


{kind=link}
{kind=link}
{kind=link}
{kind=link}
1,2,4-三氯苯胁迫对水稻分蘖盛期植株生长和生理特性的影响
引用本文
丁秀文, 张国良, 戴其根, 朱青. 1,2,4-三氯苯胁迫对水稻分蘖盛期植株生长和生理特性的影响. 作物学报, 34(3): 487-496
DING Xiu-Wen, ZHANG Guo-Liang, DAI Qi-Gen, ZHU Qing. Effects of 1,2,4-trichlorobenzene on Growth and Physiological Characteristics of Rice at Maximum Tillering Stage. 作物学报, 34(3): 487-496
Permissions
DING Xiu-Wen, ZHANG Guo-Liang, DAI Qi-Gen, ZHU Qing. Effects of 1,2,4-trichlorobenzene on Growth and Physiological Characteristics of Rice at Maximum Tillering Stage. 作物学报, 34(3): 487-496
Copyright©2014, 作物学报编辑部
作物学报编辑部
1,2,4-三氯苯胁迫对水稻分蘖盛期植株生长和生理特性的影响
摘要
-1) TCB胁迫下, 最长根长、地上部和地下部干物重极显著增加, 当中高浓度(40 mg kg-1、60 mg kg-1) TCB胁迫时, 最长根长、株高、单穴分蘖数、地上部和地下部干物重显著下降。不同浓度TCB胁迫下, 敏感基因型宁粳1号根系活力、叶绿素含量、叶片和根系可溶性蛋白质含量相对较低, 叶片和根系超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性明显降低, ]]>-1) TCB胁迫下, 根系活力、叶片和根系的可溶性蛋白含量、SOD、POD、CAT活性极显著增加, ]]>
-1) TCB胁迫时, SOD、POD、CAT活性虽然增强, 但]]>
-1) TCB胁迫时, 其根系活力、叶绿素含量极显著降低, 根系中SOD、CAT活性显著降低, ]]>
关键词:
水稻; 分蘖盛期; 1; 2; 4-三氯苯; 毒性; 生长发育; 抗氧化酶
Effects of 1,2,4-trichlorobenzene on Growth and Physiological Characteristics of Rice at Maximum Tillering Stage
Abstract
