


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
超级稻根系形态生理特征及其与产量形成的关系
[褚光 , 刘洁, 张耗, 杨建昌
, 刘洁, 张耗, 杨建昌* ]
, 刘洁, 张耗, 杨建昌]
|
|


* 通讯作者(Corresponding author): 杨建昌, E-mail:jcyang@yzu.edu.cn, Tel: 0514-87979317
第一作者联系方式: E-mail:chuguang19880210@163.com
2011年与2012年于水培池种植超级稻品种两优培九(两系杂交籼稻)和扬粳4038 (粳稻)及常规高产品种扬稻6号(籼稻)和扬辐粳8号(粳稻), 观察各生育期根系形态生理特征及其与产量形成的关系。结果表明, 与常规高产品种相比, 超级稻品种具有强大的产量潜力, 其高产潜力主要得益于较多的每穗颖花数与较大的总库容量(单位地面积颖花量)。超级稻品种在整个生育期较大的根量、生育早中期较高的单株根系氧化力、根系总吸收表面积与活跃吸收表面积及根系中较高的玉米素与玉米素核苷含量是其产量库容大和产量高的重要原因。超级稻存在着结实率较低的问题, 这可能与其在灌浆期根系活性下降较快有关。提高超级稻灌浆期根系活性, 是提高其结实率、促进其产量进一步提高的重要途径。
Two super rice cultivars, Liangyoupeijiu (
水稻是世界上最重要的粮食作物之一, 约为半数以上人口提供了饮食能量[ 1]。在我国, 以水稻为主食的人口约占45%。预测到2030年我国人口将达16亿, 届时即使耕地面积不减少, 粮食产量仍需提高55%才能满足人们对食品的需求。在新增粮食产量中, 水稻所占比率将高达40%以上[ 2]。为了增强我国粮食科技储备能力, 促成水稻单产的再次飞跃, 农业部于1996年开始组织实施“中国超级稻育种与栽培体系研究”重大科技项目[ 3], 目前已育成多个超级稻品种并在生产上推广应用, 取得了显著的增产效果, 为我国水稻生产的发展作出了重要贡献。但是生产实践证明, 部分超级稻品种在年度间和地块间存在着结实率低和产量不稳定的问题[ 4, 5, 6, 7, 8]。近年来, 一些研究者从叶片光合性能、物质生产与运转、源库关系等方面研究超级稻产量形成的特点[ 9, 10], 但超级稻结实率较低的机制仍不清楚。
根系作为水稻植株的重要组成部分, 不仅能够吸收养分和水分, 同时也可合成许多重要的生理活性物质, 因而在水稻生长发育中起着十分重要的作用[ 11, 12, 13]。水稻根系形态生理特征是根系质量的体现, 与地上部的生长发育、养分吸收、产量形成等关系非常密切。但有关超级稻根系形态生理特征及其与产量形成特别是与结实率关系的研究较少。本研究观察了超级稻整个生育期根系形态生理特征的变化, 包括根系生物量、根长、根直径、根系氧化力、根系总吸收表面积与根系活跃吸收表面积、根系中玉米素(Z)与玉米素核苷(ZR)含量、根系伤流液强度及其与产量形成的关系, 以明确超级稻品种根系形态生理特征, 为充分发挥超级稻产量潜力提供理论依据。
