

{kind=link}
{kind=link}
{kind=link}
{kind=link}
花后高温胁迫下氮肥追施后移对小麦产量及旗叶生理特性的影响
[江文文 , 尹燕枰
, 尹燕枰* , 王振林* , 李勇, 杨卫兵, 彭佃亮, 杨东清, 崔正勇, 卢昆丽, 李艳霞]
, 尹燕枰, 王振林, 李勇, 杨卫兵, 彭佃亮, 杨东清, 崔正勇, 卢昆丽, 李艳霞]
|
|


*通讯作者(Corresponding authors): 尹燕枰, E-mail:ypyin@sdau.edu.cn, Tel: 0538-8242458; 王振林, E-mail:zlwang@sdau.edu.cn, Tel: 0538-8241359
第一作者联系方式: E-mail:jiangyunwen.1989@163.com
为探明施用氮肥与减缓小麦花后高温伤害的关系并阐明其生理机制, 于2011—2012年和2012—2013年连续两个生长季, 选用小麦品种山农16和济麦22, 在增温棚中进行花后11~15 d的高温胁迫处理, 比较了氮肥全作基肥(T0)、1/2基肥+1/2拔节期追肥(T1)、1/2基肥+1/2孕穗期追肥(T2) 3种施氮方案下的产量、产量构成因素、旗叶光合特性及部分酶的活性。在高温胁迫下, T2方案较T0和T1显著增加籽粒千粒重和产量, 提高旗叶谷氨酰胺合成酶(GS)活性, 增加旗叶气孔导度和光合速率, 提高旗叶过氧化氢酶(CAT)和过氧化物酶(POD)活性, 促进花前营养器官干物质向籽粒转运, 增加开花后积累的干物质对籽粒的贡献率。两个品种结果基本一致。综合两年结果可以看出, T2处理显著优于T0和T1处理, T2处理能显著缓解高温胁迫的伤害, 提高籽粒产量。
, YIN Yan-Ping, WANG Zhen-Lin, LI Yong, YANG Wei-Bing, PENG Dian-Liang, YANG Dong-Qing, CUI Zheng-Yong, LU Kun-Li, LI Yan-Xia
This study aimed to disclose the effect and physiological basis of postponed application of nitrogen fertilizer on alleviating the high-temperature damage after anthesis. In a two-year experiment from October 2011 to June 2013, winter wheat (
