{kind=link}
{kind=link}
{kind=link}
{kind=link}
水稻第1染色体长臂上微效千粒重QTLqTGW1.2 的验证与分解
[陈玉宇1, 2 , 朱玉君1 , 张宏伟1 , 王琳琳1 , 樊叶杨1 , 庄杰云1, *  ]
]
]
|
|


本文报道了水稻第1染色体长臂上微效千粒重QTL
Validation and dissection of a minor QTL
粒重直接影响水稻产量, 属于受多基因控制的数量性状, 是水稻数量性状座位(QTL)研究领域进展最快的性状之一, 定位的千粒重QTL分布于水稻的所有12条染色体[ 1], 并已有7个得到克隆, 其中, GS3[ 2]、 GL3.1 ( qGL3)[ 3, 4]和 TGW6[ 5]主要控制粒重和粒长, GW2[ 6]、 qSW5 ( GW5)[ 7, 8]、 GS5[ 9]和 GW8[ 10]主要控制粒重和粒宽。这些已克隆的千粒重QTL具效应大、贡献率高、能稳定检测到等特点, 作用最强的 qGL3加性效应达5.57 g[ 4], 初定位时在2个不同群体中均被检测到, 对表型方差的贡献率分别达23.09%和27.99%[ 4, 11]; 作用最弱的 GS5加性效应为0.8 g[ 9], 初定位时在3个不同群体被检测到, 贡献率为12.30%’24.10%[ 9, 12, 13]。从另一方面看, 效应较小的QTL易受环境和遗传背景的干扰, 其研究仍比较困难, 至今未见微效千粒重QTL的精细定位和克隆报道。
水稻抽穗期是决定水稻品种区域和季节适应性的首要农艺性状。前人研究表明, 部分千粒重QTL对抽穗期亦呈显著效应且效应方向一致, 例如, Song等[ 6]利用FAZ1/WY3组合衍生的BC3F2群体将 GW2精细定位于只含一个开放阅读框的8.2 kb的区域内, 来自WY3的等位基因提高千粒重, 同时延迟抽穗; Xie等[ 14]利用IRGC105491/Hwaseongbyeo组合衍生的BC3F4群体将 gw9.1精细定位于37.4 kb的区域内, 来自IRGC105491的等位基因提高千粒重, 同时延迟抽穗。在水稻粒重的改良中, 所利用的千粒重QTL如果与抽穗期存在不利关联, 则千粒重的提高将伴随着抽穗的延迟, 需要考虑水稻品种区域和季节的适应性; 所利用的千粒重QTL如果对抽穗期无显著作用, 则可避免因抽穗期变异产生的不利影响。
