{kind=link}
高丹草遗传效应与杂种表现预测模型
引用本文
逯晓萍, 刘丹丹, 王树彦, 米福贵, 韩平安, 吕二锁. 高丹草遗传效应与杂种表现预测模型. 作物学报, 34(3): 466-475
LU Xiao-Ping, LIU Dan-Dan, WANG Shu-Yan, MI Fu-Gui, HAN Ping-An, LÜ Er-Suo. Genetic Effects and Heterosis Prediction Model ofSorghum bicolor ×S. sudanense Grass . 作物学报, 34(3): 466-475 
Permissions
LU Xiao-Ping, LIU Dan-Dan, WANG Shu-Yan, MI Fu-Gui, HAN Ping-An, LÜ Er-Suo. Genetic Effects and Heterosis Prediction Model of
Copyright©2014, 作物学报编辑部
作物学报编辑部
高丹草遗传效应与杂种表现预测模型
摘要
以5个高粱不育系材料为母本, 18个优质苏丹草材料为父本, 按照遗传交配设计(NCII)配制成90个杂交组合。分别在内蒙古呼和浩特和包头两地, 利用与高丹草产量相关的QTL位点标记检测亲本间的遗传差异, 并将F1的8个性状表型值对亲本材料进行标记位点的筛选, 建立标记效应和标记型值估算体系。估算特异性位点对性状表现的效应及杂种标记型值, 进而分析杂种标记型值与杂种表现的相关性, 应用逐步回归分析法建立8个性状杂种表现的预测模型, 并通过Jackknife抽样技术检测模型的精确度和稳定性。结果显示, 在分别考虑显性、加性作用下8个性状的标记型值与表型值的相关系数平均为0.65, 各性状的可决系数较大(0.51~0.88), 而且两地结果趋势一致, 表明该预测模型稳定性强, 精确度较高。该模型对高丹草的杂种表现预测以及亲本选配都具有一定的指导意义。
关键词:
高丹草; 遗传效应; AFLP; SSR; SRAP; 杂种优势; 预测模型
Genetic Effects and Heterosis Prediction Model ofSorghum bicolor ×S. sudanense Grass
Abstract
Molecular markers-based heterosis prediction can provide some advices for
Keyword:
Sorghum bicolor ×; S. sudanense; Genetic effect; AFLP; SSR; SRAP; Heterosis; Prediction model
高丹草是苏丹草[ Sorghum sudanense(Piper Stapf)]与高粱[ Sorghum bicolorL. (Moench)]杂交后产生的F1代种间杂交种, 由于两者的亲缘关系较远, 性状差异较大, 杂交后的F1结合了二者的优点, 表现出强大的杂种优势[ 1, 2], 是一种综合农艺性状优良及应用广泛的一年生饲用牧草[ 3, 4]。
近年来, 国外在高丹草育种方面的研究较多, 有许多国家的科研机构都在进行高产优质高丹草新品种的选育研究。例如加拿大育成了佳宝, 日本育成了高产优质多抗的格林埃斯等。我国自20世纪50年代开始引入并进行选育工作, 但学术界直到20世纪90年代才开始重视高丹草的研究[ 5, 6, 7]。目前, 我国对高丹草的研究取得了长足的进展[ 8, 9, 10]。国内育成了蒙农青饲1号、2号和3号[ 11, 12, 13]以及皖草3号[ 14]等品种。
如何迅速而准确地筛选优良亲本、预测所配组合的杂种优势是作物育种者关心和致力研究的问题。分子标记技术的发展为杂种优势的研究提供了新手段。目前, 利用分子标记揭示品种间遗传差异, 进而预测杂种优势已有大量的文献报道, 但尚未得出一致的结果。如赵庆勇等[ 15]和张培江等[ 16]在水稻中研究认为分子标记遗传距离与杂种优势之间呈极显著正相关; 而张涛等[ 17]的研究却认为分子标记遗传距离与F1杂种优势之间没有明显的相关性。袁力行等[ 18]在玉米研究中认为分子标记遗传距离与F1产量杂种优势呈显著正相关, 但相关程度还不足以预测杂种优势。张启发等[ 19]则认为当亲本间特殊杂合性与杂种优势的相关系数较大时, 可用于杂种优势预测, 但这种关系会因亲本的遗传多样性及杂种优势遗传机制的复杂性而改变。近年来, 张涛等[ 20]提出利用功能基因标记分析亲本的遗传距离, 研究发现功能基因标记遗传距离与杂种优势呈显著正相关。目前, 在高丹草研究中已定位了与产量相关的
