 -amylase and starch phosphorylase were not stimulated by anaerobic conditions, the activities of one amylase and two isozymes of starch phosphorylase, separated by native polyacrylamide gel electrophoresis, were enhanced in anaerobic conditions. The sucrose content in turion cells decreased rapidly in the early stage of anaerobic growth but then remained constant after the enhanced growth had started, suggesting active sucrose metabolism. The activities of sucrose-phosphate synthase, sucrose synthase and acid invertase, measured in vitro, were enhanced in extracts from turions in anaerobic conditions. The induction of the activity of sucrose synthase was most significant, suggesting that sucrose synthase plays an important role in sucrose metabolism in pondweed turions growing in anaerobic conditions.
-amylase and starch phosphorylase were not stimulated by anaerobic conditions, the activities of one amylase and two isozymes of starch phosphorylase, separated by native polyacrylamide gel electrophoresis, were enhanced in anaerobic conditions. The sucrose content in turion cells decreased rapidly in the early stage of anaerobic growth but then remained constant after the enhanced growth had started, suggesting active sucrose metabolism. The activities of sucrose-phosphate synthase, sucrose synthase and acid invertase, measured in vitro, were enhanced in extracts from turions in anaerobic conditions. The induction of the activity of sucrose synthase was most significant, suggesting that sucrose synthase plays an important role in sucrose metabolism in pondweed turions growing in anaerobic conditions.{kind=link}