在前期研究中, 我们应用珍汕973/密阳46衍生的高代群体, 在水稻第1染色体长臂上检测到2个千粒重QTL[ 15]。本研究针对其中的 qTGW1.2, 先应用BC2F8:9近等基因系验证其效应, 再构建在目标区间内分离区间呈交迭排列的5个BC2F10群体, 将 qTGW1.2分解为互引连锁的2个不同QTL。
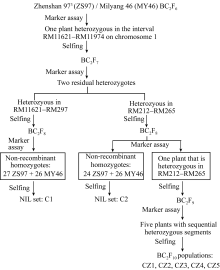
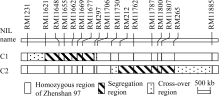
包括2套BC2F8:9近等基因系和5个BC2F10分离群体, 其构建流程(图1)如下。应用前期千粒重QTL分析[ 15]的珍汕973/密阳46 BC2F6群体, 经SSR标记鉴定, 筛选出在第1染色体长臂RM11621-RM11974区间呈杂合的单株1个, 自交产生BC2F7群体。经目标区间SSR标记检测, 从BC2F7群体中筛选出分别在RM11621-RM297和RM212-RM265区间呈杂合的2个单株, 自交收获种子。种植2个BC2F8群体各250个单株, 经分离区间标记检测, 筛选母本纯合型(即珍汕97纯合型, 下简称珍汕型)和父本纯合型(即密阳46纯合型, 下简称密阳型)材料; 自交收获种子, 产生2套BC2F8:9近等基因系, 其中, C1含珍汕型株系27个和密阳型株系26个, C2含珍汕型株系24个和密阳型株系26个。这2套近等基因系在目标区间的基因型组成如图2所示。
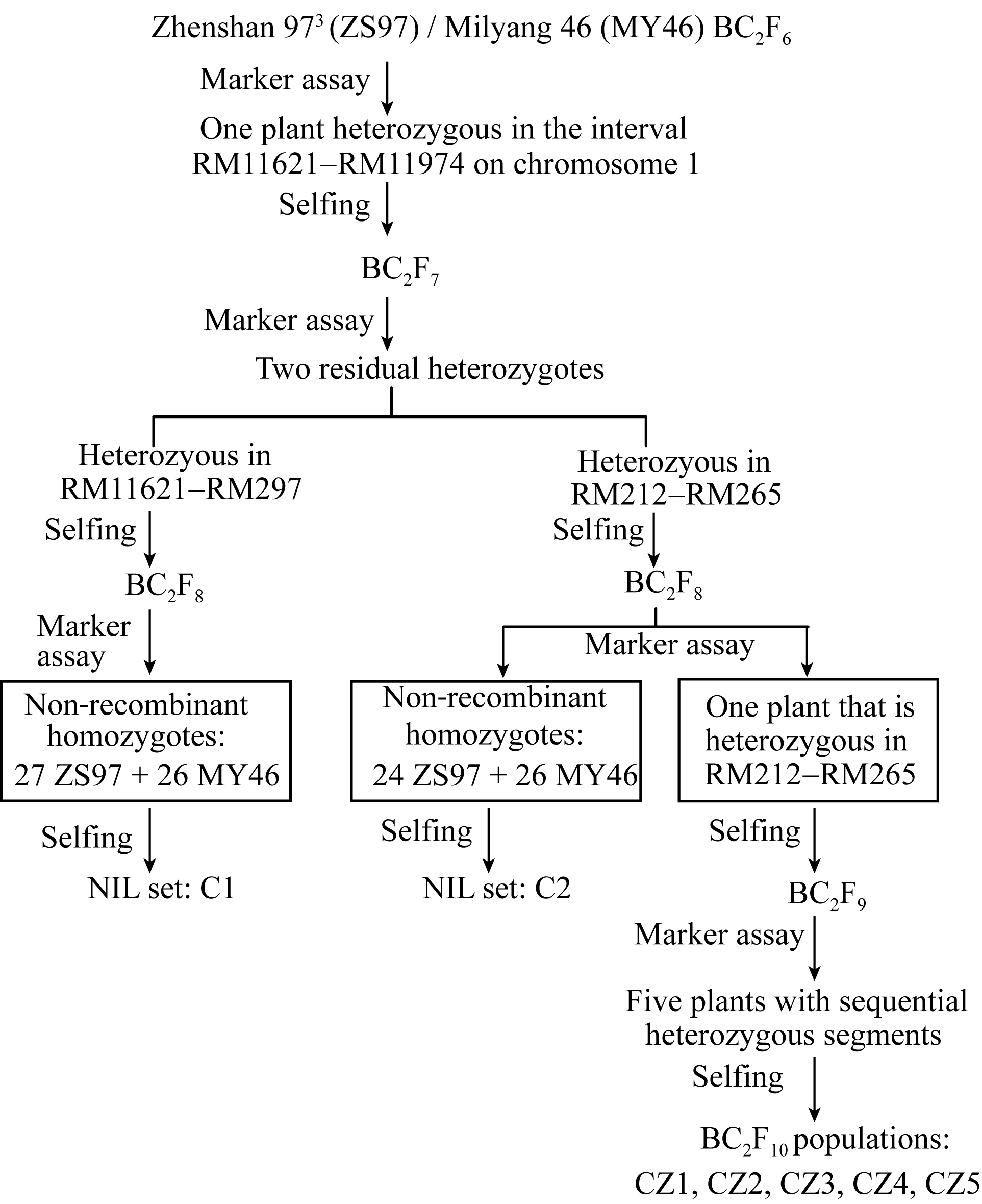 | 图1 材料构建流程图Fig. 1 Development of the rice populations used in this study |
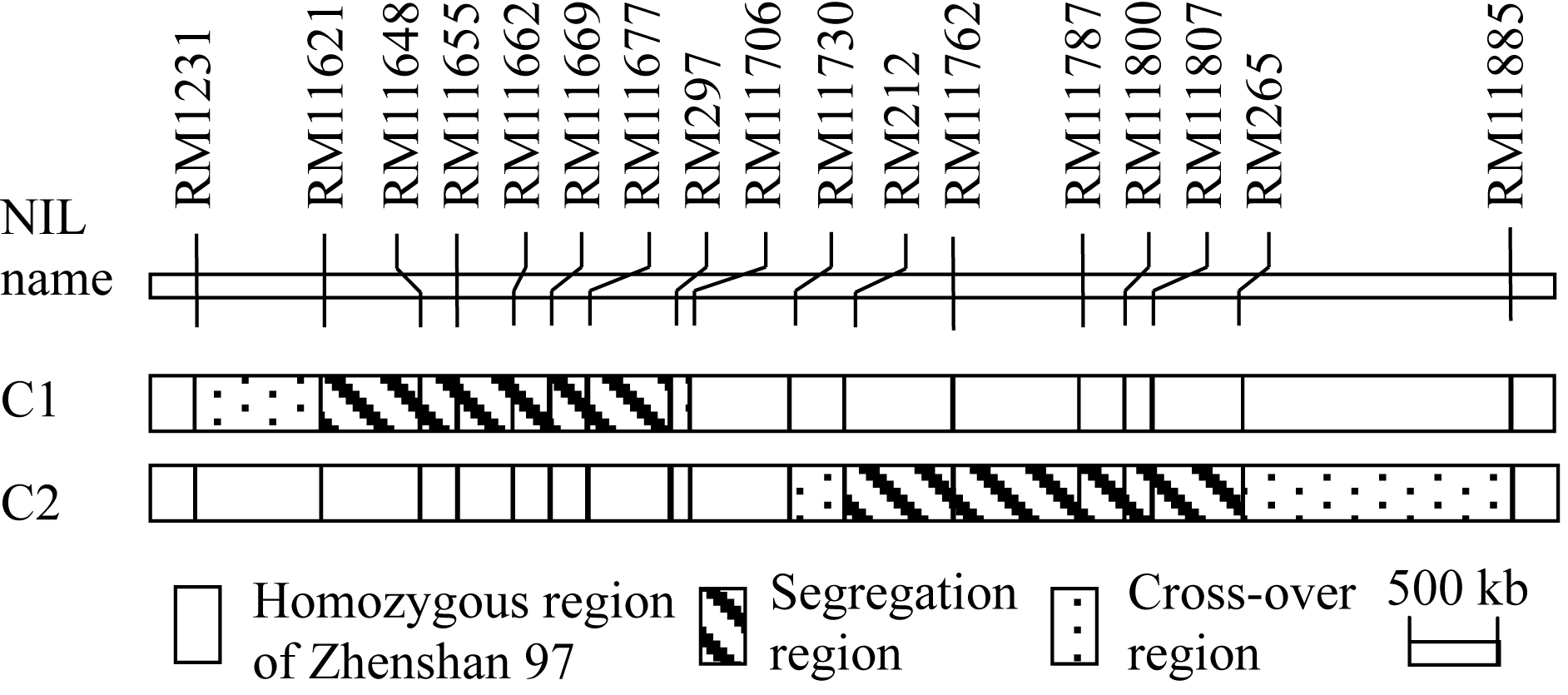 | 图2 2套近等基因系在区间RM1231-RM11885的基因型Fig. 2 Genotypes in the interval RM1231-RM11885 of two sets of near isogenic lines (NILs) |
