{kind=link}
{kind=link}
不同抗旱性花生品种根系形态及生理特性
引用本文
厉广辉, 万勇善, 刘风珍, 张昆. 不同抗旱性花生品种根系形态及生理特性. 作物学报, 34(3): 531-541
LI Guang-Hui, WAN Yong-Shan, LIU Feng-Zhen, ZHANG Kun. Morphological and Physiological Traits of Root in Different Drought Resistant Peanut Cultivars. 作物学报, 34(3): 531-541
Permissions
LI Guang-Hui, WAN Yong-Shan, LIU Feng-Zhen, ZHANG Kun. Morphological and Physiological Traits of Root in Different Drought Resistant Peanut Cultivars. 作物学报, 34(3): 531-541
Copyright©2014, 作物学报编辑部
作物学报编辑部
不同抗旱性花生品种根系形态及生理特性
摘要
以12个花生品种为试验材料, 在人工控水条件下, 通过苗期及结荚期干旱试验, 对比分析花生品种苗期根系性状与抗旱性的关系。结果表明, 花生苗期与结荚期抗旱性基本一致。利用产量抗旱系数可把12个花生品种的抗旱性划分为强、中、弱3级, 抗旱性强的品种为A596、山花11和如皋西洋生, 中度抗旱品种为花育20、农大818、海花1号、山花9号和79266, 抗旱性弱的品种有ICG6848、白沙1016、花17和蓬莱一窝猴。山花11可作为花生强抗旱性鉴定的标准品种, 79266可作为花生弱抗旱性鉴定的标准品种。山花9号、山花11、花育20的根系具有较大的根量及根系吸收能力, 而A596、如皋西洋生、农大818、山花11具有较强的根系抗氧化能力及膜稳定性。相关分析表明, 苗期重度干旱胁迫下的单株根系干重、体积、总吸收面积、超氧化物歧化酶(SOD)活性和丙二醛(MDA)含量与品种抗旱系数的相关性达极显著水平, 对照与重度干旱胁迫下的以上性状呈极显著正相关。因此, 在花生出苗后10 d进行40%土壤相对含水量的干旱胁迫, 持续胁迫至出苗24 d的单株根系干重、体积、总吸收面积、根尖SOD活性和MDA含量可鉴定花生品种的根系抗旱能力, 正常水分下的性状值也能反映根系性状的抗旱级别。山花11可作为花生根系形态及生理优异抗旱性状鉴定的标准品种。
关键词:
花生品种; 干旱胁迫; 根系特性; 抗旱性
Morphological and Physiological Traits of Root in Different Drought Resistant Peanut Cultivars
Abstract
Drought stress is a serious constraint for peanut production worldwide. It is necessary to identify the drought resistance mechanisms of different peanut cultivars in drought-resistance breeding. Under the artificial water control condition, the peanut root morphological and physiological characteristics under drought stress at seedling stage and pod-setting stage were studied using 12 different drought-resistance peanut cultivars as material. The results showed that drought resistance at seedling stage was basically identical with that at pod-setting stage. According to yield-drought resistance coefficient, 12 peanut cultivars were divided into three grades: high-resistance, including A596, Shanhua 11, and Rugaoxiyangsheng; mid-resistance, including Huayu 20, Nongda 818, Haihua 1, Shanhua 9, and 79266; and weak-resistance, including ICG6848, Baisha 1016, Hua 17, and Penglaiyiwohou. In those peanut cultivars, Shanhua11 can be used as the standard cultivar for high drought resistance identification, and 79266 as the standard cultivar for weak one. The root drought resistance mechanism of 12 peanut cultivars were different, Shanhua 9, Shanhua11 and Huayu 20 presented a larger biomass and strong absorption capacity, while A596, Nongda 818, Shanhua 11, and Rugaoxiyangsheng had strong antioxidant capacity and membrane stability under drought stress. Correlations between drought resistance and root weight, volume, total absorption area per plant, root superoxide dismutase (SOD) activity, malondialdehyde (MDA) content under serious drought stress were significant, also under control condition. Therefore, under drought stress of 40% RWC for 10 to 24 days after germination, the root weight, volume, total absorption area per plant, SOD activity and MDA content could be used for identifies the drought resistance ability of peanut roots, the resistances degree also can be reflected by the indices above under normal water condition. Shanhua 11 can be used as a suitable standard cultivar for root morphological and physiological drought resistance characteristics identification in peanut.