我国黄淮冬麦区小麦灌浆期间经常出现高温天气。据观测, 过去几十年中平均气温一直呈上升趋势, 并预测未来温度会继续上升, 出现高温天气的频率会提高[ 1, 2, 3]。小麦生育后期日最高温超过30℃会导致叶片早衰, 产量降低, 严重地区或年份减产可达10%~20%[ 4, 5]。因此, 减轻高温天气对小麦的危害对小麦生产至关重要。除了培育优良的耐热品种之外, 利用栽培管理技术也是减轻这一危害的重要途径。氮肥管理是最主要的栽培因素之一。研究表明, 不同氮肥运筹对小麦籽粒产量和花后衰老期间的代谢变化具有显著调节作用[ 6, 7, 8]。提高氮肥追施比例可在一定程度上缓解高温胁迫对小麦粒重和蛋白质质量的不利影响[ 9], 增施穗肥可减轻高温胁迫对水稻产量的不利影响[ 10], 说明氮肥运筹对高温胁迫有一定调节作用, 但其生理机制有待进一步研究。本试验利用2个小麦品种, 比较花后高温胁迫下不同时期追施氮肥对小麦产量的影响, 并阐明其生理机制, 旨在通过合理氮肥运筹来减轻高温伤害及为建立小麦抗逆栽培技术体系提供理论依据。
山东农业大学泰安试验农场试验地0~20 cm土壤含有机质12.3 g kg-1、全氮0.9 g kg-1、碱解氮87.2 mg kg-1、速效磷18.6 mg kg-1、速效钾57.5 mg kg-1。试验品种为山农16和济麦22, 分别于 2011年10月11日播种, 2012年6月9日收获; 2012年10月7日播种, 2013年6月14日收获。小区面积为3 m×3 m = 9 m2, 完全随机区组排列, 3次重复。全生育期施纯氮240 kg hm-2、P2O575 kg hm-2、K2O 120 kg hm-2, 氮肥为尿素和磷酸二铵, 磷肥为磷酸二铵, 钾肥为氯化钾。全部磷肥和钾肥于播种前与基施氮肥同时撒施于各小区, 然后翻耕; 氮肥开沟追施, 施肥前后不灌水。除温度和氮素因子外, 所有田间管理均按高产小麦管理规程进行。
共设3个施氮处理, 分别为全部基施(T0)、1/2基施+1/2拔节期追施(T1)和1/2基施+1/2孕穗期追施(T2), 其中T1处理为山东小麦高产施肥方法。
参考Xu等[ 11]的高温胁迫处理, 略有改进。增温棚用0.07 mm厚高保温高透光聚乙烯塑料薄膜制成, 面积与小区面积相同, 高约1.5 m与小麦冠层之间的距离50 cm左右。于花后11~15 d, 每天8:00—18:00盖棚模拟高温环境, 以常温为对照。棚四周开小窗保持良好通风, 高温处理期间未出现“烧叶”和明显“逼熟”现象。温湿度记录仪(TPJ-20, 浙江托普仪器有限公司)挂在棚内外距小麦群体表面约 30 cm处, 每隔1 h自动记录温度(图1)。常温下3种氮肥处理分别记做T0、T1和T2, 高温胁迫下3种氮肥处理分别记做HT0、HT1和HT2。