同时, 从上述2个BC2F8群体中各挑选出在整个分离区间呈杂合的单株1个, 自交收获种子, 获得2个BC2F9群体。根据近等基因系C1和C2的QTL分析结果, 种植分离区间为RM212-RM265的BC2F9群体, 经SSR标记检测, 筛选出在目标区间内杂合区段交迭排列的5个单株, 自交收获种子, 获
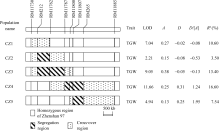
得5个BC2F10群体, 分别称之为CZ1、CZ2、CZ3、CZ4和CZ5, 它们在目标区间的基因型组成如图3所示。
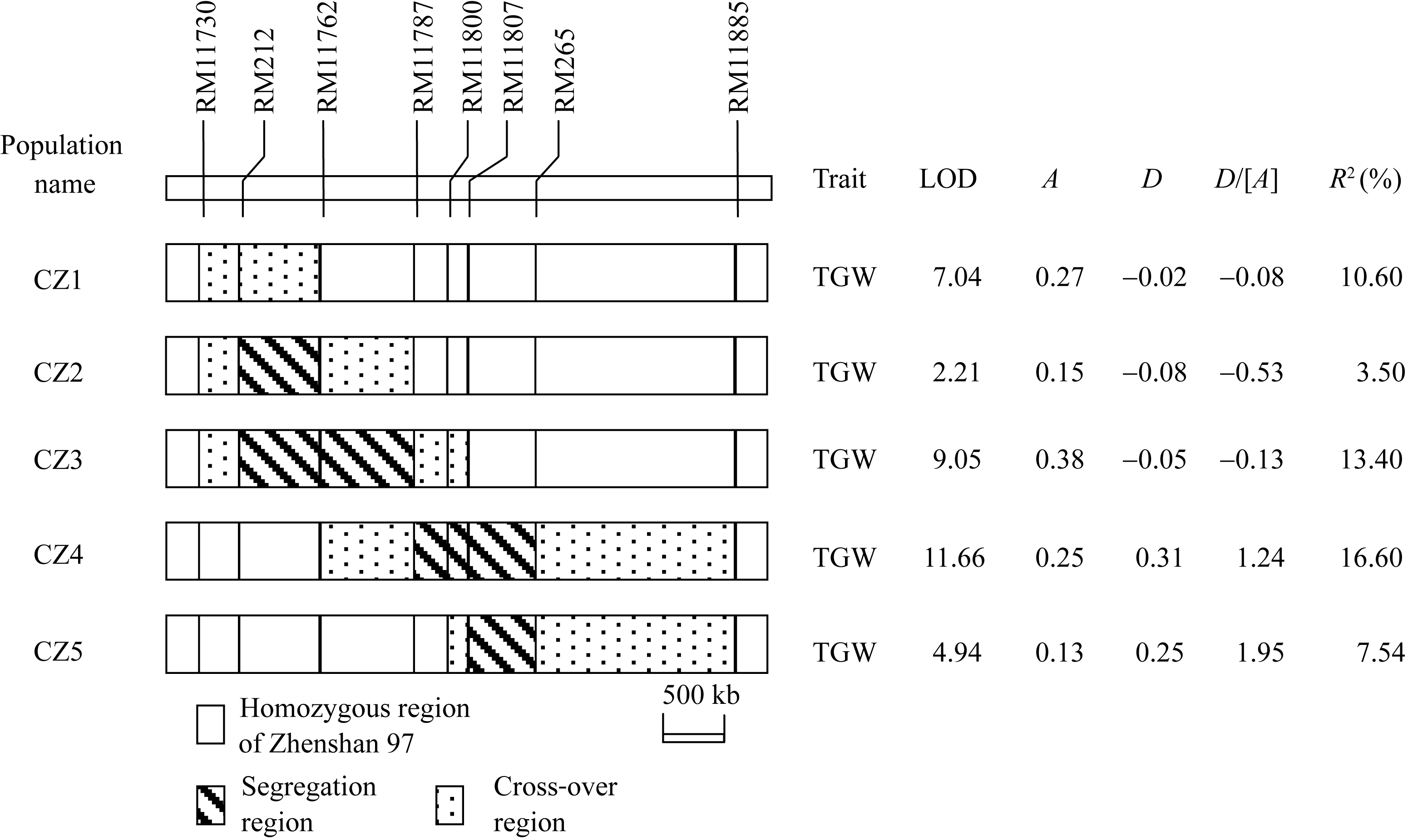 | 图3 应用5套NIL-F2群体对千粒重的QTL检测结果TGW: 千粒重; A: 加性效应, 指密阳46等位基因取代珍汕97等位基因所产生的遗传效应; D: 显性效应; D/[ A]: 显性度; R2: QTL效应对表型方差的贡献率。Fig. 3 QTL analysis for 1000-grain weight using five NIL-F2 populationsTGW: 1000-grain weight; A: additive effect measured as the genetic effect when a Zhenshan 97 allele is replaced by a Milyang 46 allele; D: dominance effect; D/[ A]: degree of dominance; R2: proportion of phenotypic variance explained by the QTL effect. |
2012年5月至9月在浙江富阳种植2套近等基因系C1和C2, 前者含珍汕型株系27个和密阳型株系26个, 后者含珍汕型株系24个和密阳型株系26个; 2个重复, 每重复每株系种植1行8个单株, 株行距16.7 cm × 26.7 cm, 完全随机区组设计, 正常田间管理。记载各单株抽穗期, 以8株平均值为基础进行数据分析; 成熟后每行取中间5株混收, 晒干后取出结实的稻谷, 称量后换算成单株产量, 并挑选出2份300粒的饱满稻谷, 采用SC-A型种子数粒仪(万深)测定千粒重、粒长和粒宽。