试验于2011—2012年在扬州大学农学院试验农场水培池进行。2个超级稻品种为两优培九(两系杂交籼稻)和扬粳4038 (粳稻), 以常规高产品种扬稻6号(籼稻)和扬辐粳8号(粳稻)为对照。5月12日至14日播种于大田, 湿润育秧, 20 d后移栽到水泥池中, 株行距为15 cm × 18 cm, 2011年单本栽两优培九, 双本栽另3个品种; 2012年4个品种均为双本栽。水培液按国际水稻研究所标准Espino营养液配方, 移栽时用海绵固定。每天用1 mol L-1 HCl或 1 mol L-1 NaOH调节pH值至5.0, 每周更换一次营养液, 若遇大雨则翌日更换营养液。各处理(品种)小区的面积为6.0 m × 1.5 m, 重复3次。全生育期严格控制病虫害。各品种于8月18日至26日抽穗, 10月7日至14日收获。
1.2.1 叶片形态与生理特性 分别于分蘖中期、穗分化始期、抽穗期、抽穗后10 d、20 d、30 d, 考察每个小区100穴植株的分蘖数, 按照平均茎蘖数取10穴植株, 分解为绿叶、枯叶、茎、鞘和穗(抽穗以后), 烘干测定干物质重。于以上时期从各小区取2个8穴, 采用美国LI-COR公司生产的LI-COR 3050型叶面积仪测定叶面积。并于上述相同时期, 选择晴朗上午, 于9:00采用美国LI-COR公司生产的LI-6400便携式光合测定仪测定稻株最上展开叶的光合速率。叶室CO2浓度为380 μmol mol-1, 使用红蓝光源, 光量子通量密度(PFD)为1400 μmol m-2 s-1, 温度28~30℃, 各处理(品种)重复测定8张叶片。
1.2.2 根系形态与生理特性 分别于上述时期, 从水泥池中取出8穴植株, 剪掉地上部分(用于测定地上部生物量与叶面积), 置70目筛网袋中, 用流水冲洗干净。各取2穴分别测定根长和根直径、根系氧化力、根系吸收表面积和根系激素。根长和根直径的测定方法为, 将根放入装有一薄层去离子水的玻璃皿中(30 cm × 30 cm), 用镊子调整根的位置以尽量避免交叉重叠, 先用扫描仪(Epson Expression 1680 Scanner, Seiko Epson Corp. Tokyo, Japan)进行图像扫描, 后用WinRHIZO根系分析系统(Regent Instruments Inc. Quebec, Canada)分析, 并于烘干后测干重。参照杨建昌等[ 14]的方法测定根系氧化力。采用甲烯蓝蘸根法[ 15]测定根系总吸收表面积与根系活跃吸收表面积。参照陈远平等[ 16]的高效液相色谱法并作改进, 提取、纯化和定量分析根系激素。用石油醚萃取去除样品中的叶绿素和脂肪等物质, 经Sep-Pak C18柱过滤以减少样品中杂质; 色谱条件改用Dubhe C184.6×250, 5 μm, 流动相为5% (v/v)乙腈、50% (v/v)甲醇、0.6% (v/v)冰乙酸, 流速为0.8 mL min-1, 采用梯度洗脱法, 检测波长254 nm; 柱温30℃, 进样量20 μL。样品回收率为85.5%±2.6%, 每一个样品至少重复4次。以外标法定量。
1.2.3 根系伤流强度 分别于抽穗后0 d、14 d、28 d, 按各小区的平均茎蘖数选取代表性植株5穴, 于18:00在各茎离水泥板10 cm处剪去地上部分植株, 将预先称重的脱脂棉置于茎杆的剪口处, 包上塑料薄膜, 于第2天上午6:00取回带有伤流液的脱脂棉并称重, 计算伤流强度。
1.2.4 考种与计产 取成熟期各小区50穴考察每穴穗数, 取10穴观察结实率(水漂法, 沉入水底者为饱粒)和千粒重。实收各小区计产。
采用Microsoft Excel 2003、SPSS 16.0和SAS统计软件分析试验数据, 用SigmaPlot 10.0绘图。