在开花期选择同一日开花的穗子挂牌标记, 每隔7 d取一次样, 其中花后7 d为高温处理前, 花后14 d为高温处理期间, 花后21 d和28 d为高温处理后。每次取20片旗叶, -40℃冰箱保存, 统一测定。
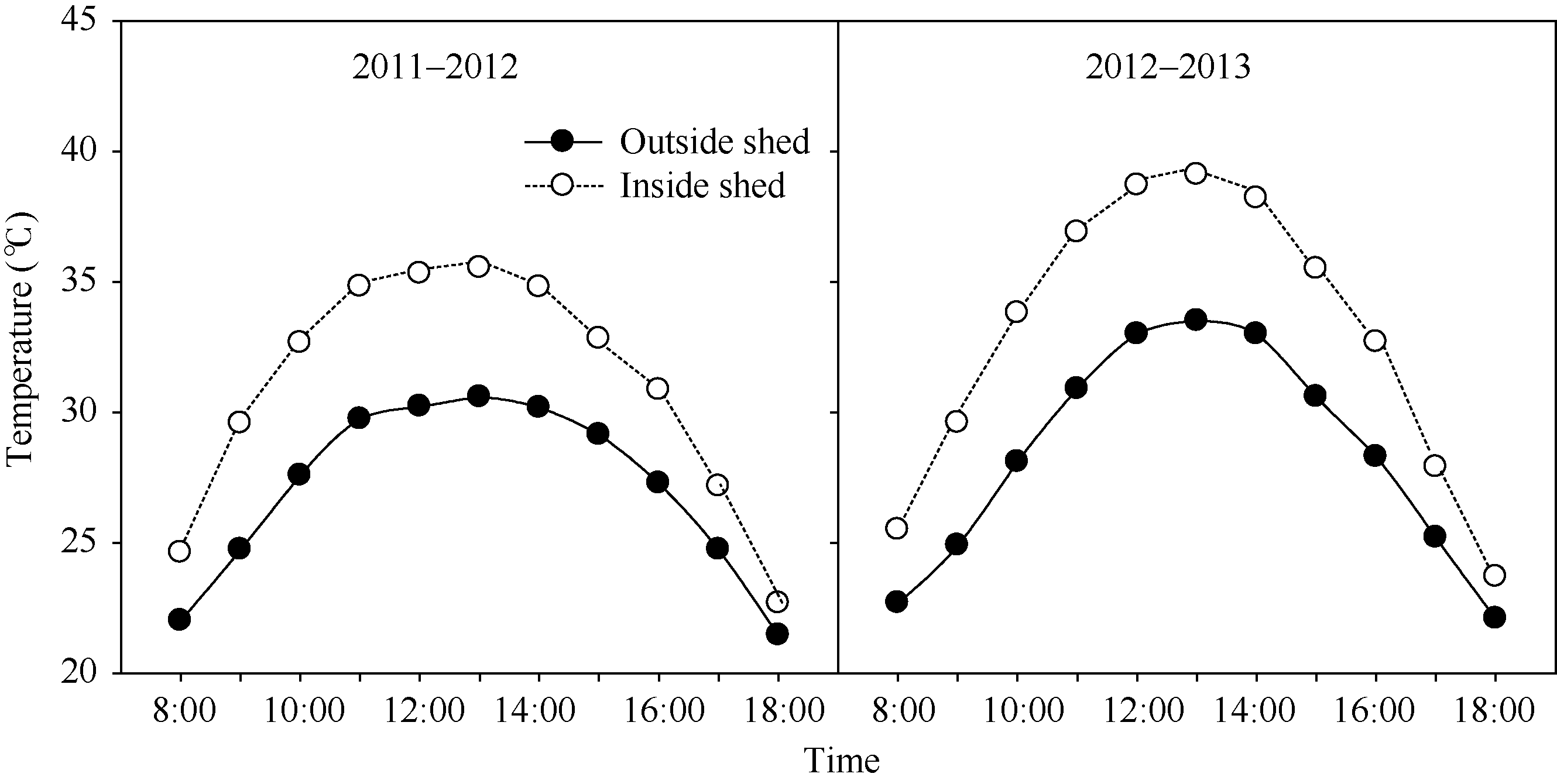 | 图1 花后11~15 d高温处理的增温棚内外温度日变化Fig. 1 Diurnal changes of temperature inside and outside plastic shed during 11-15 days after anthesis |
1.3.1 叶片酶活性 参照Pèrez-Soba等[ 12]的方法测定谷氨酰胺合成酶(GS)活性; 按Silveira等[ 13]介绍的方法测定硝酸还原酶(NR)活性; 采用氮蓝四唑(NBT)光化还原法[ 14]测定超氧化物歧化酶(SOD)活性, 以每小时反应抑制NBT光化还原50%的酶量为一个酶活力单位; 参照Tan等[ 14]的方法测定过氧化氢酶(CAT)活性; 按Klapheck等[ 15]介绍的方法测定过氧化物酶(POD)活性。
1.3.2 光合特性参数 于晴天上午8:30—11:30, 用CIRAS-2型光合仪(PP-Systems, UK)测定旗叶净光合速率( Pn)和气孔导度( Gs)。
1.3.3 干物质积累与转运 于开花期和成熟期, 按叶、茎+叶鞘、穗轴+颖壳和籽粒分别取样, 80℃烘至恒重, 称干重。营养器官开花前贮藏干物质转运量 = 开花期干重 - 成熟期干重; 营养器官开花前贮藏干物质转运率(%) = (开花期干重 - 成熟期干重)/开花期干重 × 100; 开花后干物质输入籽粒量 = 成熟期籽粒干重 - 营养器官花前贮藏干物质转运量; 籽粒产量贡献率(%) = 开花前营养器官贮藏干物质转运量/成熟期籽粒干重 × 100。
1.3.4 籽粒产量及构成因素 于小麦成熟期, 收获每小区1 m2测产和考种。
用Microsoft Excel 2003和SigmaPlot 10.0处理数据和作图, 用DPS v7.05软件对数据进行方差分析和显著性检验。