{kind=link}
花铃期渍水对棉铃对位叶蔗糖代谢及铃重的影响
[杨长琴1, 2  , 刘瑞显
, 刘瑞显2 , 张国伟2 , 徐立华2 , 周治国1, * ]
, 刘瑞显]
|
|


*通讯作者(Corresponding author): 周治国, E-mail:giscott@njau.edu.cn, Tel: 025-84396813
第一作者联系方式: E-mail:ychq2003@qq.com
试验以美棉33B为材料, 于2007年和2009年在江苏南京(32º02′N, 118º50′E)江苏省农业科学院防雨棚的水泥池中进行。以正常灌水(土壤相对含水量为75%±5%)为对照, 于棉花中部(6~8)果枝开花时进行渍水7 d和14 d (保持地面2~3 cm的水层)处理。研究花铃期渍水对棉铃对位叶蔗糖代谢及铃重形成的影响。结果表明, 花铃期渍水条件下棉铃对位叶可溶性糖、蔗糖、淀粉含量增加, 蔗糖/淀粉比值下降, 且随渍水持续期的延长变化幅度增大, 表明花铃期渍水致使叶片蔗糖外运受阻且随渍水时间的延长外运受阻加重; 棉铃对位叶蔗糖磷酸合酶、蔗糖合酶和酸性转化酶活性增强, 且随渍水持续期的延长和果枝部位的上升而增大; 棉铃对位叶可溶性糖、蔗糖和淀粉含量与蔗糖磷酸合酶、蔗糖合酶及酸性转化酶活性显著正相关(
Two year experiments were conducted at experimental station of Jiangsu Academy of Agricultural Sciences of Nanjing City with waterlogging treatments for seven days and fourteen days in pools with rainproof shelter at anthesis of the 6th or 8th fruiting branches of cotton NuCOTN 33B, taking well water conditions as control. The results showed that contents of soluble sugar, sucrose and starch of subtending leaves of bolls increased and the ratio of sucrose content to starch content decreased under waterlogging as compared with control and the increase ranges rised with lasting waterlogging duration, indicating that the sucrose exportation of subtending leaves was inhibited and