两个超级稻品种2年的产量均显著高于对照品种, 两优培九平均产量为9.86 t hm-2; 扬粳4038为9.56 t hm-2, 分别较对照品种高28.37%和17.63% (表1)。超级稻品种获得高产主要得益于其较高的总颖花数, 两优培九与扬粳4038两年的平均总颖花数分别较对照品种高60.10%和28.43%。但超级稻品种
| 表1 供试水稻品种产量及其构成因素 Table 1 Grain yield and yield components of rice cultivars tested |
两年的结实率却显著低于对照品种, 分别为74.80%和79.40%, 较对照品种低14.4和11.1个百分点。
两个超级稻品种2年抽穗期的总叶面积指数、有效叶面积指数、高效叶面积指数均显著高于各自对照品种, 分别平均高出19.15%、20.41%和29.72%。2个超级稻品种两年抽穗期颖花数/叶面积(粒叶比)显著性高于对照品种; 实粒数/叶面积与对照品种互有高低; 粒重/叶面积则与对照品种无显著差异(表2)。
| 表2 供试水稻品种抽穗期叶面积指数和粒叶比 Table 2 Leaf area index and grain-leaf ratio at heading stage of the tested rice cultivars |
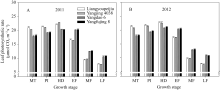
各品种剑叶净光合速率, 在各主要生育期呈现先增后降的趋势, 峰值出现在抽穗期(图1)。分蘖中期至抽穗期两年的剑叶净光合速率, 两优培九高于对照品种13.28%, 扬粳4038高于对照品种10.02%; 而在灌浆期则显著低于对照品种, 分别较对照品种低22.11%和22.87%。
 | 图1 供试水稻品种在不同生育时期叶片光合速率MT: 分蘖中期; PI: 穗分化始期; HD: 抽穗期; EF: 灌浆早期; MF: 灌浆中期; LF: 灌浆后期。Fig. 1 Leaf photosynthetic rate of the tested rice cultivars at different growth stagesMT: mid tillering; PI: panicle initiation; HD: heading period; EF: early grain filling; MF: mid grain filling; LF: late grain filling. |
在分蘖中期和穗分化始期, 4个供试品种地上部干物重无显著性差异; 在抽穗至成熟期, 超级稻品种的地上部干物重则显著高于对照品种(图2-A, B)。
 | 图2 供试水稻品种在不同生育时期地上部干物重(A, B)、根干重(C, D)和根-冠比(E, F)MT: 分蘖中期; PI: 穗分化始期; HD: 抽穗期; EF: 灌浆早期; MF: 灌浆中期; LF: 灌浆后期。Fig. 2 Shoot dry weight (A, B), root dry weight (C, D), and root-shoot ratio (E, F) of the tested rice cultivars at different growth stagesMT: mid tillering; PI: panicle initiation; HD: heading period; EF: early grain filling; MF: mid grain filling; LF: late grain filling. |
