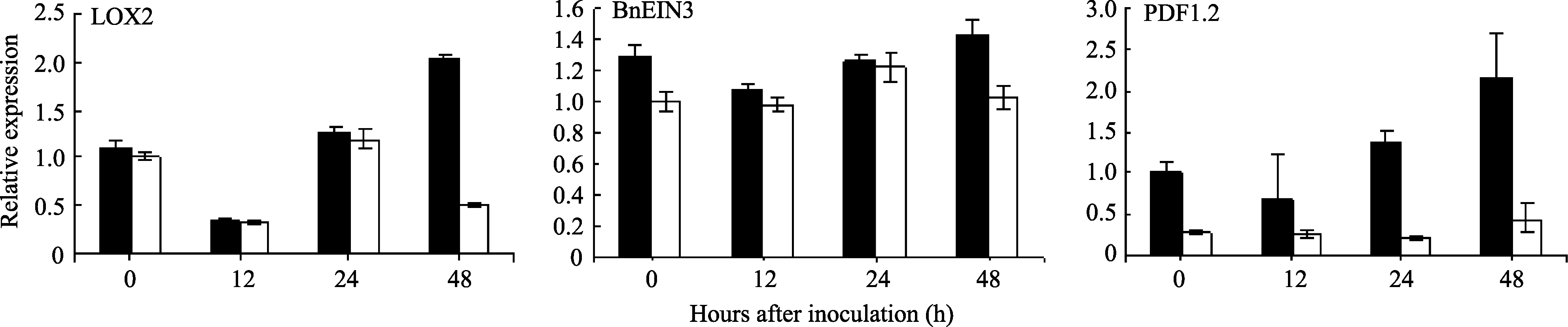
接种后24 h内, 抗、感品种中相关基因表达均呈先降后升趋势, 且表达量基本保持一致, 但在侵染后期, 抗病品种宁RS-1的 LOX2表达量迅速上升, 至48 h达最大值, 表达量是诱导前的2.0倍, 而感病品种APL01的 LOX2表达量在48 h有所下降, 表达量仅为抗病品种的1/4倍。 EIN3的表达量在接种0~48 h内呈先略降后略升的趋势, 但变化不大, 且抗、感品种间的趋势基本一致。抗病品种宁RS-1在接种12 h后, PDF1.2的表达量有所下降, 随后迅速增加, 至48 h表达量达到最大值, 为接种前的2.2倍; 而感病品种APL01在接种前后的 PDF1.2表达量没有明显变化, 一直处于较低水平。由此可见, 核盘菌诱导可激活油菜的JA/ET途径, 抗病品种的 LOX2和 PDF1.2基因表达得到增强(图5和表3)。
3 讨论
3.1 核盘菌诱导下油菜抗氧化相关酶基因的表达
在植物中, Cu/ZnSOD是3种超氧化物歧化酶中含量最丰富的一类, 主要存在于叶绿体、胞质、细胞核和质外体中[ 14]。 FeSOD每个亚基都只含一个金属离子, 从 FeSOD的一些生化指标及前导序列推测, 该酶存在于叶绿体中[ 15]。齐绍武等[ 29]研究表明, 油菜抗、感菌核病品种组织内的 SOD活性差异显著, SOD活性与品种抗性显著相关。接种核盘菌后, SOD活性均发生变化。叶绿体内氧自由基的增加常引起 FeSOD基因表达, 细胞质氧自由基增加会引起细胞质 Cu/ZnSOD基因的表达[ 14]。本研究表明, 接种核盘菌后, FeSOD在抗、感品种间的表达量均较低, 且品种间差异不显著; 而 Cu/ZnSOD在抗病品种中表达增强, 且48 h表达量显著高于感病品种, 这一结果与杨鸯鸯等[ 30]对 Cu/ZnSOD的研究结果相符, 由此可以推断 Cu/ZnSOD基因的表达与活性氧的变化
 | 图5 不同油菜品种接种核盘菌后 LOX、 EIN3和 PDF1.2的表达差异Fig. 5 Expression of LOX, EIN3, and PDF1.2in different rapeseed varieties infected by S. sclerotiorum |
相关, 该基因对清除活性氧的能力比 FeSOD强, 在油菜抗菌核病中具有重要作用。
3.2 核盘菌诱导下与油菜防御相关酶基因的表达
草酸氧化酶( OXO)可以分解病菌分泌的草酸, 生成CO2和过氧化氢(H2O2), 从而限制病菌的生长; 草酸被降解后生成的H2O2是植物和病原微生物互作中快速合成的一种早期活性氧类, 它在植物受病原微生物侵染后可引发一系列反应, 如直接杀死病原菌、作为超敏反应组织者诱导细胞壁结构蛋白的氧化交联从而阻止病菌侵入、诱导植保素生成以及最终诱导系统获得性抗性等[ 31]。GLP被称为类萌发素蛋白, 与草酸氧化酶基因同源, 是多基因编码的一种细胞壁糖蛋白[ 32]。已有研究表明, 在病原菌侵染期间, GLP可能起结构作用, 作为靶标进行蛋白交联加固细胞壁。本研究表明, 接种前后 OXO与 GLP表达量变化趋势基本一致, 且在抗、感品种间的表达差异不显著。根据此前的研究, OXO与 GLP同源, 均属于 GLP基因家族, 因此, 两基因在油菜的防卫反应中可能存在某种协作关系, 另外, 从基因的表达量来看, 在抗病品种中, OXO与 GLP主要在接种后的24 h内表达, 推测两基因可能在防卫反应的前期发挥作用。
研究发现, 植物与病原物互作过程中, 会伴随着 PAL活性的升高, 同时有木质素的积累及酚类物质和植保素的合成, 因此 PAL与植物抗病性有着密切的关系。植物 PAL基因的表达受植物病原物及病菌诱导物乙烯等的诱导调控, 并与品种抗性相关[ 33]。本研究结果表明, 接种核盘菌后, PAL在抗、感品种中均被诱导表达, 后期在抗病品种的表达量迅速上升, 且显著高于其在感病品种的表达水平。由此推测 PAL与植物抗病性有着密切的关系, 抗性品种在接种后期才诱导产生大量的 PAL, 可能是由于激活了乙烯信号途径, 随着乙烯合成的增加导致 PAL的积累。