became worse with lasting waterlogging duration. The activities of sucrose phosphate synthase (SPS), sucrose synthase (SS), and acid invertase (AI) increased and got higher with the rise in fruit branch position and waterlogging duration. The contents of soluble sugar, sucrose and starch were positively and significantly correlated with activities of SPS, SS, and AI (
受季节性降雨和灾害性天气的影响, 在棉花生长季节时有渍水发生, 严重影响棉花的生长及产量。Range等[ 1]和郭文琦等[ 2, 3]曾报道渍水对棉花生长影响主要集中在光合性能、干物质累积、产量及根系生长等方面。对其他植物的研究发现, 碳水化合物在植物响应逆境胁迫的过程中具有重要意义, 尤其非结构性碳水化合物(如蔗糖和淀粉)的代谢在很大程度上影响着植株的生长以及对环境的响应[ 4, 5, 6]。棉铃发育所需养分主要来源于其棉铃对位叶的光合作用, 而蔗糖是光合作用的主要产物和运输形式, 因此, 开展花铃期渍水对棉铃对位叶蔗糖代谢生理的研究具重要意义。
Malik等[ 7]、Chen等[ 8]和Liao等[ 9]研究表明, 淹水胁迫下大多数不耐淹植物光合产物运输到根和竞争性库器官的量减少, 导致叶片中可溶性糖和淀粉积累, 也有研究认为植物在渍水条件下由于缺氧代谢获取的能量效率较低, 必须消耗大量的碳水化合物, 叶片非结构性糖含量降低甚至糖分迅速耗尽[ 10, 11]。造成上述分歧的原因与所用材料和处理方式等有关, 但不同研究的渍水条件下叶片碳水化合物含量的变化最终均抑制植物光合和生长。水分胁迫对植物蔗糖代谢的影响是通过影响蔗糖代谢相关酶的活性来实现的。一般认为, 植物叶片中蔗糖磷酸合酶是蔗糖合成的关键限速酶[ 12], 其活性与蔗糖含量密切相关[ 13, 14]; 蔗糖合酶既可催化蔗糖合成又可催化蔗糖分解, 植物光合组织中的蔗糖合酶催化蔗糖合成的活性较高[ 15, 16]; 叶片中酸性转化酶是蔗糖降解主要酶[ 17]。叶片中蔗糖磷酸合酶、蔗糖合酶和酸性转化酶协调作用, 有利于形成蔗糖浓度梯度, 加快蔗糖从韧皮部筛管“卸载”到库器官。
棉铃重是棉花产量构成的重要因子。棉铃对位叶作为棉铃养分的主要供应者, 其生理特性直接影响棉铃发育所需养分的合成和转运[ 18]。张文静等[ 19]研究表明棉铃对位叶的生理特性存在基因型差异且与铃重的形成关系密切, 棉铃对位叶C/N适宜、内源保护酶活性高有利于铃重提高[ 20], 最近研究发现, 外源6-BA显著提高棉铃对位叶蔗糖含量和转化率及淀粉含量, 促进光合产物向棉铃的运输, 从而提高铃重[ 21]。但未见有关花铃期渍水如何影响棉铃对位叶蔗糖代谢及铃重的报道。本试验在池栽条件下研究花铃期渍水对棉花不同果枝部位叶片碳水化合物动态变化、蔗糖代谢及其对铃重的影响, 以期明确花铃期渍水影响棉花铃重的叶片蔗糖代谢生理。