在整个生长阶段, 超级稻品种的根干重均显著高于对照品种, 两优培九和扬粳4038两年的平均根干重分别较对照品种高出15.01%和14.19% (图2-C, D)。在分蘖中期和穗分化始期, 超级稻品种的根-冠比显著高于对照品种, 在抽穗期和灌浆期, 4个供试品种的根-冠比无显著性差异(图2-E, F)。
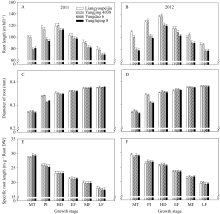
在各主要生育期内, 两优培九和扬粳4038两年的根长显著高于对照品种, 其平均根长分别较对照品种高出15.42%和15.77% (图3-A, B); 2个超级稻品种的根直径在穗分化始期显著高于对照品种, 两优培九和扬粳4038分别较其对照品种高9.50%和8.96%, 而在其他生育期无显著性差异(图3-C, D); 各生育期的比根长2个超级稻品种与对照品种均无显著差异(图3-E, F)。
 | 图3 供试水稻品种在不同生育时期根长(A, B)、根直径(C, D)和比根长(E, F)MT: 分蘖中期; PI: 穗分化始期; HD: 抽穗期; EF: 灌浆早期; MF: 灌浆中期; LF: 灌浆后期。Fig. 3 Root length(A, B), diameter of root (C, D), and special root length (E, F) of the tested rice cultivars at different growth stagesMT: mid tillering; PI: panicle initiation; HD: heading period; EF: early grain filling; MF: mid grain filling; LF: late grain filling. |
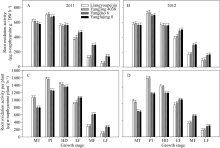
两个超级稻品种以根干重为单位的根系氧化力
在分蘖中期、穗分化始期和抽穗期均略高于对照品种, 但差异不显著; 在整个灌浆期则显著低于对照品种(图4-A, B)。超级稻品种以单株为单位的根系氧化力在分蘖中期和穗分化始期显著高于对照品种, 但在灌浆中后期则显著低于对照品种(图4-C, D)。
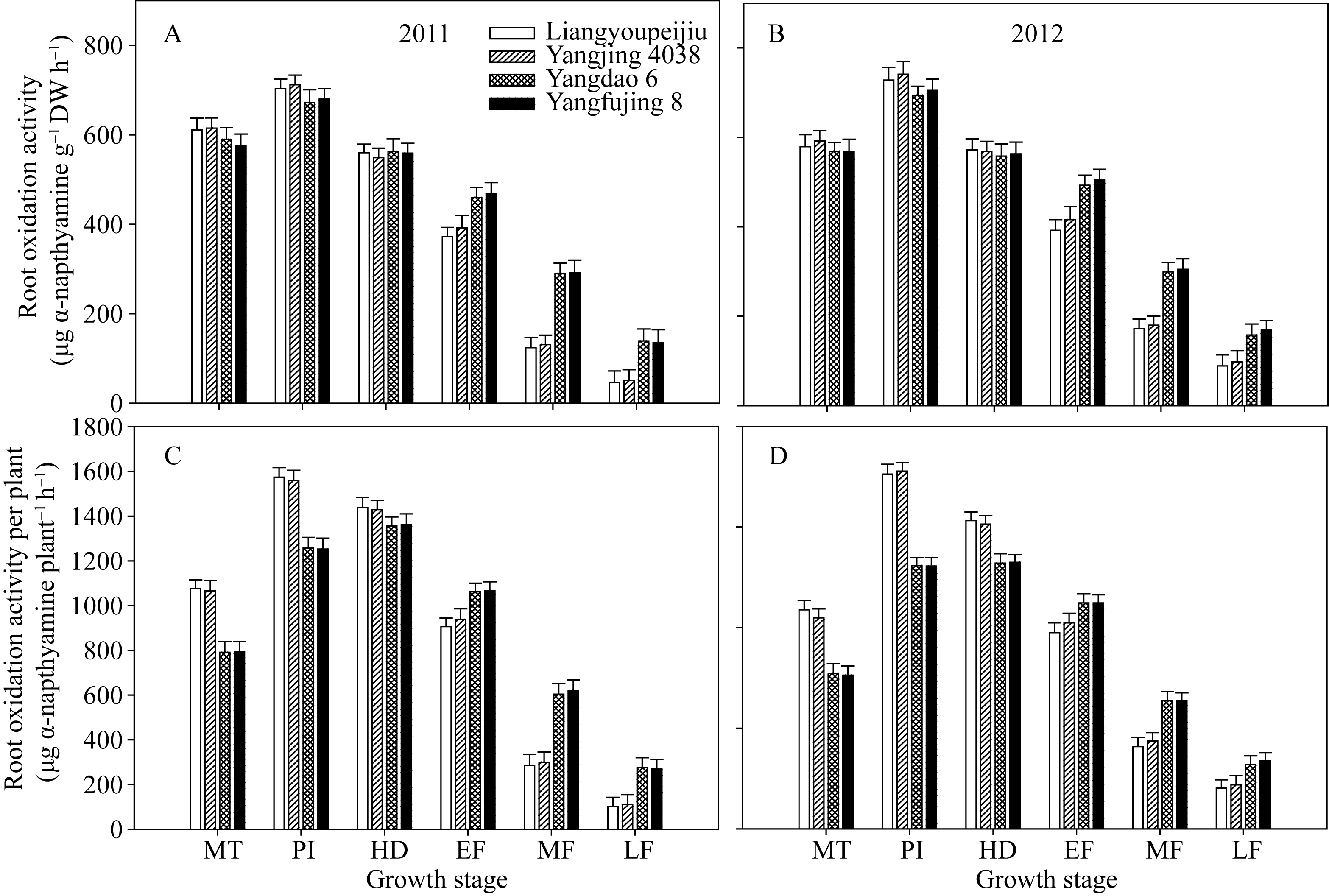 | 图4 供试水稻品种在不同生育时期单位干重根系氧化力(A, B)和单株根系氧化力(C, D)MT: 分蘖中期; PI: 穗分化始期; HD: 抽穗期; EF: 灌浆早期; MF: 灌浆中期; LF: 灌浆后期。Fig. 4 Root oxidation activity on the basis of per unit dry weight (A, B) and per plant (C, D) of the tested rice cultivars at different growth stagesMT: mid tillering; PI: panicle initiation; HD: heading period; EF: early grain filling; MF: mid grain filling; LF: late grain filling. |
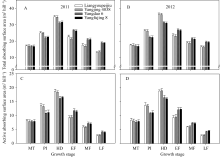