核盘菌侵染寄主植物时能分泌果胶酶, 尤其是内切-多聚半乳糖醛酸酶(endo-PGs), 而PG是核盘菌导致受感染的植物组织出现水浸状病症的主要原因[ 34], 在病菌感染植物过程中起重要作用, 在多种植物中发现PGIPs是一种细胞壁蛋白, 能够专一性抑制真菌病原菌的PGs。PGIPs抑制真菌的PG, 有利于寡聚半乳糖醛酸的积累, 而寡聚半乳糖醛酸是植物防御反应的激发子[ 35], 激发子通过信号传导途径, 将信号传递到细胞, 诱导 PGIP基因的表达。本研究结果表明, 接种前后 PGIP在抗病品种宁RS-1中表达差异极其明显, 24 h表达量为接种前的170.4倍, 而在感病品种APL01中表达量差异相对较小, 24 h表达量仅为接种前的3.5倍, 同时 PGIP在宁RS-1的表达量远远高于APL01, 24 h达1299.4倍。由此可以推断, PGIP在油菜抵御真菌侵染的防卫反应前期发挥着关键性的作用, PGIP诱导表达产生的PGIPs作为一种糖蛋白能够特异性地识别真菌分泌的PGase, 并与其形成二聚体复合物, 从而抑制了PGase的活性, 最终延缓甚至抑制了病原真菌对植物细胞壁的降解, PGIP的这种生物学特性决定了它在植物抵御真菌病害的发生中起着重要作用。这也证实了周晓婴等[ 36]的研究结果。
3.3 核盘菌诱导下与油菜防卫反应信号传导相关的基因表达
核盘菌侵染拟南芥时激活了JA/ET防卫途径, 而非SA途径。一些研究者提出, 植物防卫反应中依赖于SA途径的防卫反应作用于活体营养病原菌, 而依赖于JA/ET途径的防御反应作用于死体营养病原菌。并且SA和JA途径各自诱导表达的一些基因会相互抑制[ 37]。另有研究表明, JA能够抑制依赖SA途径的标志基因 PR的表达[ 37]。本研究中抗病品种的 PDF1.2基因可被核盘菌诱导而增强表达, 但在感病品种中则没有显著变化, 一直处于较低水平。接种病菌后, PR1在抗、感品种间的表达总体上均处于下调状态, 推测核盘菌接种可能激活了油菜JA/ET信号途径, 同时抑制了SA信号途径。
任秋红的研究表明, 在油菜中核盘菌能激活 LOX2基因的表达, 受茉莉酸正调控[ 38]。刘胜毅等[ 23]认为油菜 EIN3及拟南芥 EIN3基因在对死体营养型病原菌的抗性中发挥着重要作用, 植物可能通过上调 EIN3基因的表达来抵御病原菌的侵染。本研究结果表明, 核盘菌诱导抗病品种中 LOX、 EIN3表达量的变化趋势与JA/ET途径的标记基因 PDF1.2保持一致, 且均在48 h达最大值, 其中 LOX、 PDF1.2在抗病品种中的表达量明显高于感病品种, 由此推测, LOX、 EIN3可能参与了油菜JA/ET介导的抗病信号的启动与传导, LOX、 PDF1.2的高表达在油菜对菌核病抗性中起重要作用, 油菜可能通过上调 LOX2、 PDF1.2基因的表达来抵御病原菌的侵染。
EDS1是一种信号分子, 其编码的蛋白与脂肪酶同族, 可抵制植物对活体营养病原菌产生抗性(包括HR的产生)[ 39], 但其调控产生的HR反应有利于死体营养病原菌核盘菌在油菜上定殖, 接种后期感病品种 EDS1上调表达可能是核盘菌利用了SA与JA/ET信号的抑制效应以增加致病性[ 40], 同时也是SA防卫途径的激发子和JA/ET的抑制子[ 20]。Kamal等[ 41]发现, B. cinerea激活了 EDS1的表达, 将该基因沉默后, 烟草对 B. cinerea抗性得到增强。本研究发现, 接种核盘菌后, 抗病品种 EDS1基因先是被诱导表达, 24 h达最大值, 随后表达水平降低, 恢复到接种前水平。感病品种在接种病菌后, EDS1基因的表达水平先下调, 随后一直上调。推测抗病品种被核盘菌诱导后, 随着防御反应体系的激活, 抑制了 EDS1基因的表达, 从而减少细胞死亡的程度。
4 结论
PGIP、 Cu/ZnSOD、 OXO、 GLP、 LOX2、 PDF1.2等基因可能与油菜抗菌核病防卫反应相关, 推测抗病品种宁RS-1对菌核病的抗性可能是由于PGIP的上调表达, 抑制了核盘菌PG蛋白对侵染部位油菜组织细胞壁的降解, 从而抑制了油菜菌核病的发生与蔓延。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|