A soil culture experiment was conducted to study the effects of 1,2,4-trichlorobenzene (TCB) on morphological and physiological indexes of rice at maximum tillering stage, using rice cultivars of Ningjing 1 (TCB sensitive genotype) and Yangfujing 8 (TCB tolerant genotype), with four treatments of TCB concentrations including 0 (CK), 20, 40, and 60 mg kg-1. The results indicated that the maximum root length, plant height, tillers per hill, shoot and root dry weight of Ningjing 1 were all decreased significantly (
Keyword:
Rice; Maximum tillering stage; 1; 2; 4-trichlorobenzene; Toxicity; Growth and development; Antioxidant enzymes
氯苯类化合物(CBs)有广泛的工业和家庭用途, 常被用作化学合成(农药、染料)的中间体、溶剂、卫生用品及电解液的组成成分[ 1]。这类化合物易挥发、化学性质稳定, 是环境中分布广、难降解的一类环境污染物, 已被我国列入“水中优先控制污染物黑名单”[ 2]。CBs的辛醇-水分配系数较高、苯环上氯原子数目较多, 故能通过食物链转移、积累并放大, 进而危害人类健康[ 3, 4]。
1,2,4-三氯苯(TCB)是CBs中常见的一种, 除了上述工业用途之外, 在农业中常作为除草剂的生产原料, 也是农药六六六、林丹等的降解产物[ 5]。我国是三氯苯的生产大国[ 6], 由于它的广泛应用, TCB已在我国土壤[ 7, 8]、城市污泥[ 9]、地表水[ 10, 11]与地下水[ 12]、表层沉积物[ 4, 13]、水生生物[ 4]及陆生植物[ 7, 14]中检出。因其具有致畸、致癌、致突变作用, 已被美国环保局(EPA)列入优先控制污染物名单[ 15]。我国国家标准生活饮用水卫生标准规定TCB总量不得超过0.02 mg L-1 [ 16], 北京市将TCB列入水污染物排放标准, 并明确限定它的排放量最高不得超过0.6 mg L-1 [ 17]。