2012年12月至2013年4月在海南陵水种植CZ1、CZ2、CZ3、CZ4和CZ5, 每个群体300个单株, 株行距16.7 cm × 26.7 cm, 正常田间管理。记载各单株抽穗期, 成熟后分单株收获种子, 晒干后挑选出2份100粒饱满的稻谷, 称量后换算成千粒重。
在上述群体中, 有8个进行了SSR标记检测, 包括分离区间分别为RM11621-RM297和RM212- RM265的2个BC2F8群体、分离区间为RM212- RM265的1个BC2F9群体, 以及在RM212-RM265区间内部分区段分离的5个BC2F10。于各群体移栽后 7 d, 从每个单株取叶片约2 cm, 采用微量法[ 16]提取DNA。采用各群体的分离标记分别检测, 其中, CZ1群体为RM212, CZ2群体为RM212和RM11762, CZ3群体为RM212、RM11762和RM11787, CZ4群体为RM11787、RM11800、RM11807和RM265, CZ5群体为RM11807和RM265。SSR标记信息源自Gramene数据库(http://www.gramene.org/), PCR扩增产物和产物检测遵循以前所用方法[ 15]。
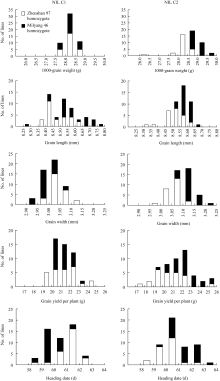
如图4所示, 在千粒重、粒长、粒宽、单株产量和抽穗期这5个性状上, 2套近等基因系各株系的表型平均值均呈连续分布, 说明同一套近等基因系的不同株系之间不存在主效基因的分离; 比较同一套近等基因系内2种基因型的表型分布, 则可发现, 2套群体在粒长、粒宽、单株产量和抽穗期这4个性状上差异较小, 但在千粒重上存在明显差异, 即C1的珍汕型和密阳型株系在众数和分布范围上均高度一致, 而C2的珍汕型株系主要分布于低值区、密阳型株系主要分布于高值区。
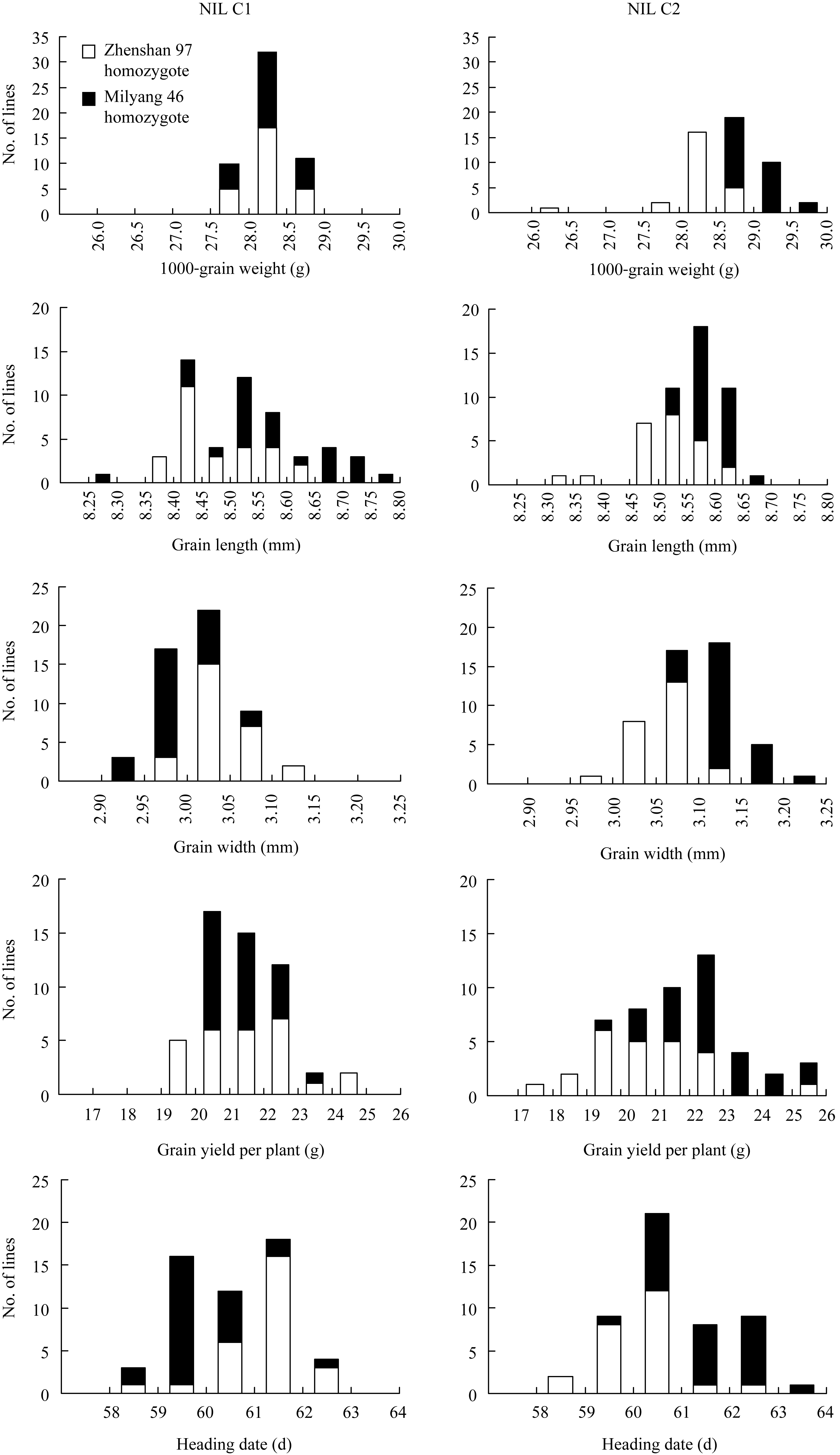 | 图4 2套近等基因系的表型分布Fig. 4 Phenotype distributions of the two NIL sets |
从这些结果可以推测, 近等基因系C2的不同基因型材料之间可能存在控制水稻千粒重的等位基因差异, 而近等基因系C1的不同基因型材料之间不存在控制水稻千粒重的等位基因差异。