试验于2007年和2009年在江苏南京(32°02′N、118°50′E, 长江流域下游棉区)江苏省农业科学院防雨棚水泥池进行, 供试土壤为黄棕壤土。2007年和2009年土壤分别含有机质15.3 g kg-1和14.3 g kg-1, 全氮0.8 g kg-1和0.9 g kg-1, 碱解氮63.4 mg kg-1和52.6 mg kg-1, 速效磷29.9 mg kg-1和23.8 mg kg-1, 速效钾107.5 mg kg-1和93.2 mg kg-1。供试棉花品种为美棉33B。2007年于4月17日播种, 5月17日移栽; 2009年于4月25日播种, 5月22日移栽。两年种植密度分别为每公顷3.6万株和3.9万株, 小区面积4.5 m × 3.3 m, 4个重复。施纯氮225 kg hm-2、过磷酸钙750 kg hm-2、氯化钾225 kg hm-2, 基肥和初花肥各占50%。其他措施按高产要求进行。
于中部6~8台果枝进入开花期(2007年7月18日; 2009年7月24日)开始水分处理。2007年设2个处理, 即对照和渍水14 d。对照全生育期土壤相对含水量(土壤相对含水量=土壤含水量/田间持水量)保持在(75±5)%; 渍水14 d处理保持地面2~3 cm的水层, 持续时间为14 d, 渍水结束后土壤相对含水量自然减少至(75±5)%。2009年设3个处理, 即对照、渍水7 d和渍水14 d, 处理方式同2007年。
1.2.1 碳水化合物含量与酶活性 试验于棉株2~4 (LFB)、6~8 (MFB)、10~12 (UFB)果枝第一、二果节开花时, 挂牌标记当日所开白花。从花后10 d开始每7 d取生长发育一致棉铃8~10个及其对位叶片(每天上午9:00~10:00取样, 棉铃作其他测定), 一部分叶片用液氮速冻并在-30℃保存测定酶活性; 另一部分叶片105℃杀青、80℃烘干, 粉碎过筛后作碳水化合物含量测定。用蒽酮比色法[ 22]测定可溶性总糖和淀粉含量; 用间苯二酚比色法[ 22]测定蔗糖含量。用果糖和UDPG比色法[ 23]测定蔗糖磷酸合酶和蔗糖合酶(合成方向)活性; 用DNS显色法[ 22]测定酸性转化酶活性。
1.2.2 铃重 吐絮期按果枝部位收标记的正常成熟棉铃20个, 晒干后称重。
由表1可见, 花铃期渍水条件下不同果枝部位棉铃铃重显著降低。渍水7 d处理下部、中部和上部果枝铃重分别下降7.10%、7.99%和19.55% (2009年); 渍水14 d处理则分别下降13.80%、14.06%和27.81% (2007年)和13.33%、12.80%和29.03% (2009
| 表1 花铃期渍水对棉花铃重的影响(2007年和2009年) Table 1 Effects of waterlogging on boll weight during flowering and boll-forming stage in 2007 and 2009 (g boll-1) |
年)。表明不同果枝部位间, 花铃期渍水对铃重的影响上部大于下部和中部; 渍水处理间随渍水持续期的延长铃重下降幅度增大。
2.2.1 可溶性糖含量