Keyword:
Peanut cultivars; Drought stress; Root traits; Drought resistance
我国花生( Arachis hypogaea)总产居世界第一位, 是稳定植物油市场的重要支柱, 但主要分布于干旱半干旱的丘陵地区, 生长季降雨量不均且年度间波动较大, 有70%的花生受到不同程度的干旱威胁, 干旱引起的花生减产率平均在20%以上, 干旱是限制花生产量提高的主要因素[ 1, 2]。解决花生旱害最直接有效的方法就是选育抗旱品种。因此, 研究花生品种的抗旱性, 准确评价抗旱相关性状, 进而筛选不同种质所具有的优异抗旱性状, 是抗旱育种的重要前提。
多年来, 国内外学者在作物抗旱性鉴定方法和指标方面做了大量的研究工作[ 3, 4, 5, 6], 主要是利用多种综合性状, 采取隶属函数值、主成分分析及灰色关联等分析方法对品种的抗旱性评价分级[ 6, 7, 8]。有关花生品种的抗旱鉴定也有初步研究。已有学者利用干旱胁迫下多种农艺性状与生理性状相对值的隶属函数值和抗旱性综合评价值(D)综合评价花生品种的抗旱性, 并对部分品种的抗旱性分级[ 9, 10]; 亦有研究利用产量的抗旱系数评价花生的抗旱性, 发现部分形态性状与抗旱性显著相关[ 11]; 品种的抗旱性主要是以自然干旱条件下叶片的萎蔫指数来分级[ 12]。由于作物抗旱性是受多基因控制的数量性状, 与基因型、农艺性状及生理生化性状密切相关, 产量是植株整体生理生态机制对干旱响应的结果, 因此, 以生理性状及萎蔫指数评价品种的抗旱性存在片面性。产量抗旱系数是整株形态及生理生化性状对干旱综合反应的结果, 是品种抗旱性鉴定的准确评价
指标, 得到了一致认可[ 13, 14]。但不同品种的抗旱机制并不完全相同, 仅用抗旱系数鉴定抗旱性不能反映品种间抗旱机制的差异。
若以产量抗旱系数评价品种抗旱性, 根据形态及生理性状与抗旱系数的相关分析筛选出抗旱性状, 对比各品种抗旱性状指标可评价其优势性状, 进而可以揭示不同品种的抗旱机制, 鉴定出具有不同优异性状的抗旱花生品种。根系不但是水分吸收的主要器官, 而且通过释放干旱信号调节气孔开度, 影响水分的散失, 与作物抗旱性关系十分密切[ 15], 不同耐旱品种根系形态特征与生理活性存在显著差异[ 16, 17, 18]。本文以5大花生类型12个品种为材料, 设置苗期与结荚期干旱胁迫试验, 利用产量抗旱系数评价品种的抗旱性, 研究根系形态及生理生化性状与抗旱性的关系, 阐述鉴定不同根系抗旱性状的方法, 筛选具有的优异抗旱性状的品种, 以期为大量品种的抗旱性状鉴定及抗旱亲本选用提供试验方法和理论依据。
1 材料与方法
1.1 供试材料
选用花生栽培种5种植物学类型中12个代表性的品种(表1)。龙生型品种的抗旱性较其他植物学类型的花生强, 珍珠豆型和多粒型品种的抗旱性较弱。所选的珍珠豆型、中间型和普通型材料是山东省不同时期主推的花生品种。
| 表1 试验材料及其植物学类型 Table 1 Materials and their botanical types |
1.2 试验设计
2011—2012年在山东农业大学农学实验站, 采用盆栽进行苗期干旱试验, 以聚酯薄膜制作的防雨棚遮雨。挖取0~30 cm耕作层土壤自然风干过筛, 风干土含水量为1.62%, 田间持水量为23.56%。盆内径30 cm、深40 cm, 每盆装风干土18 kg, 灌等量水自然蒸发失水至适宜含水量时播种, 选取饱满一致的种子2粒穴播, 每盆播3穴, 每处理种植10盆。出苗后10 d开始干旱胁迫处理, 设定土壤相对含水量分别为70% (对照)、55% (轻度干旱)和40% (重度干旱), 采用每日称重法控水, 于每天早8:00称盆重补水并记录(忽略不同盆间生物量的变化), 处理期间除盆内土壤水分差异外其他管理一致。干旱胁迫至始花期恢复供水, 持续14 d。在复水前一天取每处理长势一致的3盆, 去掉地上部, 将盆土倒入网袋, 在水桶浸泡后再冲洗去泥土, 用纸巾拭去根表面的水分后测量整株根系农艺性状, 取侧根根尖部位分别编号, 封入密封袋并迅速放入液氮速冻, 带回实验室于-40℃低温冰箱中贮存, 用于生理生化指标的测定。每处理取3盆, 植株按根茎叶分开烘干称重, 用于干物质积累量和抗旱系数的计算。