GS活性从花后7 d到花后28 d呈持续下降的趋势。山农16在高温处理前以T1处理显著高于T0和T2处理, 高温处理期间以HT1和HT2处理显著高于HT0处理, 高温处理后以T2处理显著高于T0和T1处理, 同时HT2处理显著高于HT0和HT1处理; 济麦22在花后7 d和14 d以T2处理显著高于T0和T1处理(图2)。说明高温胁迫降低了GS活性, 尤以HT0处理明显, 孕穗期追肥能提高旗叶GS活性, 增加氨的同化效率。
旗叶NR活性, 山农16在花后21 d以HT1和HT2显著高于HT0, 而花后28 d以HT2处理显著高于HT0和HT1处理; 济麦22在花后14 d和21 d均以HT0处理最高, 在花后28 d以HT1处理显著高于HT0和HT2处理。高温胁迫后, 两品种的NR活性变化不一致, 山农16以HT2处理最优(图2)。
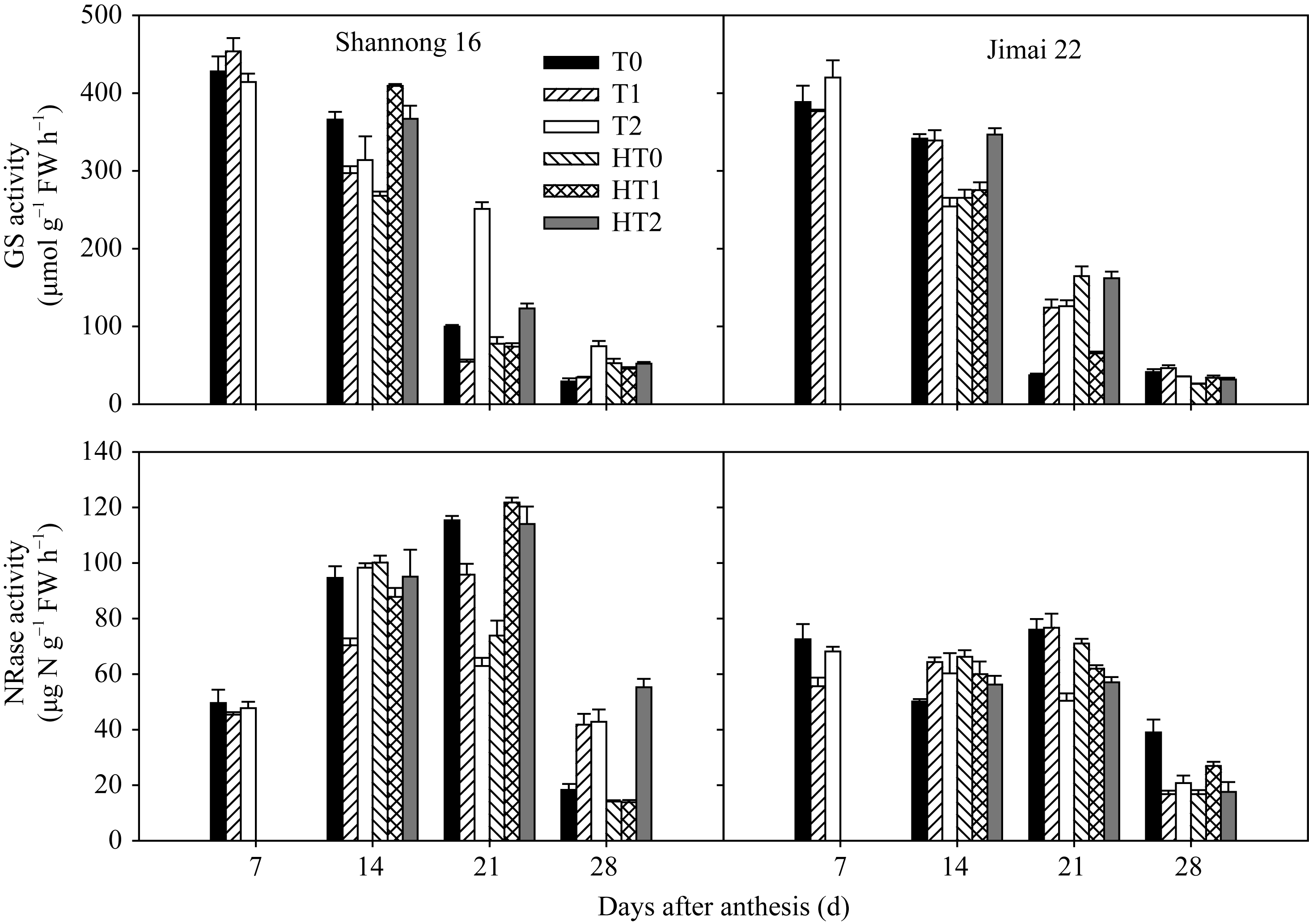 | 图2 高温胁迫和施氮处理对小麦旗叶谷氨酰胺合成酶(GS)和硝酸还原酶(NR)活性的影响(2012-2013)数据为3次重复的平均值与标准误, 不同的字母表示在0.05水平上差异显著。Fig. 2 Effects of heat stress and nitrogen application on glutamine synthetase (GS) and nitrate reductase (NR) activities in flag leaf of wheat (2012-2013)Bars superscripted with different letters are significantly different at P<0.05 as determined by LSD method. The columns and bars show the average and standard error of three replicates. |
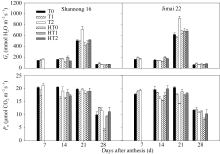
Gs在花后21 d达到最大值。山农16在花后7 d以T2处理最高, 花后14 d以HT1处理显著高于HT0和HT2处理, 在花后21 d以T2处理显著高于T0和T1处理, HT2处理显著高于HT0和HT1处理; 济麦22在花后14 d以HT1和HT2处理显著高于HT0处理, 在花后21 d, 以T2处理显著高于T0和T1处理, T1处理最低(图3)。表明孕穗期追肥更有利于气孔的张开, 利于气体交换。
山农16的 Pn值在花后7 d以T2处理最高, 在花后14 d以HT1处理显著高于HT0和HT2处理, 在花后21 d和28 d, 均以HT2处理显著高于HT0和HT1处理; 济麦22则表现为花后7 d以T2处理最高, 花后14 d以HT2处理显著高于HT0和HT1 (图3)。说明孕穗期追肥能显著增加高温处理后旗叶光合速率。