图1表明, 花铃期渍水条件下棉铃对位叶可溶性糖含量大幅度增加, 其峰值出现时期随果枝部位上升提前。2009年渍水7 d处理下部、中部和上部果枝棉铃对位叶可溶性糖含量峰值分别出现在花后31、31和17 d; 而渍水14 d处理分别出现在花后31、31和24 d。渍水7 d处理的下部和中部果枝分别于花后24 d和17 d可溶性糖含量较高, 而这正是水分处理结束后1周时间, 表明棉铃对位叶可溶性糖含量变化是响应土壤水分变化的。其后不同果枝部位以渍水14 d处理棉铃对位叶可溶性糖含量较高, 表明随渍水持续时间的延长, 棉铃对位叶增加可溶性糖含量以适应土壤水分变化。花后38 d渍水14 d处理不同果枝部位棉铃对位叶可溶性糖含量均降低, 表明叶片中可利用性糖含量降低。
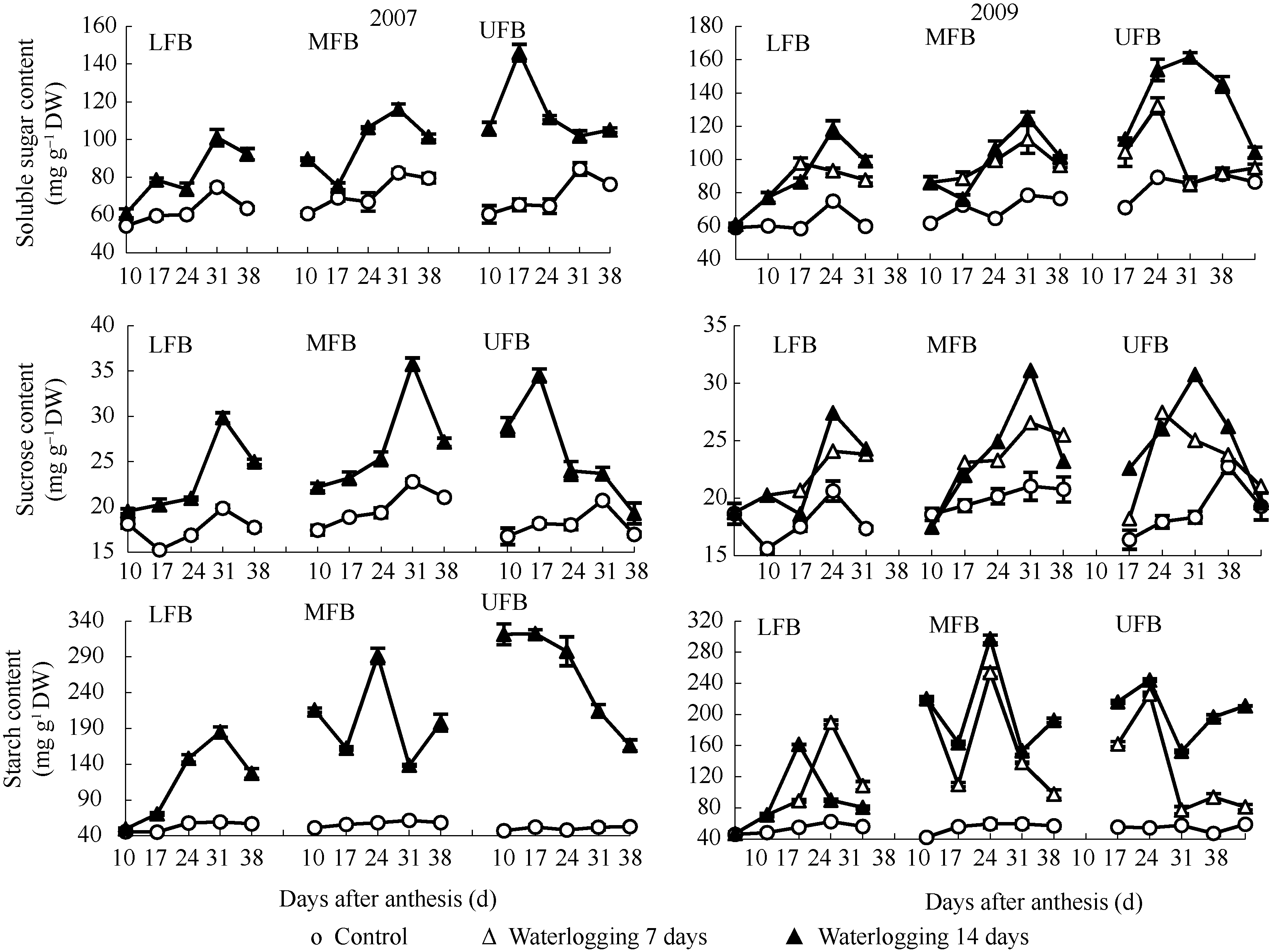 | 图1 花铃期渍水对棉铃对位叶碳水化合物含量的影响(2007年和2009年)LFB、MFB和UFB分别代表下部、中部和上部果枝。Fig. 1 Effects of waterlogging on carbohydrate contents of subtending leaves of bolls during flowering and boll-forming stage in 2007 and 2009LFB, MFB, and UFB stand for low fruiting branch, middle fruiting branch, and upper fruiting branch, respectively. |
2.2.2 蔗糖含量
与可溶性糖相似(图1), 蔗糖峰值期含量以渍水14 d处理较高。花后38 d渍水14 d处理中、上部果枝棉铃对位叶蔗糖含量降低, 表明叶片可供外运蔗糖减少。
2.2.3 淀粉含量
由图1可见, 花铃期渍水条件下棉铃对位叶淀粉含量大幅度高于对照, 且中部和上部果枝增加幅度大于下部果枝, 淀粉含量峰值出现的时间随果枝部位上升提前, 如2009年渍水7 d处理下部、中部和上部果枝棉铃对位叶淀粉含量峰值分别出现在花后31、24和17 d; 而渍水14 d处理分别出现在花后24、24和17 d。渍水处理间相比, 中部和上部果枝棉铃对位叶淀粉含量以渍水14 d处理较高, 尤其在花后后期仍保持较高水平。表明随渍水持续期的延长, 渍水对棉铃对位叶蔗糖外运抑制增强。
2.3.1 蔗糖磷酸合酶活性 图2表明, 花铃期渍水条件下棉铃对位叶蔗糖磷酸合酶的活性增加, 随着果枝部位的上升, 渍水处理与对照棉铃对位叶蔗糖磷酸合酶活性差异变大, 且其活性峰值出现时期提前。2009年渍水处理下部、中部和上部果枝棉铃对位叶蔗糖磷酸合酶活性峰值分别出现在花后31、24和17 d。渍水处理间相比, 以渍水14 d处理棉铃对位叶蔗糖磷酸合酶活性较高。表明随渍水时间的延长, 棉铃对位叶通过蔗糖磷酸合酶活性的提高以适应土壤水分变化。
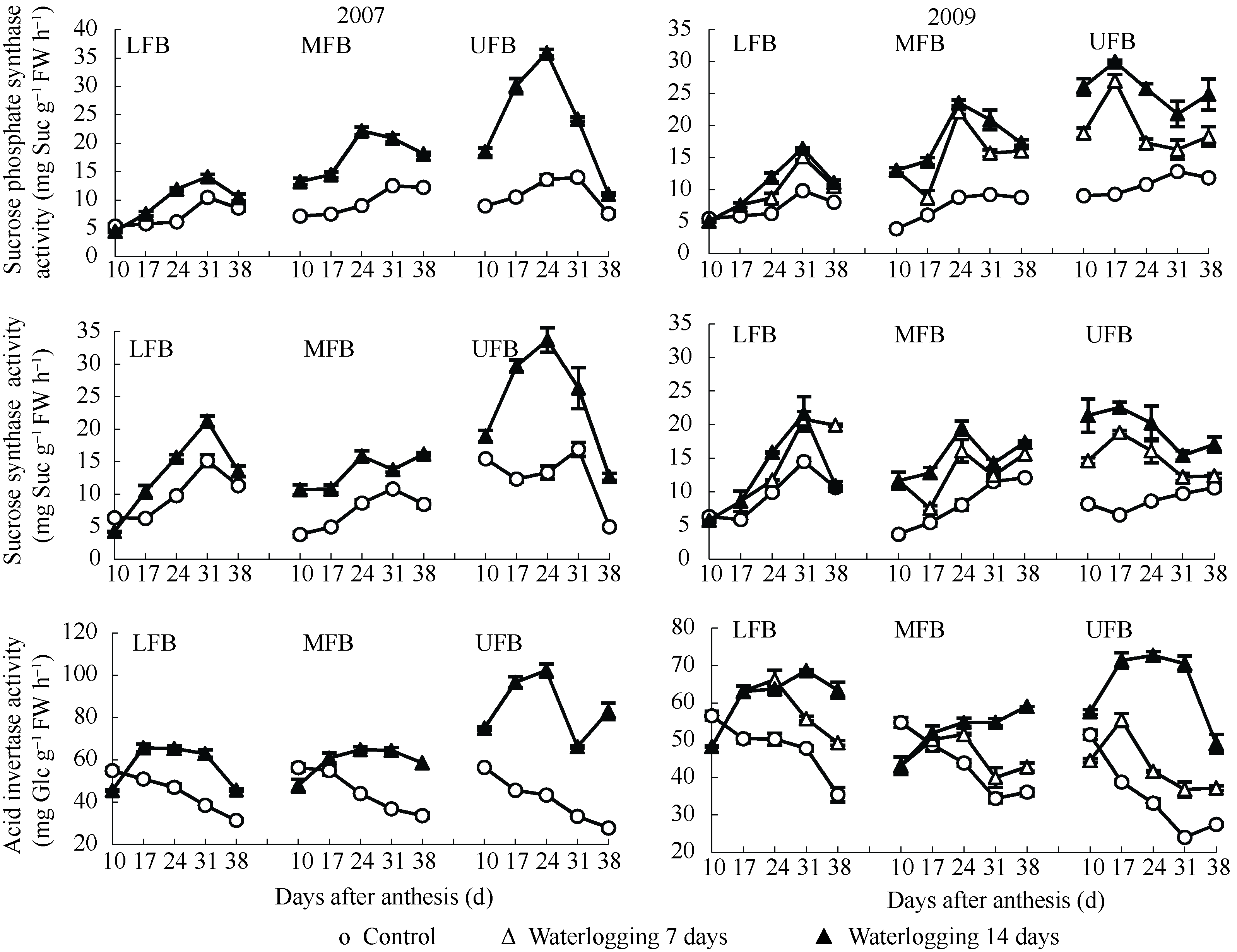 | 图2 花铃期渍水对棉铃对位叶蔗糖代谢相关酶活性的影响(2007年和2009年)LFB、MFB和UFB分别代表下部、中部和上部果枝。Fig. 2 Effects of waterlogging on sucrose metabolism enzyme activity of subtending leaves of bolls during flowering and boll-forming stage in 2007 and 2009LFB, MFB, and UFB stand for low fruiting branch, middle fruiting branch, and upper fruiting branch, respectively. |