于花生栽培池内进行结荚期干旱试验, 池子上方有电动防雨棚遮雨, 栽培池规格为3 m×3 m, 深1.5 m, 下不封底, 池墙由混凝土浇铸。试验材料同苗期干旱试验。采用平种覆膜, 行距40 cm, 穴距18 cm, 密度为每公顷13.9万穴, 每穴播2粒。结荚期维持50%±5%的土壤相对含水量, 持续至饱果出现(30 d), 用CNC503DR型中子水分仪测定0~80 cm土层含水量, 每20 cm一层, 每4 d测定一次, 采用测墒补灌法保证在处理水平, 以70%±5%相对含水量作为对照, 每处理3次重复。其他生育期不遮雨, 同一般高产田管理。收获时从每小区量取4.5 m2实收测产, 计算抗旱系数。
1.3 测定项目与方法
1.3.1 抗旱性 参照胡标林等[ 13]方法计算抗旱系数(drought resistance coefficient, DC)和抗旱指数(drought resistance index, DI), DC = Yd/ Yp, DI= ( Yd / Yp) × ( Yd / Ymd), 式中 Yd为干旱胁迫下产量或生物量, Yp为非胁迫下产量或生物量, Ymd为参试品种干旱胁迫下的平均产量。以抗旱系数评价品种的抗旱性。
1.3.2 根系农艺性状 取整株根系, 以排水法测定根系体积, 用吸水纸小心吸干根系表面水分, 以甲烯蓝吸附法测定根系总吸收面积和活跃吸收面积[ 19]; 以根系总吸收面积与干重的比值计算根系比吸收面积[ 15]。105℃杀青半小时, 70℃烘干至恒重称取整株根系干重。
1.3.3 根系生理特性 称取0.5 g根系样品用5 mL含1%聚乙烯吡咯烷酮(PVP)的50 mol L-1磷酸缓冲液(pH 7.0)及少量石英沙, 冰浴研磨, 匀浆于10 000× g冷冻离心20 min, 上清液即为酶提取液。参照王爱国等[ 20]的方法测定超氧化物歧化酶(superoxide dismutase, SOD)活性, 利用SOD抑制氮蓝四唑(NBT)在荧光下的还原作用, 酶活性以抑制NBT还原50%为一个酶活性单位。参照Wakamatsu等[ 21]的方法测定过氧化氢酶(catalase, CAT)活性, 以每分钟减少0.01为1个酶活单位。参照林植芳等[ 22]的方法测定丙二醛(malondialdehyde, MDA)含量。参照陈建勋等[ 23]的方法测定抗坏血酸(ascorbic acid, AsA)和还原型谷胱甘肽(glutathione, GSH)含量。
1.4 数据处理
采用Microsoft Excel 2003计算试验数据平均值, DPS7.5统计分析及SigmaPlot 10.0作图。各性状相对值(relative value, RV)=水分胁迫下性状测定值/正常供水性状测定值。
2 结果与分析
2.1 不同花生品种的产量及抗旱性
由表2可知, 苗期干旱胁迫降低了花生的干物质积累量, 随干旱胁迫加剧呈逐渐下降趋势, 而不同品种对干旱胁迫的响应存在显著差异。12个品种在轻度干旱和重度干旱胁迫下的苗期生物量抗旱系数, 2011年分别为0.77~0.99和0.58~0.82, 2012年分别为0.83~0.97和0.50~0.79。结荚期干旱胁迫降低了各花生品种的产量, 不同品种的降幅不同, 反映了品种抗旱性差异(表2)。2011年12个品种产量的抗旱系数和抗旱指数分别在0.73~0.97和0.57~1.17之间; 2012年也分别在0.76~0.99和0.49~1.24之间, 两年结果基本一致。