2套近等基因系千粒重和单株产量的方差分析结果显示(表1), C2的2种基因型材料之间在两性状上均呈极显著差异( P<0.0001), 而C1在两性状上都未呈显著差异。进一步分析C2群体中2种基因型材料在粒长和粒宽上的差异表明, 其珍汕型和密阳型材料在两粒形性状上均极显著差异( P<0.0001)。由于RM212- RM265在C2群体呈分离、在C1群体呈珍汕型(图2), 故可将 qTGW1.2界定在RM212-RM265及其两侧交换区间的区域内。该区间同时控制千粒重、粒长、粒宽和单株产量, 其增效等位基因均来自于密阳46, 加性效应分别为0.402 g、0.036 mm、0.036 mm和0.850 g, 贡献率分别为45.21%、24.13%、48.66%和13.12%。
| 表1 2套近等基因系中基因型变异对千粒重、粒长、粒宽、单株产量和抽穗期的作用 Table 1 Genotypic effects on 1000-grain weight, grain length, grain width, grain yield, and heading date in the two NIL sets |
在抽穗期性状上, 2套近等基因系均检测到显著变异, 加性效应分别为0.657 d和0.696 d, 贡献率分别为24.52%和26.50%, 其中, 促进抽穗期的等位基因在C1群体来自密阳46、在C2群体来自珍汕97。
以5套BC2F10群体对千粒重QTL检测显示(图3), 每一套群体均检测到千粒重QTL, 且加性效应方向一致, 增效等位基因均来源于密阳46。在5套群体中, CZ1的分离区间(含交换区间, 下同)位于RM11730和RM11762界定的区域内, CZ5的分离区间位于RM11800和RM11885界定的区域内, 两者互不交迭, 故可判断, 这2个区间分别存在1个控制千粒重的QTL, 后文称其为 qTGW1.2a和 qTGW1.2b。