经农田灌溉、污泥农用、农药使用和挥发扩散沉降[ 18, 19]等多种渠道, TCB直接或间接进入农田生态系统。水稻是我国主要的粮食作物之一, 在全国粮食生产与消费中占主导地位。分蘖期是水稻生长发育的重要时期之一, 此期内营养生长迅速, 直接影响单株分蘖数和总穗数, 对水稻产量起着决定性作用。以往TCB胁迫对作物生理机制影响的研究主要集中在苗期[ 5, 20, 21, 22, 23, 24, 25], 抽穗期也有少量报道[ 26], 至今未见水稻分蘖期生长及对TCB胁迫生理响应的报道。本文在前期筛选出的耐TCB胁迫代表品种[ 27]的基础上, 研究水稻分蘖盛期对TCB胁迫的生态毒性与生理响应, 以期为分蘖盛期水稻抗逆栽培管理提供依据。
1 材料与方法
1.2 试验设计
选用长×宽×高为60 cm×50 cm×25 cm的塑料周转箱, 每箱装土35 kg, 加水浸泡, 施尿素2 g作为基肥, 调节pH值在5.0~5.5之间。
设置0、20、40、60 mg TCB kg-1干土4个处理, 每个处理3次重复。将TCB溶液(无水乙醇作为溶剂)加入土中, 搅拌均匀, 放置3 d后, 将秧苗(三叶一心)移栽进周转箱。行株距为10 cm × 8 cm, 双本栽。栽培管理与常规高产栽培一致。
1.3 测定项目与方法
1.3.1 形态指标的测定 在水稻分蘖盛期取代表性植株3穴, 用直尺测量最长不定根根长; 每穴取10根长势基本一致的不定根, 排列整齐后用游标卡尺测其根粗, 取平均值; 用米尺测量株高; 将单穴植株地上部分和地下部分在105℃烘箱内杀青15 min, 然后80℃烘至恒重后分别称取干重。
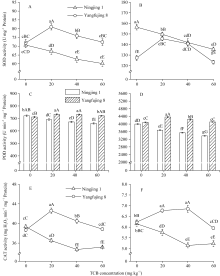
1.3.2 生理指标的测定 采用α-萘胺法测定根系活力[ 28], 单位为μg h-1 g-1 FW; 采用SPAD-502型叶绿素计测定叶绿素含量; 采用考马斯亮蓝法测定叶片和根系的蛋白质含量[ 28], 以mg g-1 FW为单位; 采用氮蓝四唑(NBT)光还原法[ 29]测定超氧化物歧化酶(SOD)活性, 以抑制反应50%的酶量为一个酶活性单位(U), 酶活性单位以U mg-1 Protein表示; 采用愈创木酚法测定过氧化物酶(POD)的活性[ 28], 以每分钟 A470值变化0.01作为一个酶活性单位(U), 酶活性以U min-1 mg-1 Protein表示; 采用碘量法测定过氧化氢酶(CAT)活性[ 30], 以每分钟分解H2O2的毫克数表示酶活性大小, 单位为mg H2O2 min-1 mg-1 Protein。采用硫代巴比妥酸(TBA)法测定丙二醛(MDA)含量[ 31], 单位为nmol g-1 FW; 用羟氨氧化法[ 32]测定超氧阴离子自由基( 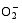
1.4 数据分析与作图
用SPSS16.0进行方差分析, Duncan’s测验法多重比较, Origin7.5软件作图。
2 结果与分析
2.1 TCB胁迫对两种基因型水稻分蘖盛期形态指标和干物重的影响