两年中苗期轻度干旱胁迫的生物量抗旱系数与结荚期产量抗旱系数有一定的差异( r = 0.56、0.58), 重度干旱胁迫下苗期与结荚期抗旱系数大小排序基本一致( r = 0.93、0.94, P<0.01)。表明重度干旱胁迫下生物量抗旱系数鉴定的苗期抗旱性和以产量抗旱系数鉴定的结荚期抗旱性基本一致。根据抗旱系数的大小, 12个品种抗旱性为如皋西洋生>山花11>A596>花育20>农大818>山花9号>海花1号>79266>蓬莱一窝猴>花17>白沙1016>ICG6848。
2.2 干旱胁迫下不同花生品种根系形态
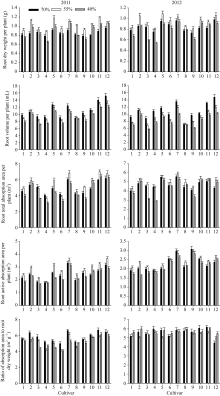
花生品种根系形态特性差异显著, 不同品种根系性状对干旱胁迫的响应不同, 根系生物量、总吸收面积、活跃吸收面积在轻度干旱下有所增加, 重度干旱下不同程度降低; 根系体积随干旱胁迫加重呈逐渐降低的趋势; 根系比吸收面积在不同干旱胁迫下表现较稳定(图1)。品种间2年结果表现的规律基本一致, 以2012年结果为例进行分析。
2.2.1 根系干重 12个品种根系干重在0.73~ 0.98 g之间, A596、海花1号、花育20和山花9号具有较大的根系生物量(图1)。苗期55%相对含量的干旱胁迫不同程度地增加了各花生品种的单株根系干重, 相对值为1.04~1.22, A596和如皋西洋生的增幅较大。土壤含水量降至40%, 不同品种单株根系干重较对照有不同程度的降低, 相对值为0.70~1.11, 农大818、如皋西洋生和山花11仍高于对照, 花育20、如皋西洋生和山花9号具有较大的根系生物量。
2.2.2 根系体积 在对照、轻度干旱及重度干旱胁迫下, 12个花生品种根系体积变化范围分别为7.13~ 14.80 mL、7.07~11.95 mL、6.03~10.27 mL; 花育20、山花9号、山花11的根系体积在对照及干旱胁迫下均显著高于其他品种(图1)。55%相对含水量的干旱胁迫下, 各品种根系体积相对值为0.78~0.99, 其中, A596、农大818与对照接近, 其他品种的降幅明显但品种间无显著差异。40%相对含水量的干旱胁迫下, 各品种根系体积相对值为0.62~0.97, 山花9号、花育20的根系体积显著高于其他品种, A596、农大818的根系体积仍与对照接近。
2.2.3 根系吸收能力 由图1可知, 山花9号、山花11、花育20的根系总吸收面积和活跃吸收面积在对照及干旱胁迫下均较高; ICG6848、白沙1016、蓬莱一窝猴则表现相反的规律。55%相对含水量干旱胁迫降低了白沙1016、花育20、花17和蓬莱一窝猴的根系总吸收面积与活跃吸收面积, 其他品种均有不同程度的增加。在40%相对含水量的干旱胁迫下农大818、山花9号和山花11的根系总吸收面积与活跃吸收面积仍高于对照, ICG6848、白沙1016与蓬莱一窝猴大幅降低且相对值显著低于其他品种。品种间的根系比吸收面积无显著差异, 相对值在0.91~1.24之间。
| 表2 苗期及结荚期干旱胁迫下不同花生品种的抗旱性 Table 2 Drought resistance of peanut cultivars under drought stress at seedling and pod-setting stages |