在其他群体中, CZ2和CZ4的分离区间分别覆盖 qTGW1.2a和 qTGW1.2b所处区间, 且不与另一个QTL区间交迭, 说明这2个群体的千粒重变异分别由 qTGW1.2a和 qTGW1.2b控制; 从遗传作用模式看, 在CZ1和CZ2群体中分离的千粒重QTL显性效应小、表现为加性作用, 在CZ4和CZ5群体中分离的千粒重QTL显性效应大、表现为正向超显性, 进一步说明CZ1和CZ2同受 qTGW1.2a控制, 而CZ4和CZ5同受 qTGW1.2b控制。在CZ3群体中, 其分离区间覆盖CZ1的分离区间, 同时只与CZ5的部分交换区间重迭; 另外, 从遗传作用模式看, CZ1和CZ3群体表现为加性作用, 而CZ5群体表现为正向超显性, 说明CZ3群体的千粒重变异由 qTGW1.2a控制。
综上所述, CZ1、CZ2和CZ3群体的千粒重变异均由 qTGW1.2a控制, 可将 qTGW1.2a界定在这3个群体的共有分离区间内, 即RM11730和RM11762之间934 kb的区域内。该QTL呈加性作用, 增效等位基因来源于密阳46, 在CZ1、CZ2和CZ3中加性效应分别表现为0.27、0.15和0.38 g, 贡献率分别为10.60%、3.50%和13.40%。CZ4和CZ5群体的千粒重变异均由 qTGW1.2b控制, 可将 qTGW1.2b界定在这2个群体的共有分离区间内, 即RM11800和RM11885之间2.1 Mb的区域内。该QTL呈正向的超显性, 增效等位基因亦来源于密阳46, 在CZ4和CZ5中加性效应分别表现为0.25 g和0.13 g, 贡献率分别为16.60%和7.54%。
在抽穗期性状上, 5个BC2F10群体中均未检测到显著的QTL效应, LOD值变幅为0~1.21, 而在采用BC2F8:9近等基因系的前一个试验中, 这5个群体覆盖的RM212-RM265分离区间呈现出对抽穗期的显著作用(表1)。鉴于BC2F8:9近等基因系中检测到的抽穗期QTL作用很小, 加性效应只有0.7 d, 上述冲突既可能源于近等基因系中检测到的作用属假阳性, 也可能由于BC2F10群体的研究基于单株、基因型内变异较大, qTGW1. 2对抽穗期的作用难以达到显著水平; 此外, 2个试验分别在浙江长日照和海南短日照下进行, 也可能由于该QTL在海南短日条件下不表现。