TCB胁迫下, 除了20 mg kg-1 TCB处理下扬辐粳8号的株高、分蘖数与对照(CK)差异不显著以外, 其他处理下两个基因型水稻分蘖盛期最长根长、根粗、株高、单穴分蘖数与对照差异显著(图1和表1)。敏感基因型宁粳1号的最长根长、株高、单穴分蘖数、地上部和地下部干物重均随TCB胁迫浓度的增加而显著减少, 耐性基因型扬辐粳8号这5个指标在低浓度(20 mg kg-1) TCB胁迫下显著增加, 中高浓度(40 mg kg-1和60 mg kg-1) TCB胁迫下显著递减, 且递减幅度均小于宁粳1号。低浓度TCB胁迫下, 宁粳1号地上部和地下部干物重比未进行TCB胁迫时分别降低2.9%和9.3%; 而扬辐粳8号分别增加0.6%和1.7%。中高浓度TCB胁迫对宁粳1号地上部和地下部干物重(与未进行TCB胁迫相比)的降幅均大于扬辐粳8号。2个基因型水稻的根粗随着TCB胁迫浓度的增加而显著递增, 且中高浓度(40 mg kg-1和60 mg kg-1) TCB胁迫下, 敏感基因型宁粳1号根粗的增幅大于耐性基因型扬辐粳8号。说明低浓度TCB对耐性基因型扬辐粳8号的生长有一定的刺激作用, 能促进其生长; 而当TCB浓度过高时, 扬辐粳8号的生长也受到显著抑制。宁粳1号对TCB胁迫的耐性较差, 表现为生长延缓, 植株矮小, 根系变黑。
 | 图1 TCB胁迫下两种基因型水稻分蘖盛期的生长情况Fig. 1 Phenotypes of the two rice genotypes grown under TCB stress at maximum tillering stage |
2.2 TCB胁迫对2种基因型水稻分蘖盛期根系活力的影响
由图2-A可知, 未进行TCB胁迫时, 两种基因型水稻的根系活力差异不显著; 相同浓度TCB胁迫下, 扬辐粳8号的根系活力极显著高于宁粳1号。宁粳1号的根系活力在中高浓度TCB胁迫下极显著下降, 并呈递减趋势。扬辐粳8号的根系活力在低浓度(20 mg kg-1) TCB胁迫下极显著增加, 40 mg kg-1 TCB胁迫时, 与对照差异不显著, 而当TCB浓度达到60 mg kg-1时, 其活力极显著降低。
2.3 TCB胁迫对2种基因型水稻分蘖盛期叶片叶绿素含量的影响
未进行TCB胁迫时, 2种基因型水稻的叶片叶绿素含量差异不显著; 相同浓度TCB胁迫下, 扬辐粳8号叶绿素含量极显著高于宁粳1号(图2-B)。当TCB胁迫浓度大于20 mg kg-1后, 两种基因型水稻的叶绿素含量均随TCB胁迫浓度增加极显著递减, 且宁粳1号的下降幅度明显大于扬辐粳 8号。
2.4 TCB胁迫对两种基因型水稻分蘖盛期可溶性蛋白质含量的影响
未进行TCB胁迫时, 两种基因型水稻叶片和根系可溶性蛋白质含量差异不显著; 相同浓度TCB胁迫下, 扬辐粳8号叶片和根系可溶性蛋白含量均极显著高于宁粳1号(图3)。宁粳1号叶片和根系可溶性蛋白含量随TCB胁迫浓度的增加而极显著递减。扬辐粳8号叶片可溶性蛋白含量均极显著高于对照, 其叶片可溶性蛋白含量在中低浓度TCB胁迫下呈递增趋势, 60 mg kg-1下有所下降; 而根系可溶性蛋白含量在中高浓度TCB胁迫下均极显著高于对照。