QTL位点[ 21]。因此, 本研究在对高丹草遗传图谱构建和产量性状QTL定位以及重组自交系构建与高产种质的创新、近等基因系研究的基础上[ 21, 22, 23, 24, 25, 26, 27], 利用与高丹草产量相关的QTL位点标记(表1)检测亲本间的遗传差异, 并应用逐步回归分析法建立基于加性-显性效应的高丹草杂种表现分子标记预测模型。并对得到的预测模型, 采用Jackknife抽样技术进行检验评价, 确定其稳定性和精确度。旨在建立一个简便而有效的预测指标体系指导高丹草杂种优势育种中亲本的选配, 同时也为以标记型值预测其它作物杂种表现提供一种有效的新途径。
1 材料与方法
1.1 试验材料及设计
2011年以5个高粱不育系(不育系值由其配套的同核异质保持系代替)和18个优良苏丹草及其杂种后代材料为亲本, 分别在内蒙古呼和浩特和林县(HH)和包头萨拉齐(BT)环境下种植, 种植标准相同, 按照NCII设计, 配制了90个杂交组合(不育系和亲本的具体名称及来源见表2)。每个材料种植3个重复, 4行区, 行长5.0 m, 行距0.4 m。在高丹草成熟时期,
| 表1 与高丹草产量相关的QTL标记 Table 1 Yield related QTLs markers on sorghum-sudan grass |
| 表2 供试材料的编号及来源 Table 2 Accession number and sources of experiment materials |
取30株测定其株高、茎重、穗长、分蘖数、叶长、叶宽、成株期叶片数、单株鲜重共8个农艺性状指标。
1.2 分子标记的检测和统计
1.2.1 DNA的提取及检测 在高丹草3~4片嫩叶时取样, 采用TIANGEN-植物基因组试剂盒提取高丹草叶片基因组DNA。对提取出的DNA用1%的琼脂糖凝胶电泳检测其完整性及浓度。
1.2.2 分子标记分析 采用具有代表性的AFLP、SSR和SRAP分子标记(分布在整个基因组的AFLP和SSR以及分布在开放阅读框的SRAP), 且是与高丹草产量相关的QTL标记[ 21, 22, 23, 24, 25], 根据引物和扩增片段大小排序编码, SSR扩增位点编码形式为Sx, AFLP扩增位点编码形式为ExMy-z: 其中E和M分别代表上下游引物, x、y为引物编号, z为按扩增片段大小所排序号; SRAP扩增位点编码形式为MxEy-z, 其中M和E分别代表上下游引物, x、y为引物编号, z为按扩增片段大小所排序号。
1.2.3 标记位点筛选 所有亲本材料都经过9代以上的自交或姊妹交, 可以认为所有亲本材料在各位点上是纯合的, 根据3组法筛选, 以各条带为因子, 杂种子代按在双亲间存在的标记状态的差异性分为3组: 1/1组, 即在此位点上, 某条带在杂交双亲中均出现; 1/0组, 即在此位点上, 某条带只在一个亲本中出现; 0/0组, 即在此位点上, 某条带在杂交双亲中均不出现。采用单向分组的方差分析法, 对3组性状表型均值进行 t测验, 筛选与本研究中高丹草农艺性状相关的标记位点。
1.3 标记效应值的估计
根据3组标记型的杂种F1性状表现均值差异显著性, 估算该标记位点的加性效应 a和显性效应 d, 因而可得到其计算方法 a=1/2( 
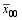
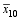

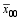
根据第 i和第 j个杂交亲本在第 h个标记位点的标记型和该位点的加性、显性效应, 该标记位点对该杂种F1的实际效应 gwh= a( Wih- Wjh-1) d( Wih- Wjh)。其中, Wih和 Wjh分别是第 i和第 j个杂交亲本在第 h个标记位点的标记型, 取值为“0”或“1”。
在加性、显性情况下, 各位点间的效应是可以累加的, 即符合线性可加性原则, 杂交亲本及杂种的标记型值
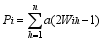
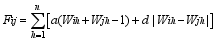
式中, Fij是亲本 i和亲本 j的杂种F1的标记型值, n为与性状相关的特异标记位点总数。
1.4 加性-显性效应的杂种表现预测模型的建立
性状基因型值( G)和表型值( P)的计算方法如下: 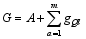

其中, A为常数, gQl为第l个QTL位点的效应, m为