在QTL初定位研究中, 由于受到群体规模的限制和复杂遗传背景的影响, 其定位精度较低, 一般超出10 cM[ 20, 21]。针对初定位检测到的QTL区域, 构建目标区间分离、背景基本一致的材料进一步验证和分解, 往往把一个区间较大的QTL分解为精度较高的不同QTL[ 22, 23]。在前期研究中[ 15], 我们应用衍生于珍汕973/密阳46高代群体的次级定位群体, 在水稻第1染色体长臂上1个约12 Mb区间中分解出互斥连锁的2个微效千粒重QTL, 其效应可在不同世代和不同地区被稳定检测到, 其中 qTGW1.2位于RM11615和RM11800之间4.5 Mb的区域内, 其加性效应为0.41’0.62 g, 增效等位基因来源于密阳46。本研究进一步构建背景同质性更高、分离区间更小的群体, 验证了 qTGW1.2的效应, 并将其分解为紧密连锁的2个QTL, 其增效等位基因均来自密阳46, 加性效应方向一致, 为互引连锁; 其中, qTGW1.2a位于RM11730和RM11762之间934 kb的区域内, 加性效应为0.15’0.38 g, 遗传作用模式为加性; qTGW1.2b位于RM11800和RM11885之间2.1 Mb的区域内, 加性效应为0.13’0.25 g, 遗传作用模式为正向超显性。这些结果表明, 虽然微效QTL定位易受环境和遗传背景影响, 但在遗传背景高度纯合的情况下, 其效应能被稳定地检测到; 因此, 通过遗传背景的高度同质化, 可对效应较低的QTL进行验证和分解, 甚至于进一步精细定位和克隆。
在水稻分子设计育种中, 对抽穗期的作用是决定QTL应用潜力的关键因素之一。利用延迟抽穗、提高粒重的QTL会使水稻的生育期延长, 从而影响其在特定生态区域的种植, 而且, 生育期延长会影响不同季节作物的轮种, 而与抽穗期无不利关联的QTL将更适合于在育种中应用。在我们前期的研究中, qTGW1.2区域在不同年份、不同地点及不同遗传背景的群体均没有表现出对抽穗期的作用[ 15, 24]。本试验继续分析 qTGW1.2是否存在对抽穗期的作用, 在以单株鉴定为基础的NIL-F2群体中, qTGW1.2对抽穗期未呈显著作用, 而在以近等基因系为材料的随机区组试验中, qTGW1.2表现出对抽穗期的显著作用, 且效应方向与其对千粒重的作用一致。不过, 在检测到抽穗期QTL的近等基因系材料中, 其加性效应很小, 仅仅只有0.7 d, 可见 qTGW1.2在育种中应用可以在不明显延长生育期的前提下改良水稻粒重。