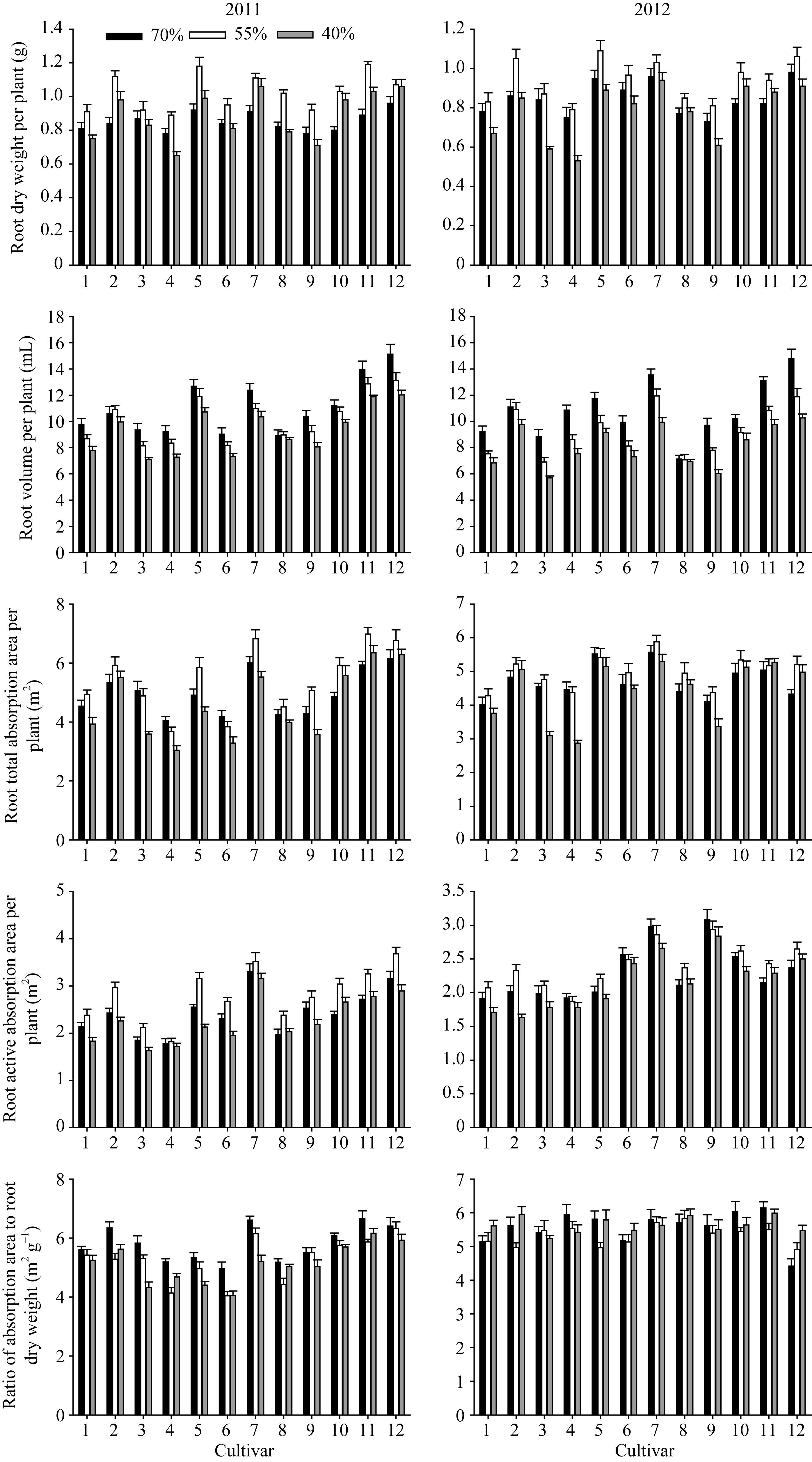 | 图1 干旱胁迫下不同花生品种苗期根系形态指标品种编号1~12分别代表79266、A596、ICG6848、白沙1016、海花1号、花17、花育20、农大818、蓬莱一窝猴、如皋西洋生、山花11和山花9号。Fig. 1 Roots morphological traits of different peanut cultivars under drought stress at seedling stageThe number of cultivars from 1 to 12 represented 79266, A596, ICG6848, Baisha 1016, Haihua 1, Hua 17, Huayu 20, Nongda 818, Penglaiyiwohou, Rugaoxiyangsheng, Shanhua 11, and Shanhua 9, respectively. |
2.3 干旱胁迫下不同花生品种根系的抗氧化能力
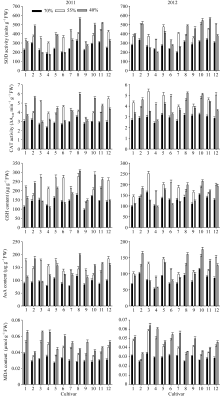
不同花生品种的根系抗氧化能力显著不同, 能表现品种间抗旱性的差异。随干旱胁迫加重, 抗旱性强的品种根系SOD及CAT活性、GSH及ASA含量呈逐渐增加的趋势, MDA含量缓慢增加; 其他品种以上抗氧化指标呈先增加后降低的趋势, MDA含量在轻度干旱时就迅速增加(图2)。两年结果的趋势基本一致, 文中分析了2012年的结果。
2.3.1 SOD、CAT活性 由图2可知, 农大818、山花11、A596、如皋西洋生的两种酶活性在对照及干旱胁迫下均较高。苗期干旱胁迫增加了两种酶的活性, 随土壤含水量的下降, A596、农大818、如皋西洋生、山花11呈逐渐增加的趋势; ICG6848、白沙1016、花17、79266、山花9号呈先增加后降低的趋势; 海花1号与蓬莱一窝猴表现稳定。55%相对含水量的干旱胁迫下, 海花1号与花育20的SOD活性增幅最大, ICG6848、白沙1016与山花9号的CAT活性显著升高, 农大818与蓬莱一窝猴的两种酶活性均未明显升高。40%相对含水量的干旱胁迫下, A596、农大818、如皋西洋生、山花11的2种酶活性及其增幅显著高于其他品种。
2.3.2 GSH、ASA含量 随干旱胁迫加重A596、农大818、如皋西洋生、山花11的根系GSH、ASA含量逐渐增加, 且在对照与干旱胁迫下均有较高的含量; 其他品种均呈先增加后降低的趋势(图2)。55%相对含水量的干旱胁迫下, 12个品种的根系GSH、ASA含量相对值分别在1.06~1.64和1.21~1.75之间, 总趋势是抗旱性强的品种其相对值较小, 其含量增加幅度较小, 抗旱性弱的品种, 增加幅度较大。40%相对含水量的干旱胁迫下, GSH、ASA含量相对值分别在0.76~1.50和0.94~1.68之间, 总趋势是抗旱性强的品种其含量增加幅度较大。
2.3.3 MDA含量 55%土壤相对含水量的干旱胁迫下, 12个品种根系MDA含量相对值在1.06~ 1.75之间, 其中, ICG6848、79266、白沙1016、花17快速增加, A596与如皋西洋生基本维持在对照水平(图2)。40%相对含水量的干旱胁迫下, 根系MDA含量相对值在1.35~2.06之间, 山花11、农大818、A596、如皋西洋生的相对值显著低于其他品种, 膜伤害程度较轻。两种干旱胁迫下均表现为抗旱性强的品种的增加幅度明显低于抗旱性弱的品种。
2.4 花生根系特性与品种抗旱性的关系