| 表1 TCB胁迫对2种基因型水稻分蘖盛期各形态指标的影响 Table 1 Effect of TCB on morphological index in the two rice genotypes at maximum tillering stage |
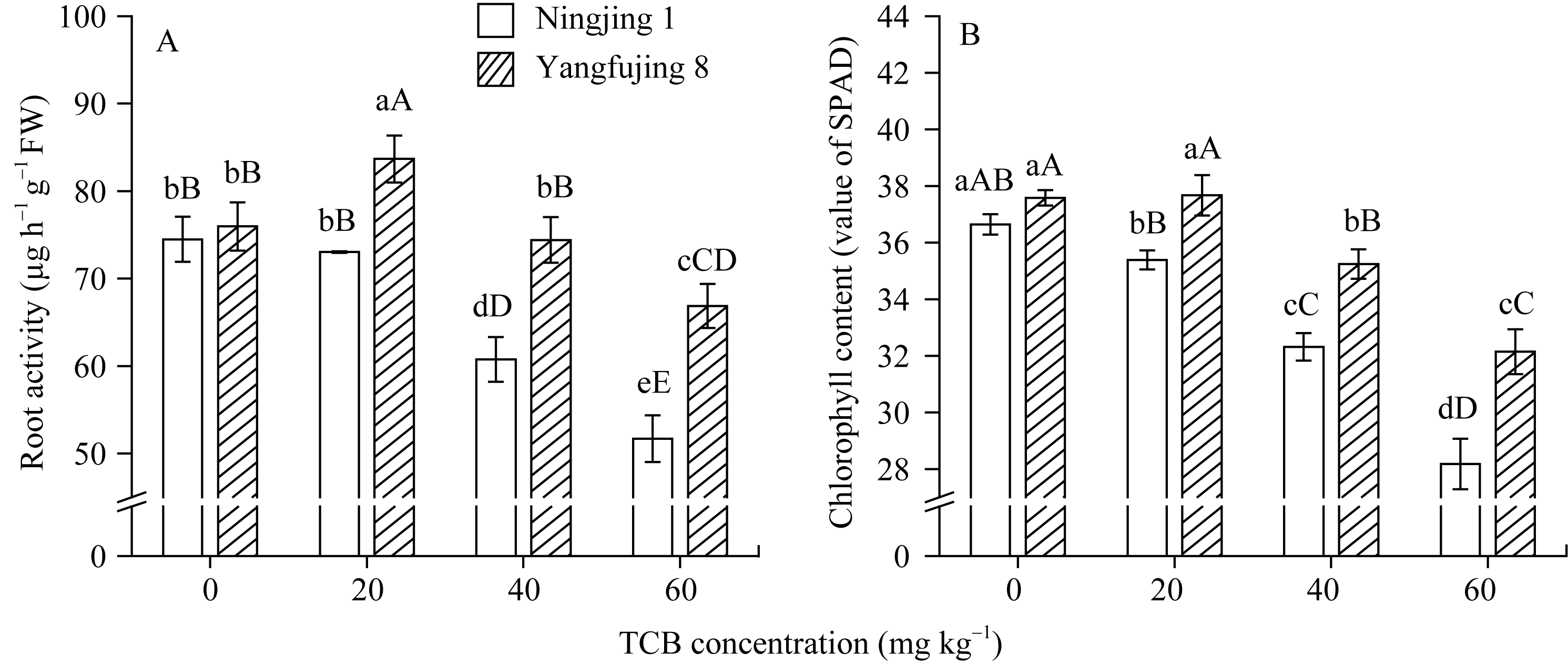 | 图2 TCB胁迫对分蘖盛期水稻根系活力和叶片叶绿素含量(SPAD值)的影响柱上标以不同大、小写字母时分别表示0.01和0.05差异显著水平。Fig. 2 Effect of TCB on root activity and chlorophyll content (value of SPAD) of leaves in the two rice genotypes at top tillering stageBars superscripted by different letters are significantly different at 0.01 (capital) and 0.05 (lowercase) probability levels, respectively. |
40 mg kg-1) TCB胁迫下, 扬辐粳8号叶片和根系SOD活性均显著高于对照, 且从20 mg kg-1开始呈递减趋势。高浓度TCB胁迫下叶片SOD活性与对照差异不显著, 根系SOD活性显著低于对照。
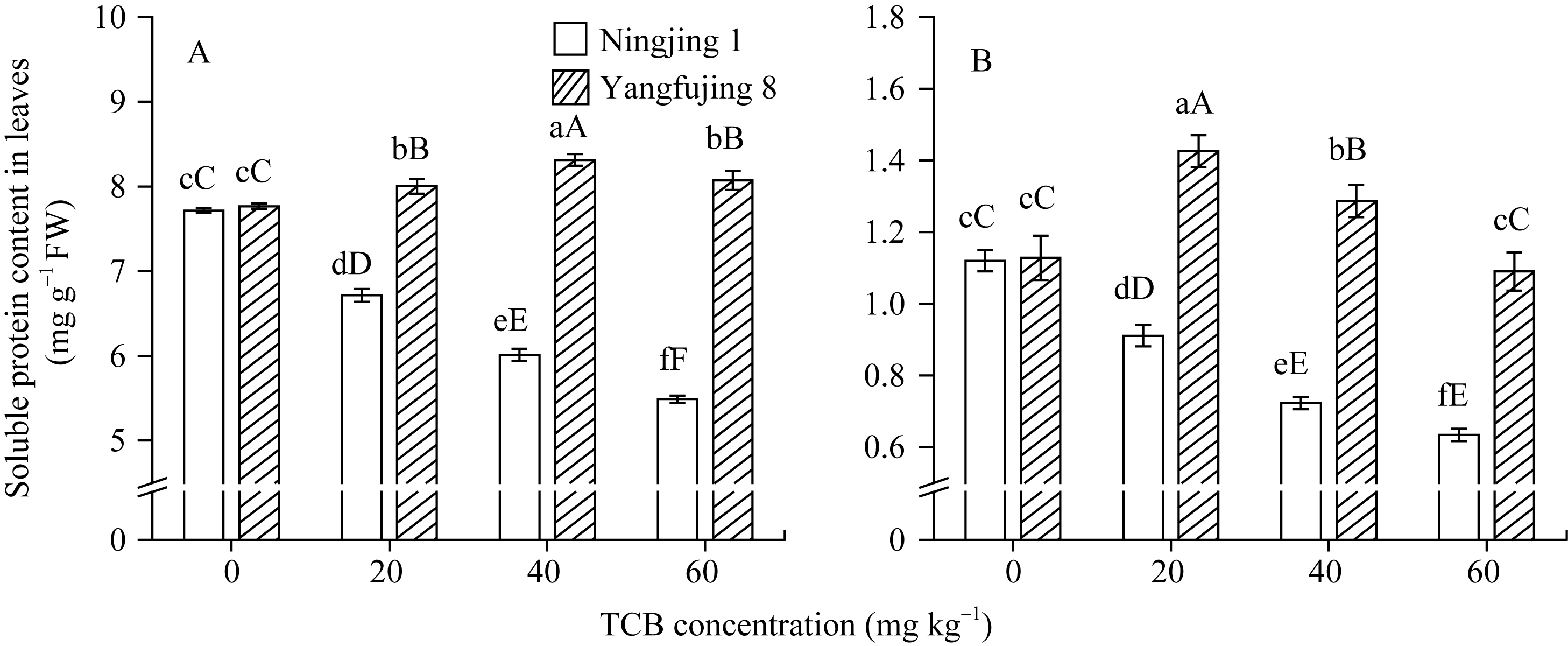 | 图3 TCB胁迫对分蘖盛期水稻叶片(A)和根系(B)可溶性蛋白质含量的影响柱上标以不同大、小写字母时分别表示0.01和0.05差异显著水平。Fig. 3 Effect of TCB on the soluble protein content in rice leaves (A) and roots (B) at top tillering stageBars superscripted by different letters are significantly different at 0.01 (capital) and 0.05 (lowercase) probability levels, respectively. |
2.5.2 对POD活性的影响 TCB胁迫下, 宁粳1号叶片(图4-C)和根系(图4-D) POD活性均极显著低于对照, 且随TCB浓度增加而递减。扬辐粳8号叶片POD活性在中低浓度胁迫下, 均显著高于对照, 且20 mg kg-1与40 mg kg-1间差异不显著; 而根系POD活性在中低浓度TCB胁迫下均极显著高于对照, 从20 mg kg-1开始呈下降趋势, 60 mg kg-1胁迫时与对照差异不显著。