水稻千粒重的遗传力较高, 受环境的影响较小, 可在育种材料的早期世代选择[ 25]; 在现代水稻育种中, 粒重的增加对水稻产量的提高具有重要的作用[ 26, 27]。龚金龙等[ 28]对大穗型杂交粳稻产量构成因素协同特征研究的结果表明, 在满足一定穗数和具备稳定结实率的基础上, 稳定和提高千粒重才能有效发挥大穗型品种的增产潜力。本研究所选用水稻组合的亲本珍汕97和密阳46分别为中国大面积推广三系杂交稻组合汕优10号的保持系和恢复系, 所检测到的QTL是研究水稻骨干亲本重要等位基因变异的重要候选对象。从本研究结果可知, qTGW1.2对千粒重和单株产量的效应方向一致, 表明该粒重QTL对水稻的单株产量具有显著作用, 可应用于水稻高产育种; 在 qTGW1.2区间分解出的2个QTL中, qTGW1.2b呈正向超显性, 可能对汕优10号F1代杂种优势具有重要贡献, 这对进一步研究水稻杂种优势分子遗传机理具有重要意义。
Liu等[ 29]应用特青/珍汕97的190个F7重组自交系在第1染色体长臂上RM297-RM319区域检测到1个微效千粒重QTL, 覆盖了 qTGW1.2a所在的区间。说明 qTGW1.2a在不同组合中对千粒重发挥着显著作用。迄今为止, 在 qTGW1.2a和 qTGW1.2b区间均未见粒重QTL精细定位和克隆的报道; 目前, 我们已构建了分离区间进一步缩小的群体, 以开展这2个千粒重QTL的精细定位。
应用2套珍汕973/密阳46 BC2F8:9近等基因系, 对水稻第1染色体长臂上微效千粒重QTL qTGW1.2进行了遗传学验证, 并将其进一步界定在RM212- RM265及其两侧交换区间的4.5 Mb区域内。进而应用5个珍汕973/密阳46 BC2F10群体, 将 qTGW1.2分解为互引连锁的2个QTL, 其中, qTGW1.2a位于RM11730和RM11762之间934 kb的区域内, 呈加性作用; qTGW1.2b位于RM11800和RM11885之间2.1 Mb的区域内, 呈正向超显性。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|