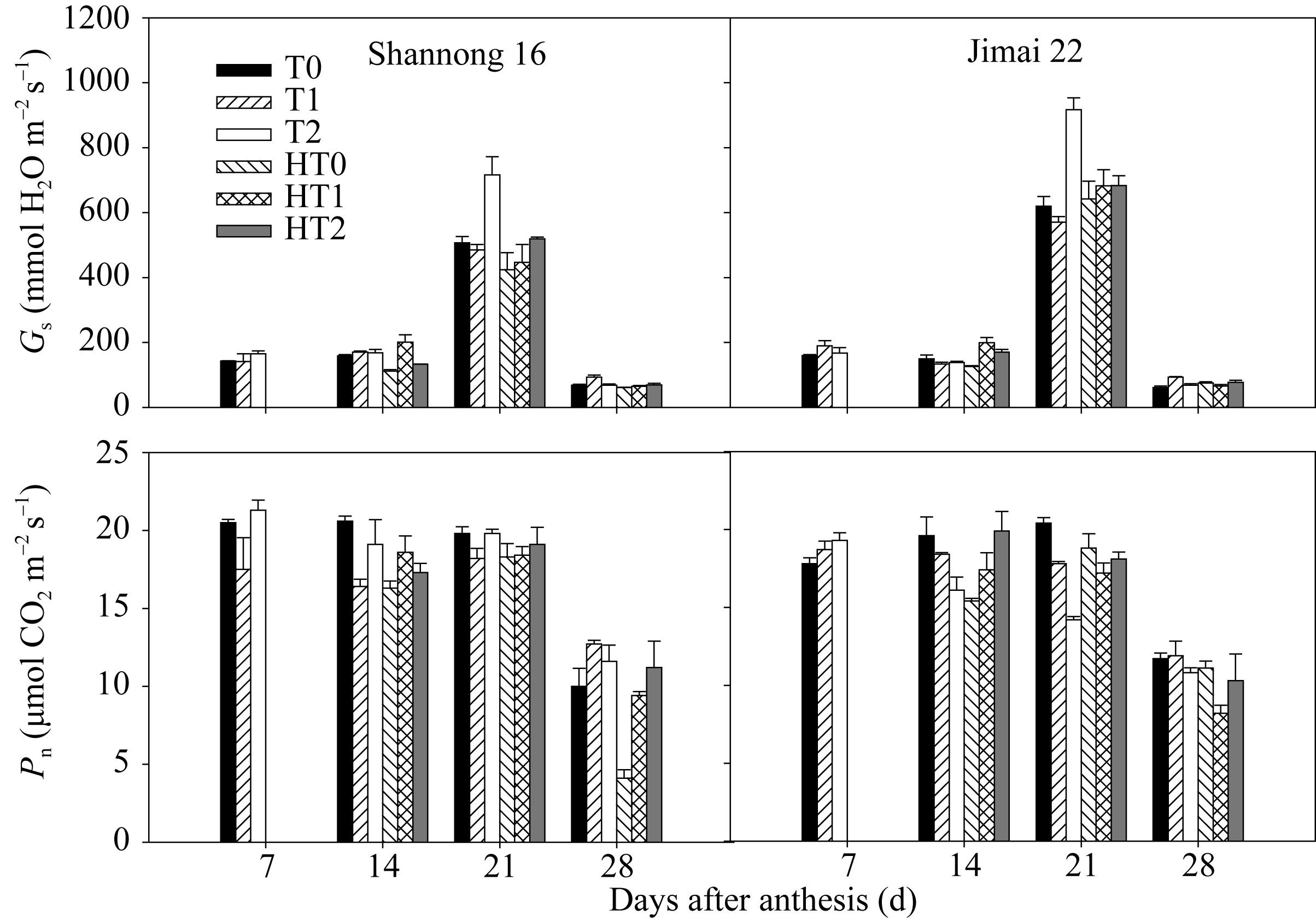 | 图3 高温胁迫和施氮处理对小麦旗叶气孔导度( Gs)和光合速率( Pn)的影响(2012-2013)数据为3次重复的平均值与标准误, 不同的字母表示在0.05水平上差异显著。Fig. 3 Effects of heat stress and nitrogen application on stomatal conductance ( Gs) and photosynthetic rate ( Pn) in flag leaf of wheat (2012-2013)Bars superscripted with different letters are significantly different at P<0.05 as determined by LSD method. The columns and bars show the average and standard error of three replicates. |
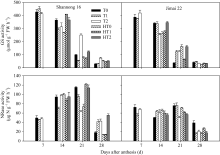
山农16旗叶SOD活性在花后7 d以T1和T2处理显著高于T0处理, 在花后28 d以HT2处理显著高于HT0和HT1处理; 济麦22旗叶SOD活性在花后14 d和21 d均以HT1处理显著高于HT0和HT2处理, 在花后28 d以HT1和HT2处理显著高于HT0处理(图4)。表明拔节期和孕穗期追肥对提高高温胁迫下旗叶SOD活性有显著作用。
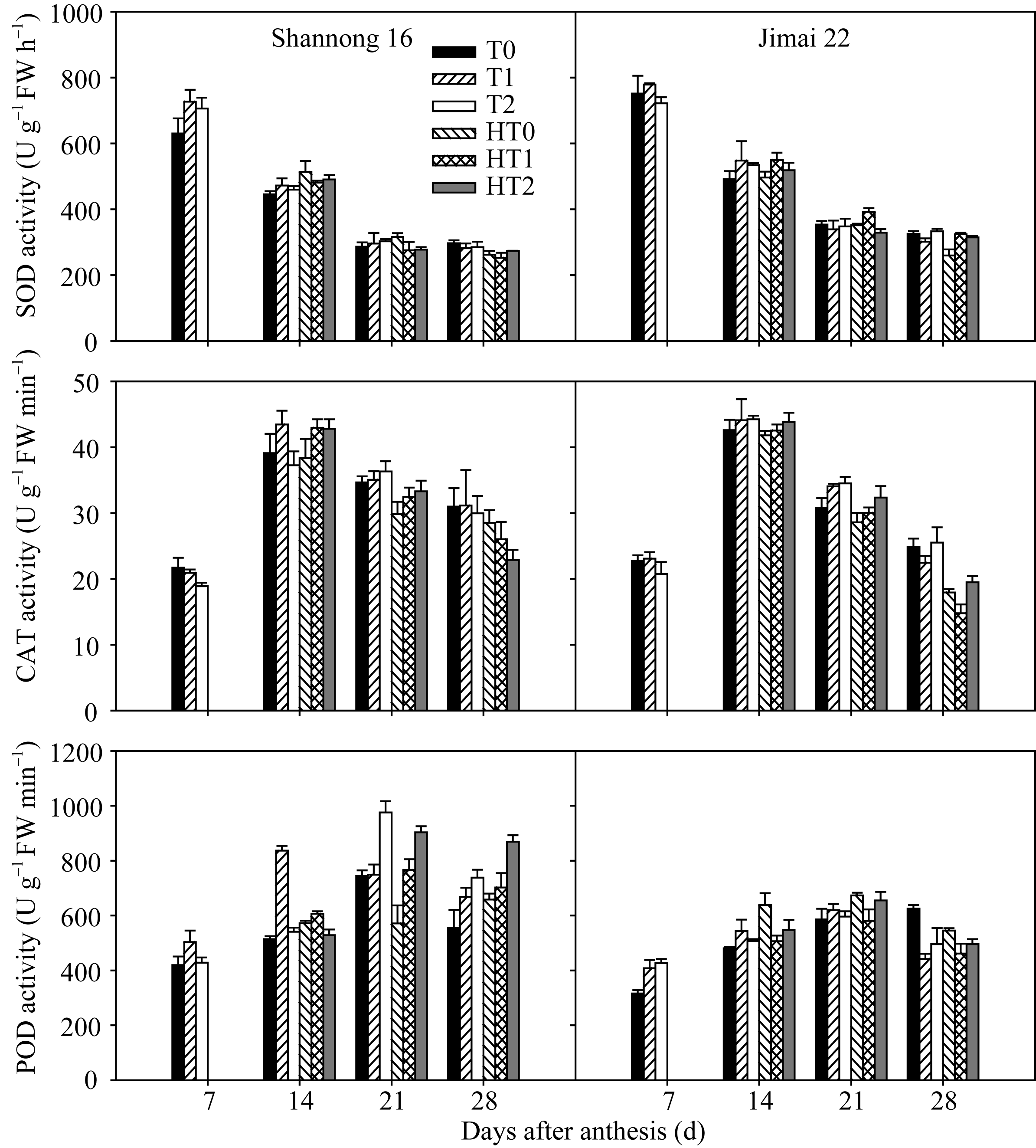 | 图4 高温胁迫和施氮处理对小麦旗叶超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性的影响(2012-2013)数据为3次重复的平均值与标准误, 不同的字母表示在0.05水平上差异显著。Fig. 4 Effects of heat stress and nitrogen application on activities of superoxide dismutase (SOD), hydrogen peroxidase (CAT), and peroxidase (POD) in flag leaf of wheat (2012-2013)Bars superscripted with different letters are significantly different at P<0.05 as determined by LSD method. The columns and bars show the average and standard error of three replicates. |