2.5.3 对CAT活性的影响 TCB胁迫下, 宁粳1号叶片(图4-E)和根系(图4-F) CAT活性均极显著低于对照, 且随TCB浓度增加而先递减后略有所上升(40~60 mg kg-1)。扬辐粳8号叶片CAT活性随TCB浓度增加先升高后降低, 在中低浓度TCB胁迫下均极显著高于对照, 而60 mg kg-1时与对照差异不显著; 根系CAT活性在中低浓度TCB胁迫下显著增加, 并呈上升趋势, 高浓度TCB胁迫下CAT活性下降, 与对照相比均达到极显著水平。
2.6 TCB胁迫对2种基因型水稻分蘖盛期 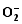
TCB胁迫下, 宁粳1号叶片(图5-A)和根系(图5-B) 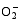
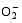
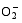
2.7 TCB胁迫对两种基因型水稻分蘖盛期MDA含量的影响
TCB胁迫下, 宁粳1号叶片(图5-C)和根系(图5-D) MDA含量均随TCB浓度增加而极显著升高。扬辐粳8号叶片和根系MDA含量在20 mg kg-1时均极显著低于对照, 而后呈上升趋势, 并显著或极显著高于对照。
3 讨论
随着食品安全问题被屡屡曝光, 人们对农产品质量与安全的关注度日益增加。而CBs可通过多种途径进入农田, 被作物吸收后, 在体内积累、迁移、转化或释放[ 33, 34, 35]。人类长期接触或食用后在体内富集, 进而危害身体健康。水稻、小麦、玉米等单子叶植物的生长发育均易受CBs的影响, 其中水稻的报道较多。
根系是作物吸收水分和养分的主要器官, 也是作物接触土壤中污染物的关键部位之一。以往研究认为TCB胁迫使水稻幼苗的根系伸长受到抑制, 细胞异常如细胞间隙加大、细胞壁显著加厚等, 根系活力也有所降低[ 5, 24, 36]。本研究结果显示, 分蘖盛期水稻的根系对TCB浓度的响应存在基因型差异。低浓度TCB胁迫下, 敏感基因型宁粳1号的最长根长、地下部干物重、根系活力分别比对照降低9.1%、9.3%、2.0%; 而扬辐粳8号分别增加3.2%、1.7%、10.0%。在中高浓度TCB胁迫下, 2种水稻的最长 根长、地下部干物重均极显著降低, 根粗均极显著增加, 但宁粳1号的降(增)幅均大于扬辐粳, 说明中高浓度TCB对根系的毒害效应严重, 即使是耐性基因型扬辐粳8号的地下部生长也受到阻碍, 可能的原因是TCB浓度过高时, 其较高的疏水性导致其在植株根系表面大量吸附, 破坏了细胞器结构及根系稳态; 但由于耐性基因型根系抗氧化酶、谷胱甘肽- S-转移酶和顺式还原酮加双氧酶等解毒酶[ 36]受到