两个超级稻品种的根系总吸收表面积与根系活跃吸收表面积在分蘖中期与对照品种无显著差异; 在穗分化始期和抽穗期均显著高于对照品种; 在整个灌浆期均显著低于对照品种(图5)。
 | 图5 供试水稻品种在不同生育时期根系总吸收表面积(A, B)和活跃吸收表面积(C, D)MT: 分蘖中期; PI: 穗分化始期; HD: 抽穗期; EF: 灌浆早期; MF: 灌浆中期; LF: 灌浆后期。Fig. 5 Total absorbing surface area (A, B) and active absorbing surface area (C, D) of the tested rice cultivars at different growth stagesMT: mid tillering; PI: panicle initiation; HD: heading period; EF: early grain filling; MF: mid grain filling; LF: late grain filling. |
4个供试品种根系中玉米素(Z)+玉米素核苷(ZR)的含量随着生育进程, 呈先增后降趋势。根系中Z+ZR含量在穗分化始期和灌浆早期有2个高峰。2个超级稻品种根系中Z+ZR的含量, 从分蘖中期至抽穗期均显著高于对照品种; 但在灌浆中后期却显著低于对照品种(图6)。
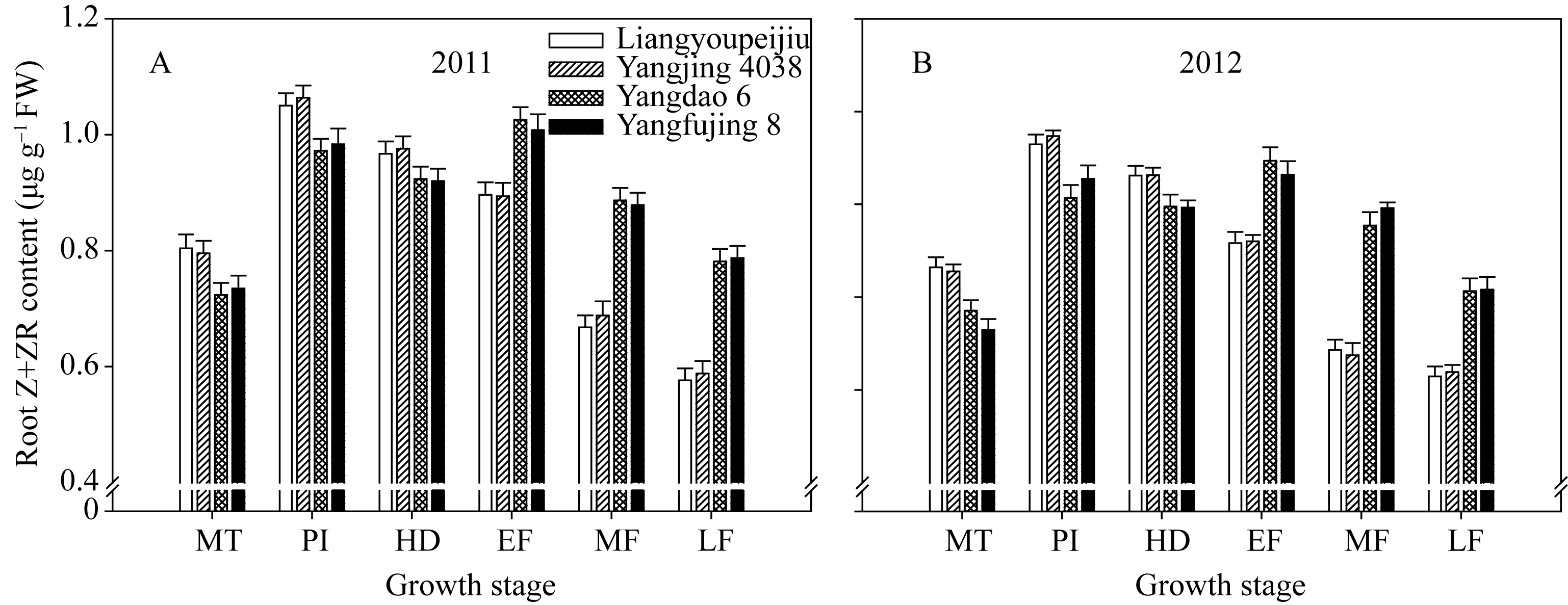 | 图6 供试水稻品种在不同生育时期根系中玉米素(Z)+玉米素核苷(ZR)含量MT: 分蘖中期; PI: 穗分化始期; HD: 抽穗期; EF: 灌浆早期; MF: 灌浆中期; LF: 灌浆后期。Fig. 6 Zeatin (Z) + zeatin riboside (ZR) content in roots of the tested rice cultivars at different growth stagesMT: mid tillering; PI: panicle initiation; HD: heading period; EF: early grain filling; MF: mid grain filling; LF: late grain filling. |
2个超级稻品种的根系伤流量强度在抽穗期显著高于对照品种, 两优培九与扬粳4038分别较对照品种高17.46%和12.90%; 但在灌浆中后期则显著低于对照品种, 在灌浆中期分别较对照品种低30.79%和27.06%, 在灌浆后期分别较对照品种低39.17%和35.47% (图7)。
 | 图7 供试水稻品种在不同生育时期根系伤流液量HD: 抽穗期; MF: 灌浆中期; LF: 灌浆后期。Fig. 7 Root bleeding amount of the tested rice cultivars at different growth stagesHD: heading period; MF: mid grain filling; LF: late grain filling. |
水稻根系的形态与生理特征及其与产量形成的关系一直是水稻生理研究的一个热点, 同时也是争论的一个焦点, 其核心问题是高产水稻品种应该具有怎样的根系形态与生理特征?有研究认为, 超高产水稻应有根系发达、发根力强、根数多、扎根深, 后期根系活力高、吸收面积广、吸收能力强、衰老较晚且缓慢, 对肥水等环境胁迫的缓冲能力强等特点[ 17, 18, 19]。也有研究认为, 根系不仅可以吸收水分和养分, 同时也需要消耗地上部产生的光合同化产物以维持根系的生长。与地上部相比, 用于生产单位根干物质量的能量是地上部的2倍, 根-冠比过大或