山农16旗叶CAT活性在花后14 d以T1处理显著高于T0和T2处理, 同时HT1和HT2处理显著高于HT0处理, 在花后21 d以HT1和HT2处理显著高于HT0处理; 济麦22旗叶CAT活性在花后14 d以HT2处理显著高于HT0和HT1处理, 在花后21 d和28 d均以HT2处理显著高于HT0和HT1处理(图4)。可见HT2处理较优, 高温胁迫下孕穗期追肥可以显著提高旗叶CAT活性。
山农16旗叶POD活性在花后7 d以T1处理最高, 在花后14 d以T1处理显著高于T0和T2处理, HT1处理显著高于HT0和HT2处理, 在花后21 d和28 d均以T2处理显著高于T0和T1处理, HT2处理显著高于HT0和HT1处理; 济麦22旗叶POD活性在花后7 d以T1和T2处理显著高于T0处理, 在花后21 d以HT0和HT2处理显著高于HT1处理(图4)。结果表明, 高温胁迫下孕穗期追肥能显著提高旗叶POD活性。
2011—2012年度, 山农16开花前贮藏在营养器官中的干物质开花后向籽粒的再分配量和再分配率以T2处理最高, 高温处理后向籽粒的再分配率以HT0和HT1处理显著高于HT2处理, 开花后干物质积累量以T1和T2处理显著高于T0处理, 高温处理后以HT2处理显著高于HT0和HT1处理, 开花后干物质积累量对籽粒贡献率高温处理较常温对照T0和T1处理均显著下降, 而T2处理明显升高; 济麦22开花前贮藏在营养器官中的干物质开花后向籽粒的再分配量以T2>T1>T0, 同时HT1和HT2处理显著高于HT0处理, 向籽粒的再分配率以T1和T2显著高于T0处理, 高温处理后也以HT1和HT2处理显著高于HT0处理, 开花后干物质积累量对籽粒的贡献率以T0处理最高, 同时HT0处理显著高于HT1和HT2处理, 高温处理较常温对照T0处理下降, T2处理升高, 但变化程度未达显著水平(表1)。2012—2013年度, 干物质积累与转运呈相同变化趋势。说明高温胁迫下, 孕穗期追肥能显著提高开花前贮藏在营养器官中的干物质开花后向籽粒的再分配量, 增加开花后积累的干物质对籽粒贡献率。