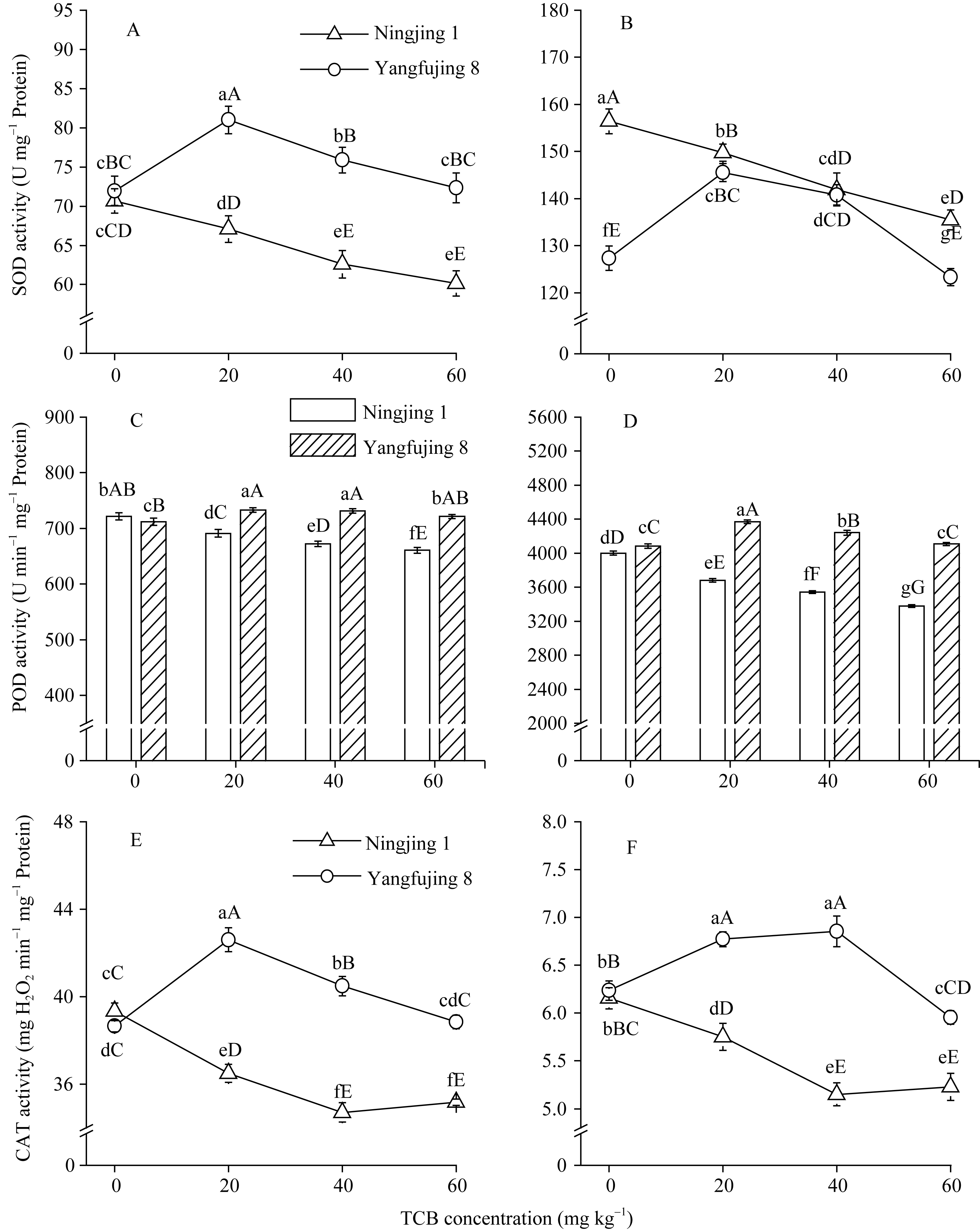 | 图4 TCB胁迫对分蘖盛期水稻叶片(A, C, E)和根系(B, C, D)抗氧化酶活性的影响柱或曲线上标以不同大、小写字母时分别表示0.01和0.05差异显著水平。Fig. 4 Effect of TCB on the activity of antioxidant enzymes in rice leaves (A, C, E) and roots (B, C, D) at top tillering stageBars or curves superscripted by different letters are significantly different at 0.01 (capital) and 0.05 (lowercase) probability levels, respectively. |
Enzymes activities in the curve with different letters are significantly different at P<0.01 (capital) and P<0.05 (lowercase), respectively.
诱导表达, 导致其受到的毒害作用小于宁粳1号。
TCB对水稻苗期株高[ 24]、地上部生物量[ 24]和成熟期有效分蘖数[ 37]抑制作用显著, 本研究显示TCB对分蘖盛期水稻的地上部也有明显的影响。敏感基因型宁粳1号的株高、地上部干物重、单穴分蘖数在TCB胁迫下均显著低于对照, 而耐性基因型扬辐粳8号在低浓度TCB胁迫下, 株高、单穴分蘖数、地上部干物重与对照相比, 增幅分别为0.6%、8.6%和0.6%, 说明扬辐粳8号对低浓度的TCB胁迫有较强的耐受性, 但当TCB浓度过高时, 对水稻植株的生物毒性加大, 影响了细胞的分裂与生长, 对营养物质的吸收和利用产生干扰, 从而使植株的长势长相异常。
光合作用是绿色植物生长发育的物质基础和能量来源。低温、干旱、盐胁迫、重金属等逆境条件均能影响作物的光合作用, 最终导致减产歉收。王泽港等[ 24, 26]研究发现TCB的添加会导致水稻
当大、小写字母不同时分别表示0.01和0.05差异显著水平。
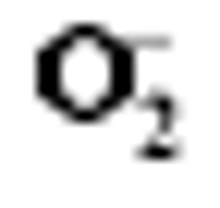
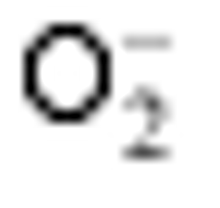
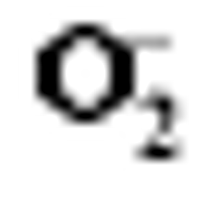
Values of A, B, C, and D with different letters are significantly different at P<0.01 (capital) and P<0.05 (lowercase), respectively.