根量过大会消耗大量的光合同化产物, 并与穗部竞争光合同化产物, 从而不利于高产的形成[ 20, 21, 22]。以上两类不同的研究结果反映了根系形态生理特征与产量形成之间复杂的关系。我们观察到超级稻根系生物量在整个生育期均显著高于对照品种。超级稻品种发达的根系为地上部的生长提供了充足的养分、水分和植物激素, 优化了地上部结构, 改善了地上部生长, 为高产奠定了基础。叶片生长发育和光合作用所需要的无机养分大部分是由根系提供的, 根系所需的营养主要是由叶片光合作用所产生的。因此叶片的功能与根系生理活性是相互影响、相互制约的。超级稻在整个生育期具有较高的叶面积指数, 保证了充足的碳水化合物被运送至根部, 进而促进根系更好地生长。笔者认为超级稻强大的根系与地上部有着良好的互作效应, 这是超级稻获得高产的主要原因。
灌浆期超级稻根系生理活性的强弱也是争论的一个焦点。Cao等[ 23]研究认为, 超级稻品种协优9308结实期根系伤流液强度和叶片中ABA含量显著低于对照品种, 而Z+ZR含量显著高于对照。许明等[ 24]研究发现, 北方超级粳稻品种在生育中后期具有明显的根系生理优势, 其根系衰老缓慢、根系生理活性强。但也有学者观察到, 在灌浆中后期超级稻根系活性显著低于对照[ 25, 26, 27]。我们观察到, 在灌浆中后期超级稻品种的剑叶净光合速率、根系氧化力(以根干重和以单株为单位)、根系中Z+ZR含量、根系总吸收表面积与根系活跃吸收表面积以及根系伤流液强度在生育早中期均显著高于对照品种, 而在生育中后期却显著低于对照品种。超级稻生育前中期较大的根量和根系活性促进其地上部生物产量与库容的形成, 为最终获得高产打下了重要的生理基础。
我们的试验结果证实了超级稻具有巨大的产量潜力的研究报道[ 9, 28, 29]。但我们也观察到, 与对照品种相比, 超级稻品种的结实率较低。类似的结果也有不少报道[ 4, 5, 6, 7, 8]。超级稻结实率低有许多原因。一个原因可能是与较大的库容和相对较小的源供给有关; 在本试验中, 抽穗期2个超级稻品种的平均颖花数与地上部干物重之比(颖花数/g)为30.7, 对照品种为27.2。另一个重要原因可能与灌浆期的根系衰老较快有关。我们观察到, 灌浆中后期超级稻品种根系活性显著低于对照品种; 灌浆期平均根系氧化力、根中Z+ZR含量、根系总吸收表面积与根系活跃吸收表面积以及根系伤流液强度与结实率呈极显著正相关( r = 0.83**~0.96**)。因此, 在灌浆期维持较高的根系活性(根系氧化力、根中Z+ZR含量、根系总吸收表面积与根系活跃吸收表面积以及根系伤流液强度), 可以促进超级稻籽粒灌浆, 提高结实率, 进而实现超级稻的高产潜力。
与常规高产品种相比, 超级稻具有强大的产量潜力, 这主要得益于其较多的每穗颖花数和较大的总库容量。其整个生育期较大的根系和地上部生物量、生育早中期较高的单株根系氧化力、根系总吸收表面积与活跃吸收表面积、根系中较高的玉米素(Z)与玉米素核苷(ZR)的含量是其产量潜力大的主要原因。但超级稻品种存在着结实率较低的问题, 这与其在灌浆中后期根系活性以及剑叶净光合速率下降较快有关。对其改进将是提高超级稻结实率和实现其产量潜力的重要途径。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|