| 表1 不同处理对小麦开花后营养器官干物质积累量和干物质再分配的影响 Table 1 Effects of different treatments on dry matter accumulation and translocation amount of vegetative organ after anthesis in wheat |
不同处理对穗数影响差异不显著, 高温胁迫显著降低了千粒重和籽粒产量; 山农16千粒重以T2处理最高, 同时HT2处理显著高于HT0和HT1处理(2011—2012年度HT2较HT1差异未达显著水平), 籽粒产量以HT1和HT2处理显著高于HT0处理(2011—2012年度HT1较HT0差异未达显著水平); 济麦22千粒重以T1和T2处理显著高于T0处理, 同时HT1和HT2显著高于HT0处理, 籽粒产量以T1和T2处理显著高于T0处理, HT2高于HT1和HT0处理, 但差异未达显著水平(表2)。
| 表2 不同处理对小麦籽粒产量及构成因素的影响 Table 2 Effects of different treatments on grain yield and yield components of wheat |
高温是一种严重的非生物胁迫因素, 它影响植物生长发育中各种生理和生化变化[ 16, 17, 18]。植物已经形成了许多抵抗机制来减缓高温胁迫的有害影响[ 19]。研究表明, 花前高温驯化能提高小麦旗叶抗氧化酶活性, 从而减缓花后高温对旗叶的伤害[ 20]。采用轻干湿交替灌溉方式能降低水稻冠层相对湿度, 减小叶片活性氧生成速率, 增加抗氧化物质抗坏血酸和还原性谷胱甘肽含量, 提高内源细胞分裂素浓度及籽粒亚精胺和精胺浓度, 从而增强水稻抗高温能力[ 21]。可见, 不同调控措施可增强植物抗逆机制以减缓高温的伤害。本研究结果表明, 高温胁迫下氮肥追施后移至孕穗期可以显著提高小麦旗叶GS、CAT和POD的活性, 有利于增强小麦的抗热性。
关于氮肥追施时期对小麦产量的影响说法不一。于振文等[ 22]提出, 基肥和拔节肥各1/2是实现高产、优质的适宜氮肥分配方案。潘庆民等[ 23]认为, 拔节期和挑旗期追施氮肥能显著增加籽粒产量。由于氮肥追施的适宜时期与土壤肥力有关[ 24], 在运用前氮后移施肥技术时, 必须掌握好地力基础和播种量2个关键因子, 按照预定的单位面积成穗数, 灵活分配氮肥的数量和施用时期[ 25]。在本试验条件下, 拔节期和孕穗期追肥较基施全部氮肥能显著增加产量, 但高温胁迫下, 则以孕穗期追肥更有利于产量增加。