2.3.2 蔗糖合酶活性 图2表明, 花铃期渍水条件下棉铃对位叶蔗糖合酶活性动态与蔗糖磷酸合酶活性动态相似, 渍水处理间相比, 以渍水14 d处理的酶活性较高。
2.3.3 酸性转化酶活性 图2表明, 渍水7 d处理花后17 d后棉铃对位叶酸性转化酶活性动态与对照相似但高于对照。渍水14 d处理下部和中部果枝花后10 d棉铃对位叶酸性转化酶活性略低, 其后大幅上升, 因而有利于花后初期叶片蔗糖外运但后期可供外运蔗糖减少。随花后天数增加和果枝部位上升, 渍水14 d处理酸性转化酶活性与对照及渍水7 d处理间差异变大, 如2009年渍水14 d处理下部、中部和上部果枝花后31 d棉铃对位叶酸性转化酶活性分别是对照的1.43、1.59和2.94倍; 是渍水7 d处理的1.23、1.37和1.91倍。表明渍水时间延长对棉铃对位叶酸性转化酶活性影响加大, 影响叶片可供外运的蔗糖水平。
花铃期渍水条件下棉铃对位叶中碳水化合物含量、蔗糖代谢酶活性均大幅度增加。相关分析发现(表2), 对照棉铃对位叶可溶性糖、蔗糖和淀粉含量与蔗糖磷酸合酶呈极显著正相关( P<0.01)、与蔗糖合酶活性呈正相关但未达显著水平、与酸性转化酶活性则呈显著负相关( P<0.05); 渍水处理未改变棉铃对位叶可溶性糖、蔗糖和淀粉含量与蔗糖磷酸合酶、蔗糖合酶的关系, 且与蔗糖合酶活性的相关达极显著水平( P<0.01), 但棉铃对位叶蔗糖、可溶性糖和淀粉含量与酸性转化酶活性呈显著正相关( P<0.05)。表明对照棉铃对位叶酸性转化酶活性的增加促进了蔗糖的降解外运, 而渍水处理棉铃对位叶酸性转化酶活性的增加促进了蔗糖降解的同时叶片中碳水化合物含量并未降低, 即酸性转化酶活性的增加并未促进叶片蔗糖外运。
| 表2 棉铃对位叶碳水化合物含量与蔗糖代谢相关酶活性相关关系 Table 2 Relationship between carbohydrate contents and enzyme activities related to sucrose metabolism of subtending leaves of bolls |
棉铃对位叶碳水化合物是光合作用的产物, 蔗糖则是光合作用的主要产物, 也是向库器官长距离运输的主要形式。叶片中蔗糖的水平取决于光合速率、蔗糖代谢和蔗糖外运的协调作用[ 24]。郭文琦等[ 25]盆栽试验结果表明, 渍水条件下棉株光合速率下降, 光合产物减少, 致使铃数减少、铃重降低。但有关棉铃对位叶-棉铃蔗糖代谢及运转的生理机制缺乏研究。本文研究发现, 花铃期渍水条件下棉铃对位叶可溶性糖、蔗糖和淀粉含量均大幅度增加, 与小麦[ 7]、宽叶独行菜( Lepidium latifolium)[ 8]等大多数不耐渍水物种具有相同的特征。利容千等[ 11]认为, 由于植物在渍水条件下缺氧代谢获取能量的效率较低, 必须消耗大量的碳水化合物, 因此, 渍水条件下叶片碳水化合物的累积可能是其耐渍水环境的重要因素[ 4]。Albrecht等[ 26]和Islam等[ 27]研究还表明, 渍水条件下叶片碳水化合累积是碳水化合物向库器官和根系运输受阻的结果, 因此, 棉铃对位叶碳水化合物累积不利于库器官-棉铃的发育。