幼苗期叶绿素含量、Rubisco酶活性的下降及抽穗期水稻叶片叶绿素含量、净光合速率( Pn)、最大光能转换效率( Fv/ Fm)、光合电子传递的量子化效率( ΦPSII)、光系统PSII的潜在活性( Fv/ Fo)、光化学猝灭系数( qP)及非光化学猝灭系数( qN)呈递减趋势; 杜青平、张波等[ 5, 38]发现水稻和小麦幼苗叶片中叶绿素a、b和叶绿素总含量均随TCB浓度的增加而呈降低趋势。本研究表明, 低浓度TCB使分蘖盛期宁粳1号叶 绿素含量比对照降低3.4%, 而对扬辐粳8号却增加0.2%。中高浓度的TCB胁迫使水稻光合器官受到伤害[ 26], 对两品种分蘖盛期的叶绿素含量均有极显著的抑制作用, 从而引起光合作用减弱, 干物重降低。
蛋白质是植物生长发育、新陈代谢过程中的重要物质。已有研究表明TCB能诱导水稻和小麦幼苗蛋白质的表达[ 23, 39], 主要包括解毒酶、细胞壁物质、激素、原初或次生代谢与调节等的相关酶, 进而导致植株体内蛋白质含量的增加[ 25]。本研究显示, 不同浓度TCB胁迫下, 分蘖盛期宁粳1号叶片和根系可溶性蛋白质分别与各自对照相比, 叶片的降幅(13.0%、22.1%、28.8%)远小于根系(18.7%、35.4%、43.4%); 扬辐粳8号叶片可溶性蛋白质的表达受到了诱导, 增幅分别为3.1%、7.1%、3.9%。中低浓度TCB胁迫下, 扬辐粳8号根系可溶性蛋白质含量的增幅分别为14.1%、26.4%; 而高浓度胁迫时, 根系可溶性蛋白质含量与对照相比差异不显著。说明扬辐粳8号对TCB胁迫的耐性强, 中低浓度能够充分诱导适应逆境相关蛋白及酶的表达, 而宁粳1号耐性较差, 蛋白合成受阻或者分解加快, 导致蛋白质含量较低, 对有机污染胁迫适应性差。
超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等是植物体内清除活性氧的重要抗氧化酶, 也是酶保护系统的重要组分。酶保护系统启动后, 能及时清除一定数量过剩的活性氧和过氧化物, 从而减轻过氧化作用的伤害, 提高细胞的抗逆性。丙二醛(MDA)含量反映了细胞膜脂过氧化程度及器官的衰老情况。高浓度TCB胁迫下, 水稻敏感品种幼苗的MDA含量显著增加, SOD、POD、CAT活性有所降低, 根系受到的伤害明显大于叶片[ 5, 25]。张国良等[ 22]认为小麦敏感品种根系的SOD、POD活性在低浓度TCB胁迫下有所升高, 而CAT活性却显著降低。刘宛等[ 20, 21]发现TCB能够使萌发的大豆种子活性氧代谢紊乱, 超氧阴离子自由基( 
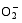
低浓度TCB胁迫对扬辐粳8号的生长发育有明显的促进作用, 可能主要有以下三方面原因。(1)植株的解毒系统适应性强, 抗氧化酶活性有所提高、谷胱甘肽-S-转移酶和顺式还原酮加双氧酶(ARD)受到诱导表达[ 36], 有利于清除活性氧和有毒物质、维持细胞结构的稳定, 进而增强植株的抗逆能力; (2) MicroRNA是一类与植物适应逆境胁迫有关的调控基因表达的小分子RNA[ 40], CB等有机污染胁迫可能诱导或下调水稻相关MicroRNA的表达, 从而调节植株的代谢过程; (3) 低浓度TCB能增加以其为碳源的微生物数量, 提高土壤肥力[ 41, 42], 而扬辐粳8号对TCB胁迫有一定耐受限度, 在低浓度TCB下仍能正常吸收与利用养分。
4 结论
不同浓度TCB胁迫下敏感基因型和耐性基因型水稻的生长和生理特性差异显著, 敏感基因型随胁迫浓度的增加生长和生理活性受抑程度显著递增; 而耐性基因型在低浓度TCB胁迫下, 植株解毒能力
较强, 最长根长、地上部和地下部干物重均极显著高于对照, 根系活力、叶片和根系可溶性蛋白含量较高, 酶保护系统清除活性氧的能力较强, 膜脂过氧化程度较低。中高浓度TCB胁迫才对耐性基因型水稻的生长表现出明显的抑制作用。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|