氮代谢是植物的基本生理过程之一, 植物的生长、繁殖所需要氨基酸、蛋白质、核酸和其他细胞成分的合成都涉及到氮代谢[ 26]。许多非生物胁迫都会影响植物的氮代谢。NR是氮代谢关键酶, 催化硝酸盐还原, 是植物吸收利用氮素的第一步, 其活性在一定程度上反映了植物氮代谢水平[ 27]。NR是诱导酶, 其活性容易受到光照、温度、水分等环境因素的影响。GS/GOGAT (谷氨酰胺合成酶/谷氨酸合成酶)循环是同化铵态氮的主要过程, 由其产生的谷氨酰胺和谷氨酸是植物体内合成包括氨基酸、核酸、叶绿素、多胺和碱基等含氮有机物的前体物质[ 28]。本研究表明, 高温胁迫下孕穗期追肥显著增加旗叶GS活性, 在山农16中NR活性也显著提高, 说明孕穗期追肥保证小麦花后叶片中氮代谢系统的畅通和高效运转, 提高植株的氮代谢水平, 减轻高温胁迫造成的伤害。
超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)是植物体内重要的抗氧化酶, 分别参与解除超氧化物、过氧化氢和过氧化物的毒害作用[ 29], 它们是酶促活性氧清除系统的主要组成成分, 对防止膜质过氧化, 减轻逆境造成的膜损伤和延缓植物衰老有重要作用[ 30]。有研究表明, 增加氮素营养的供应可以提高植物细胞酶促防御系统的活性[ 31]。本试验研究表明, 孕穗期追肥可以显著提高高温胁迫下旗叶CAT和POD活性, 而拔节期和孕穗期追肥对SOD活性的影响不显著。说明孕穗期追肥提高了小麦对活性氧等有害物质的清除能力, 有效减轻了高温的伤害作用。
高温胁迫下, 小麦孕穗期追肥显著增加旗叶气孔导度( Gs)和光合速率( Pn), 以保证气体正常的流通和交换。小麦灌浆期光合产物是籽粒干物质主要来源, 包括直接运输到籽粒的光合产物和暂时贮存在营养器官(主要是茎和叶)中的光合产物, 约占粒重90%~95%[ 32]。因此, 光合速率的升高有利于小麦籽粒产量的增加。小麦籽粒产量大部分来自花后干物质的积累及花前营养器官干物质的再分配[ 33]。本研究表明, 高温胁迫下,孕穗期追肥能显著促进开花前贮藏在营养器官中的干物质开花后向籽粒的转运, 增加开花后积累的干物质对籽粒贡献率, 这可能是高温处理后孕穗期追肥使小麦籽粒产量增加的一个重要原因。各处理对千粒重影响最大, 而对穗数和穗粒数都没有显著影响, 说明千粒重的变化是产量变化的关键因素。拔节期和孕穗期追肥处理对旗叶SOD活性影响差异不显著, 这表明拔节期和孕穗期追肥在一定程度上效果相似, 但综合来看, 孕穗期追肥处理对高温的缓解作用显著优于拔节期追肥。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|