正常供水时花生根系干重、体积、吸收面积、抗氧化酶活性与品种的抗旱系数的相关性未达显著水平(表3)。55%相对含水量干旱胁迫下, 2年间仅SOD活性和MDA含量与品种的抗旱性呈极显著相关( P<0.01)。在40%相对含水量胁迫下, 2年间根系干重、体积、吸收面积、SOD、CAT活性及GSH、AsA含量与品种抗旱系数的相关性分别达显著( P<0.05)或极显著水平( P<0.01)。MDA含量在正常供水及干旱胁迫下均与抗旱性极显著负相关。轻度及重度干旱胁迫下, 根系干重、根系体积、根系总吸收面积、根系活跃吸收面积、SOD活性、AsA和MDA含量与对照水平下对应的各性状的相关性显著或极显著(表4), 抗旱性较强的花生品种在正常生长及干旱胁迫下均具有较大的根系生物量及较强的抗氧化能力。
3 讨论
不同花生品种在抗旱性方面存在遗传上的差异, 这是许多研究者的一致结论[ 2, 9, 10, 11]。本研究分别以花生苗期干旱处理的生物量抗旱系数和结荚期干旱处理的产量抗旱系数评价12个花生品种的抗旱性, 发现苗期重度干旱胁迫下品种的抗旱性与结荚期抗旱性基本一致, 相关极显著( P<0.01)。12个花生品种两年的结荚期干旱处理的产量抗旱系数分别在0.73~0.97和0.76~0.99之间, 结果基本一致, 说明品种间抗旱性存在明显的遗传差异。根据封海胜等[ 24]的标准, 把12个花生品种的抗旱性划分为强、中、弱3级。有研究表明龙生型品种A596是强抗旱性花生品种, 79266的抗旱性较弱[ 9, 11]。张智猛等[ 6, 10]研究认为抗旱系数大于0.98为抗旱性强的品种, 小于0.86为弱抗旱品种。本文发现, A596和79266的抗旱系数分别为0.97和0.87, 本试验条件下, 产量抗旱系数高于0.97为抗旱性强的品种, 0.87~0.97为中度抗旱品种, 小于0.87为抗旱性弱的品种, 与前人研究结果基本一致[ 6, 9, 10, 11]。但由于干旱时间、胁迫强度等不同, 同一品种的抗旱系数存有差异。12个品种的产量抗旱指数在0.49~1.24之间, 反映了在干旱胁迫下品种的产量差异。通过对抗旱系数与抗旱指数的综合评价, 山花11是抗旱高产品种, 如皋西洋生是抗旱低产品种, ICG6848与白沙1016属弱抗低产品种, 海花1号与山花9号属中抗高产品种。
作物抗旱机制的鉴定和评价, 不仅与所用品种抗旱性差异有关, 选用的评价性状也非常重要, 抗旱相关性状缺乏适当的鉴定方法及评价标准已成为
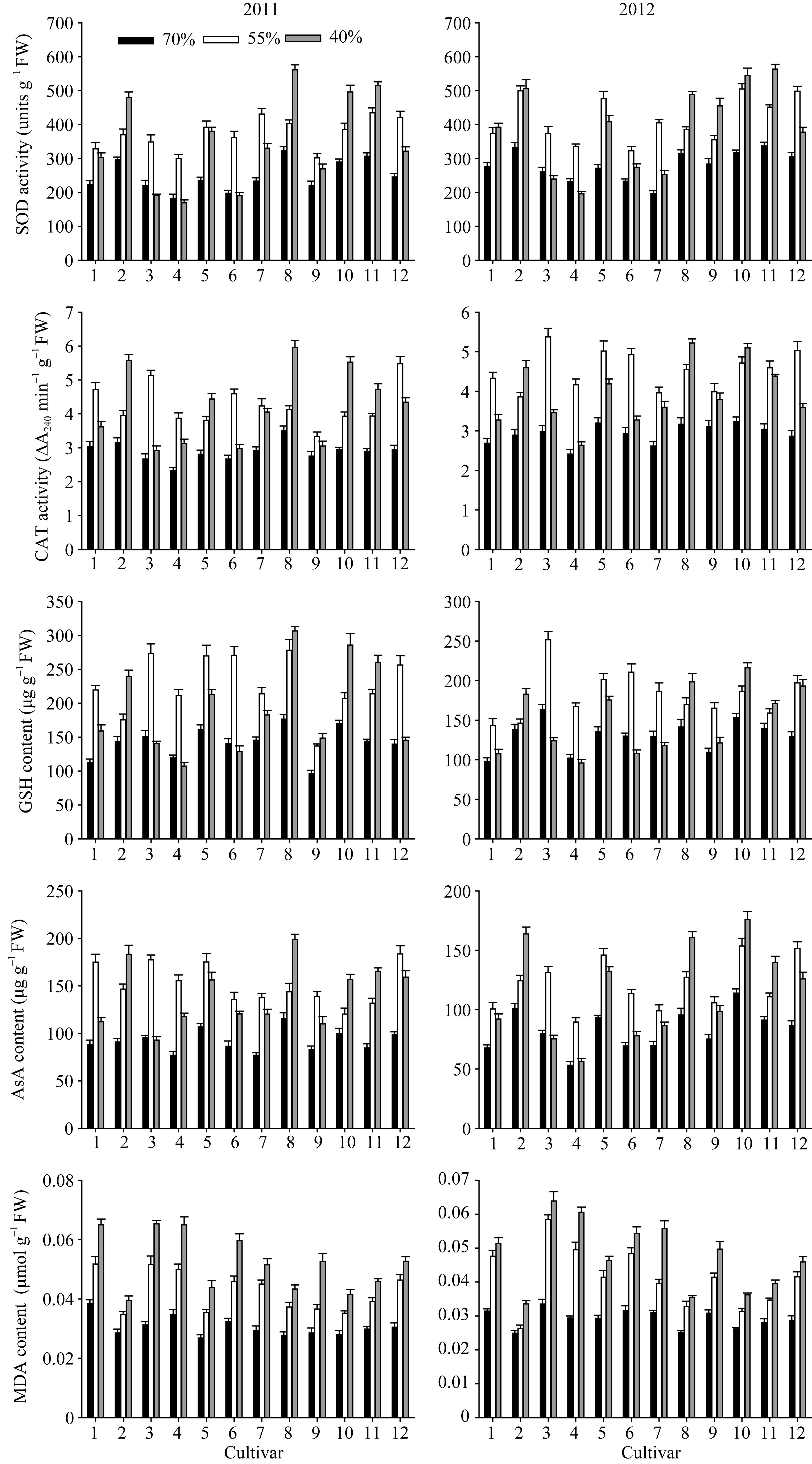 | 图2 干旱胁迫下不同花生品种苗期根系抗氧化保护相关生理性状品种编号1~12分别代表79266、A596、ICG6848、白沙1016、海花1号、花17、花育20、农大818、蓬莱一窝猴、如皋西洋生、山花11和山花9号。Fig. 2 Roots physiological traits related to antioxidant system of peanut cultivars under drought stress at seedling stageThe number of cultivars from 1 to 12 represented 79266, A596, ICG6848, Baisha 1016, Haihua 1, Hua 17, Huayu 20, Nongda 818, Penglaiyiwohou, Rugaoxiyangsheng, Shanhua 11, and Shanhua 9, respectively. |
| 表3 苗期花生根系形态及生理性状与品种抗旱性的相关系数 Table 3 Correlation coefficients of root traits of peanut cultivars with drought resistance |
TAA: 总吸收面积; TAA/DR: 比吸收面积; AAA: 活跃吸收面积。*表示 P< 0.05的显著水平,**表示 P< 0.01的显著水平。