叶片蔗糖代谢变化对棉铃发育的影响一方面致使铃重降低, 就不同果枝部位而言, 渍水处理棉铃对位叶碳水化合物增加的幅度以中部和上部果枝大于下部果枝、且随渍水持续期延长而增加, 而铃重随果枝部位上升和渍水持续期的延长降幅增大; 另一方面致使棉铃脱落、铃数减少, 如2009年对照下部、中部和上部果枝成铃数分别为每株6.0、4.7和2.7个, 渍水7 d不同果枝部位成铃数分别降低了12.3%、42.4%和34.0%、渍水14 d分别降低了17.4%、35.2%和65.4%, 随果枝部位上升, 铃数降幅增大。
叶片碳水化合物水平受蔗糖代谢相关酶的调节。Yang等[ 28]和王维等[ 29]研究表明, 轻度、适度的土壤水分胁迫可诱导水稻茎部蔗糖磷酸合酶活性升高而酸性转化酶下降, 利于贮藏碳水化合物的输出, Hirano等[ 30]研究发现淹水下水稻上层叶片的蔗糖磷酸合酶活性被高度激活, Harada等[ 31]发现厌氧条件下水池草芽蔗糖磷酸合酶、蔗糖合酶和酸性转化酶活性均增加。本试验中渍水处理棉铃对位叶蔗糖磷酸合酶和蔗糖合酶活性大幅度增加, 酸性转化酶活性在花后10 d低, 其后也大幅度增加。表明渍水条件下棉铃对位叶蔗糖代谢较活跃, 蔗糖合成和降解代谢同时进行[ 32]。相关分析发现, 渍水处理棉铃对位叶可溶性糖、蔗糖和淀粉含量与蔗糖磷酸合酶、蔗糖合酶及酸性转化酶活性均呈正相关( P<0.05)。渍水逆境导致植物同化力下降和同化作用受阻, 植物蔗糖代谢改变, 而植物在渍水条件下的生存能力取决于能源物质(糖)的水平[ 33], 且植物体内的碳水化合物首先用于维持呼吸和生存能量需求[ 34], 因而推测较高的酶活性与维持植株在渍水下生存的能量需求有关[ 34], 而一旦碳水化合物含量降低甚至耗尽, 植物生长将很慢或难以恢复[ 35, 36, 37]。因此棉铃对位叶酶活性增强, 用于维持生存的基本代谢消耗增加, 而减少了棉铃对位叶蔗糖外运, 随果枝部位的上升和渍水持续期延长酶活性增加的幅度增大, 是渍水条件下铃重随果枝部位上升和渍水持续期延长降低的生理原因。本文研究还发现, 渍水7 d处理下部、上部和渍水14 d处理下部果枝棉铃对位叶蔗糖磷酸合酶、蔗糖合酶活性峰值时间与可溶性糖、蔗糖和淀粉含量峰值时间一致, 表明棉铃对位叶碳水化合物含量对酶活性的响应较快; 而渍水7 d处理中部和渍水14 d处理中部、上部果枝棉铃对位叶可溶性糖和蔗糖含量峰值迟于蔗糖代谢相关酶活性和淀粉含量的峰值时间, 表明随果枝部位上升和渍水持续期的延长, 棉铃对位叶可溶性糖和蔗糖含量对酶活性调节响应变慢, 且在糖含量峰值前碳水化合物外运已受很大影响。与不同果枝部位受渍害程度、渍水起始时作用的果枝部位及渍水持续期有关。综上, 渍水下叶片蔗糖代谢激活生成的碳水化合物首先用于维持生存的能量需求, 蔗糖外运受阻。蔗糖有效利用是影响棉铃发育的重要原因。因此, 生产中渍水条件下棉花铃数和铃重的提高可从植株生长恢复和促进蔗糖的有效利用入手, 研究结果为渍水条件下制定调节措施提高棉花产量提供了理论依据。
花铃期渍水条件下, 棉铃对位叶光合产物减少, 蔗糖代谢被激活, 碳水化合物含量大幅度增加, 维持生存需要蔗糖含量增加而外运减少, 随果枝部位的上升和渍水持续期的延长蔗糖外运受阻加剧, 最终铃重随果枝部位上升和渍水持续期延长降幅增大。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|