与该性状有关的QTL数目, E为随机误差。
采用逐步回归法利用实际标记效应对性状表型值建立预测模型 y= Xβ+ ε。
1.5 Jackknife抽样检验
利用近年来广泛应用于模型评价和参数估计的Jackknife抽样技术对所得到的预测模型进行检验评价。将两地的数据合并分析, 每次从90个杂交组合中剔除一个, 计算可决系数和预测残差, 评价预测模型的稳定性和精确度。
2 结果与分析
2.1 性状相关标记的筛选
利用SSR、AFLP、SRAP标记扩增23个亲本获得的多态性位点数(两地合并)分别为126、283、330, 将其采用3组法进行筛选得到与各农艺性状指标相关的标记位点(表3)。
由3可以看出, 3种分子标记在0.01显著水平下扩增出的615个标记位点中, 筛选出与8个农艺性状相关的位点496个, 占总位点数的80.6%。平均每个特异性位点与2.61个农艺性状相关。在显著水平 α=0.001的条件下, 各性状的相关位点数目有了较大程度的降低, 在所有的标记位点中, 筛选出了246个与8个农艺性状相关的位点。
在筛选出的位点中, 只表现加性效应的位点称为加性位点; 只表现显性效应的称为显性位点, 还有少量数目的位点既有加性效应又具有显性效应, 称为加-显性位点。其数目为显性位点>加性位点>加-显位点。
| 表3 加性、显性位点的筛选结果 Table 3 Selection result of specific DNA marker sites |
为了建立稳定性和精确度高的预测模型, 利用 α=0.001显著水平下筛选到的特异位点计算后续各项效应指标。
2.2 标记型值与杂种表现的相关分析
2.2.1 标记效应值的估计 在对8个农艺性状特异性标记位点的筛选过程中(表3)发现以显性作用为主。两地的表现趋势一致, 以单株鲜重为例将两地结果合并分析见表4。
从表中可以发现, 在与单株鲜重相关的29个特异性标记位点中, 13个具有加性作用, 16个具有显性作用。没有出现加-显性作用的位点。13个加性位点的标记效应值在-0.1475~0.2685之间, 16个显性位点的标记效应值在-0.2195~0.1635之间。
每个性状受到加性作用和显性效应影响的程度,用母本高粱和父本苏丹草以及二者的互作方差评估, 结果见表5。经 F检验, 除高粱的单株鲜重和茎重之外, 所有母本因素(高粱)和父本因素(苏丹草)以及互作的方差均达到极显著水平( P≤0.001)。高粱和苏丹草的株高、叶长、成株叶片数等性状的方差同时显著高于互作方差, 其他性状如穗长、分蘖数、叶宽均以高粱的方差为优势。所有性状的亲本方差均值大于互作方差。说明在高丹草杂种表现中显性作用强于加性作用。
| 表4 单株鲜重相关位点及标记效应值 Table 4 Loci and the value of the marker effect for fresh weight |
| 表5 高丹草8个农艺性状的方差分析结果 Table 5 Variance of eight agronomic traits of sorghum-Sudan grass |
2.2.2 杂种标记型值与杂种表现的相关性分析
按照线性可加原理计算各个亲本及杂交子代的标记型值和特异性位点的加-显性效应值。以单株鲜重为例, 表6列出了其在0.001显著水平下的各个亲本及子代的标记型值。通过分析发现, 23个亲本的标记型值在-0.5445~0.7825之间变化, 其平均值为0.0156; 90个杂种子代的标记型值的范围为-1.3496~1.8897, 平均值为0.6836。杂交亲本的单株鲜重的变异幅度低于其杂种子代F1。
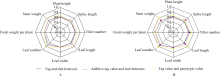
当只考虑显性作用时, 对8个性状的标记型值和其表型值进行相关性分析, 各个性状的显性标记型值与其表型值之间呈显著的正相关, 相关系数在0.5745~0.7213之间; 当考虑加性作用时, 相关系数在0.5906~0.7033之间, 虽然相关系数有所变化, 但是波动范围并不是很大, 说明各个性状的杂种表现受加性作用和基因互作效应的影响并不很大, 更主要的还是显性效应的影响大一些(图1)。从图1可以看出, 以基因型值进行筛选时, 标记型值与基因型值间的相关性在8个性状上都有很大的提高。相关系数在0.6136~ 0.8272之间, 而且两地结果趋势一致, 说明基因型值相对于性状表型值可以更准确地反映杂种性状表现, 这是由于基因型值排除了部分随机误差等原因。
| 表6 亲本及子代F1单株鲜重标记型值 Table 6 Marker effect values of plant fresh weight in F1 and their parents |