TAA: total absorption area; TAA/DR: ratio of total absorption area to root dry weight; AAA: active absorption area.* Significant at P< 0.05;** significant at P< 0.01.
| 表4 苗期不同干旱处理下根系性状间的相关系数 Table 4 Correlation coefficients of root traits under drought stress and control |
TAA: 总吸收面积; TAA/DR: 比吸收面积; AAA: 活跃吸收面积。*表示 P< 0.05的显著水平;**表示 P< 0.01的显著水平。
TAA: total absorption area; TAA/DR: ratio of total absorption area to root dry weight; AAA: active absorption area.* Significant at P< 0.05;** significant at P< 0.01.
作物抗旱机制研究的关键问题。本研究选择的12个花生品种, 分属五大植物学类型, 抗旱性差异较大, 利于研究花生的抗旱机制。作物首先感受土壤干旱的器官是根系, 根系特性与抗旱性有密切的关系, 已成为研究和改良作物抗旱性的一个重要组成部分[ 13, 14, 15, 16]。大量的研究认为根系干重、根系体积、根系吸收面积等形态指标与品种抗旱性密切相关[ 16, 25, 26]。根系总吸收面积与活跃吸收面积是衡量根系吸收功能和活力的重要指标, 干旱胁迫下较大的根系吸收面积能提高作物吸水效率, 有利于吸收尽可能多的水分, 提高抗旱性[ 27, 28]。前人只在定性水平上研究, 无法定量描述各性状的抗旱级别, 亦没有研究具体品种的根系特性。本研究发现, 抗旱性越强的花生品种根系干重、根系体积、根系总吸收面积、根系活跃吸收面积越大, 说明较大的根量及较强的根系吸收能力可以有效抵御干旱胁迫对植株所造成的伤害。山花9号的根系干重和根系体积最大, 花育20的根系总吸收面积和活跃吸收面积显著高于其他品种, 山花11的根系体积和总吸收面积较大, 这是它们抗旱的根系机制; A596与如皋西洋生有较强的抗旱性, 但根系生物量及吸收能力未有显著优势, 可能是其他抗旱机制起主导作用。杨守萍等[ 26]研究认为应以干旱胁迫下根系性状的相对值鉴定品种的抗旱性, 刘莹等[ 29]的研究表明干旱胁迫下根系性状占整株的比重可作为衡量大豆抗旱性的指标。本研究认为, 根系形态变化是作物抗旱的机制之一, 并不是整株对干旱胁迫的综合响应, 不能全面反映品种的抗旱性。相关分析表明, 两年间重度干旱胁迫下单株根系总吸收面积与品种抗旱系数均呈极显著正相关( P<0.01), 根系干重和体积与抗旱性的相关系数也达显著( P<0.05)或极显著水平。虽然对照处理的根系性状与品种抗旱性的相关性未达显著水平, 但对照与轻度和重度干旱处理各根系性状的相关性分别达显著和极显著水平, 正常生长时具有较大根系及较强吸收能力的品种, 在干旱胁迫下仍具有显著优势。因此, 可用40%相对含水量胁迫下的单株根系干重、体积和总吸收面积鉴定花生种质的根系抗旱能力。如果鉴定的种质规模较大, 难以实现控水条件, 则正常水分下的根系性状也能反映品种间抗旱性的差异, 这减少了大量的测定工作, 便于多次重复与大规模的品种鉴定。
干旱胁迫导致植物体内活性氧水平提高时, SOD、CAT等酶活性和GSH、AsA等非酶类抗氧化物质增加, 以保持体内活性氧积累与清除系统的平衡[ 2, 30]。齐伟等[ 31]研究认为, 根系SOD、CAT、POD活性在水分胁迫下与品种的抗旱性密切相关, 抗旱性强的品种能维持较高的SOD、CAT活性; 尚晓颍等[ 32]研究认为根系SOD、CAT活性可以用来鉴别干旱胁迫下品种的抗旱性。本研究表明, 干旱胁迫下, 抗旱性越强的品种根系SOD、CAT活性及GSH、AsA含量越高, 说明相对较高的抗氧化酶活性及抗氧化剂含量能减轻活性氧对细胞的伤害, 维持细胞膜的稳定性, 保证根系生长及吸收能力正常。山花11、A596、农大818、如皋西洋生在对照和干旱条件下均保持较高的SOD、CAT活性及GSH、AsA含量, MDA含量显著低于其他品种, 这是它们抗旱的根系生理机制; ICG6848、白沙1016、花17在对照和干旱条件下均有较高的MDA含量, 膜伤害程度较严重, 这可能是它们抗旱性弱的主要原因之一。相关分析表明, 两年间重度干旱胁迫下的根系SOD活性和MDA含量均与品种抗旱性呈极显著相关, 正常灌水下根系MDA含量与品种抗旱性的相关系数也达显著水平, 它们是花生苗期抗旱的根系生理性状。本研究还发现, 对照与重度干旱处理的根系SOD活性和MDA含量呈极显著正相关, 不同品种间正常水分时根系的抗氧化能力能反映干旱胁迫下的趋势, 王思思等[ 33]在玉米研究中也得出相似结论。由此可见, 抗旱花生品种的选育应高度重视正常灌水条件下根系的生理特性。
干旱的持续时间及胁迫强度影响各抗旱性状的测定值, 直接以测定数值的大小划分抗旱性状的等级难免出现误差, 盖钧镒等[ 34]提出了大豆品种生育期组鉴定的标准品种, 依据标准品种作为参考来划分品种的生育期可以排除环境因素的影响。陈加敏[ 35]在研究大豆品种抗旱性时, 根据抗旱性强弱分别筛选出了抗旱标准品种与干旱敏感标准品种。本研究抗旱性强的3个品种中, 山花11是当前生产上使用的品种(中间型, 连续开花), 如皋西洋生、A596为龙生型, 交替开花, 匍匐生长, 生育期特长, 产量非常低, 植物学性状和生物学特性与当前推广品种差异最大, 代表性差。因此, 确定山花11为抗旱性强的标准品种。两年结果发现, 79266均是中抗级别中抗旱系数最低的, 处于中抗和弱抗的临界值。另外, 考虑到79266曾是我国北方花生产区大面积推广的品种, 目前仍有一定种植面积, 也是许多育成品种的亲本之一。属于连续开花型, 与当前推广品种的植物学性状、生物学特性、产量水平差异较小, 在生产中有广泛的代表性。而处于弱抗级别的前两位品种蓬莱一窝猴和花17植物学类型均普通型, 交替开花, 生育期较长, 产量水平较低, 植物学性状和生物学特性与当前推广品种差异较大, 代表性差。因此确定79266为花生弱抗旱性鉴定的标准品种。山花11不仅具有稳定的强抗旱性, 其根系形态和抗氧化能力在对照及干旱胁迫下均表现突出, 可作为花生根系性状鉴定的标准品种, 这样可消除因试验环境不同而产生的性状差异, 使品种抗旱性状得以量化, 鉴定结果更科学、准确。若根系性状优于山花11, 即可认为是品种的优异抗旱性状, 由此可以评价不同花生品种抗旱的主导机制。
4 结论
根据抗旱系数筛选出A596、如皋西洋生和山花11为强抗旱性花生品种。山花11可作为花生强抗旱性鉴定的标准品种, 79266可作为弱抗旱性鉴定的标准品种。在花生出苗后10 d进行40%土壤相对含水量持续14 d (出苗后24 d)的干旱胁迫下, 以单株根系干重、体积、总吸收面积、根尖SOD活性和MDA含量可鉴定花生种质的根系抗旱能力, 出苗后维持70%相对含水量至始花期的上述性状值也能反映种质根系抗旱能力的差异。山花11可作为花生根系优异抗旱性状鉴定的标准品种。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|