 | 图1 杂种标记型值与性状表型值的相关性A代表呼和浩特的样本数据分析; B代表包头的样本数据分析。Fig. 1 Correlation between marker-type values of hybrid and their phenotypeA: data analysis came from Huhhot sample; B: data analysis came from Baotou sample. |
2.3 杂种表现预测模型建立
根据杂种标记型值与杂种表现的相关性分析可知, 杂种表现主要是受加显性效应的控制, 而与互作效应的关系较小, 因此, 在建立杂种表现分子标记预测模型时, 可不考虑互作效应的影响。
采用多元逐步回归分析方法, 用显著水平 α=0.001条件下筛选到的特异位点数目, 以90个杂种子代的各个特异性标记位点的标记效应值为自变量, 以杂种表现性状值为因变量, 构建了8个性状的预测模型回归方程(表7)。
在8个性状预测模型中, 各个方程所涉及的标记位点在6~12个不等, 其中, 穗长只含有6个标记位点; 叶长含有12个标记位点。而叶宽和茎重的预测模型中有2个共同位点S75-6和S49-6; 叶长和单株鲜重有一个共同位点E39M51-2。标记效应和各个性状之间的复相关系数(0.609~0.936)较高, 说明两组变量之间的线性相关程度较高, 进一步证明建立多元回归线性方程是科学的, 也是可实现的。
| 表7 性状表现预测模型 Table 7 Traits of the prediction model |
3 讨论
提高遗传效应预测杂种优势水平的重要因素是增加与杂种表现相关的QTL连锁的位点数目, 而且除了有一定的数目外, 每个位点的贡献率对预测结果也有一定的影响, 与QTL连锁位点的贡献率越低, 预测的效果越差。这一点在受大量微效多基因控制的产量性状上表现尤为突出[ 31, 32, 33]。所以, 对杂种表现进行预测研究时应尽量筛选出贡献率高的位点。游书梅等[ 34]利用14个与子粒相关QTL标记, 分析10个杂交水稻亲本的遗传差异, 进而分析遗传距离与千粒重及千粒重杂种优势的相关性, 结果显示, 遗传距离与千粒重和对照之间呈显著正相关, 与母本优势之间呈极显著正相关, 表明利用与子粒相关的QTL标记来预测杂交水稻的千粒重及其杂种优势是可行的。本研究采用的标记是经过构建遗传图谱及基因定位[ 21, 22, 23, 24, 25]后筛选得到的与8个产量相关性状连锁的标记, 并且具有一定的贡献率, 所以, 研究
| 表8 预测模型的评价 Table 8 Evaluation of prediction models |
结果与前人研究[ 35]相比, 各性状的决定系数 R2都明显提高。另外, 由于所研究的性状均为数量性状, 而数量性状由于受环境的修饰作用大, 因此, 为了提高预测模型的精确度, 本研究在试验设计上安排了2个不同环境, 使研究结果更加可靠。
利用分子标记与QTL连锁分析可以提供与杂种优势相关的信息, 鉴定与杂种优势相关的标记位点, 确定亲本在QTL上的差异, 可以有效预测杂种优势。Vuylsteke等[ 36]在对玉米杂种表现预测研究中应用QTL效应分析取得了较好的预测效果。
采用遗传效应预测杂种优势比遗传距离预测有更大的优势, 因为预测需要的标记位点较少, 因而可以排除部分干扰, 所以在育种中应用的可行性强。本研究对各个亲本及子代的标记型取值都是在0.001显著水平下进行, 并在分别考虑显性作用和加性作用下对标记型值与其表型值进行相关分析, 结果显示, 8个性状的标记型值与表型值的相关系数平均值为0.65, 标记型值与基因型值间的相关系数(0.6136~0.8272)也较高, 而且两地结果趋势一致, 说明基因型值相对于性状表型值可以更准确地反映杂种性状表现。
利用近年来广泛应用于模型评价和参数估计的Jackknife抽样技术对所得预测模型的检验评价显示, 各性状的决定系数较大(0.51~0.88), 且其剩余误差和预测残差都小于调查数据的随机误差, 说明预测模型的精确度较高、稳定性较强。本研究建立的遗传预测模型是来自两个不同环境条件下的结果, 因此, 可在一定范围内指导育种中亲本的选配。
4 结论
建立了基于加性-显性效应的杂种表现分子标记预测模型。经过对模型的精确度和稳定性的检测表明, 该模型对高丹草的杂种表现预测以及亲本选配具有一定的指导意义。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|