李彬, 邓元宝, 颜学海, 杨阳, 刘彭强, 杜勇, 谢培, 王德正, 邓其明, 李平. 一个粳稻来源抗稻瘟病基因的鉴定、遗传分析和基因定位. 作物学报, 2014, 40(1): 54-62 LI Bin, DENG Yuan-Bao, YAN Xue-Hai, YANG Yang, LIU Peng-Qiang, DU Yong, XIE Pei, WANG De-Zheng, DENG Qi-Ming, LI Ping. Identification, Genetic Analysis and Gene Mapping of a Rice Blast Resistance Gene inJaponica Rice. Acta Agron Sin, 2014, 40(1): 54-62
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Identification, Genetic Analysis and Gene Mapping of a Rice Blast Resistance Gene inJaponica Rice
LI Bin1, DENG Yuan-Bao1, YAN Xue-Hai1, YANG Yang1, LIU Peng-Qiang1, DU Yong1, XIE Pei1, WANG De-Zheng2, DENG Qi-Ming1,*, LI Ping1,*
1Rice Research Institute of Sichuan Agricultural University, Wenjiang 611130, China
2Anhui Academy of Agricultural Sciences, Hefei 230031, China
Abstract
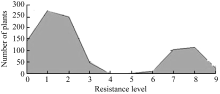
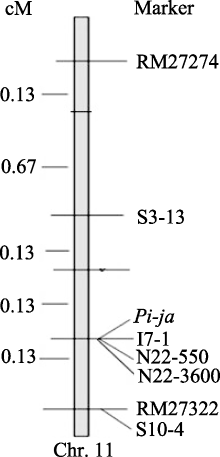
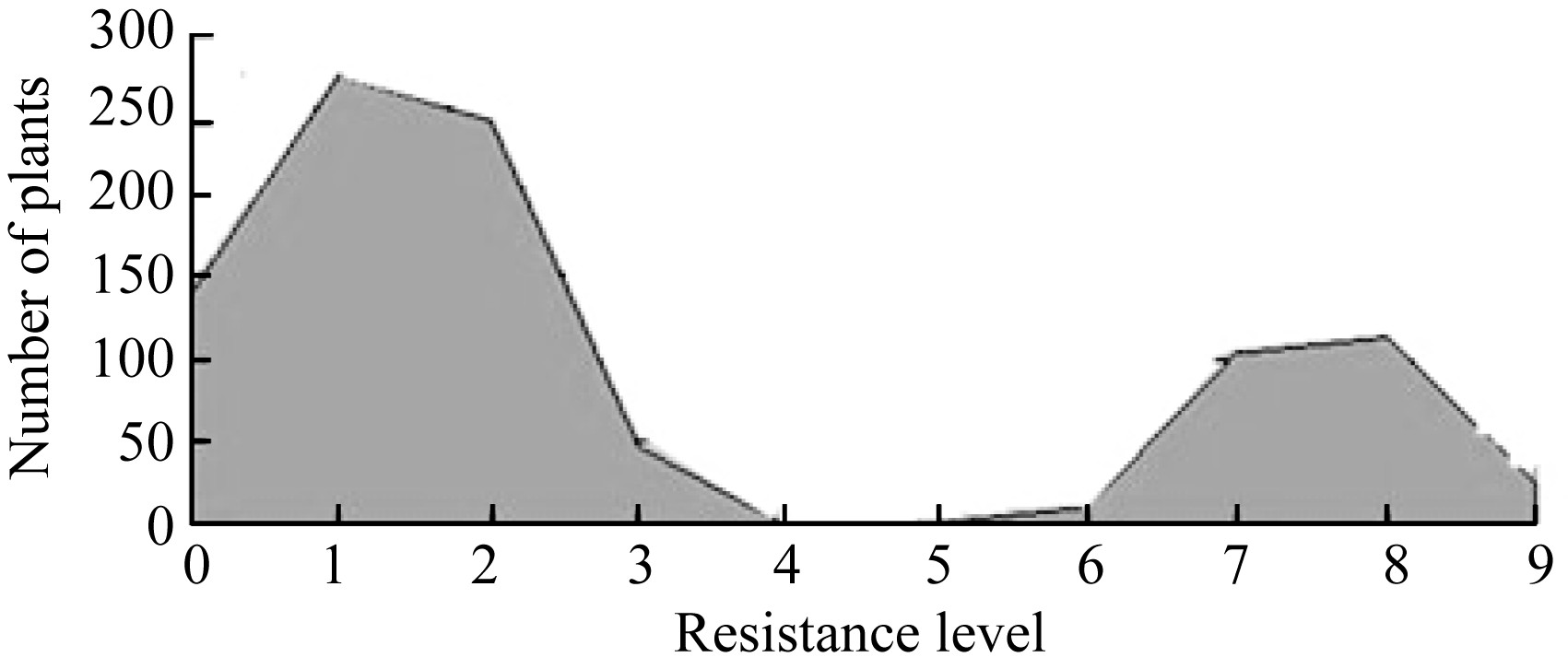
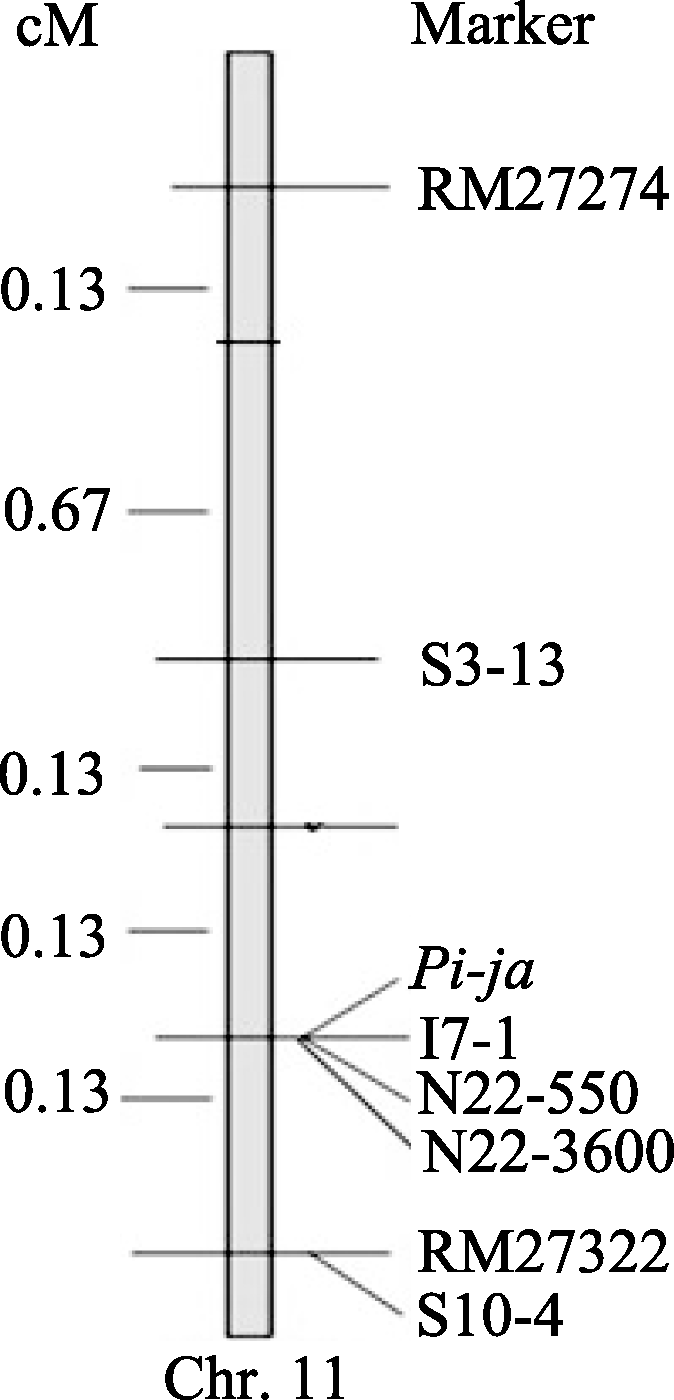
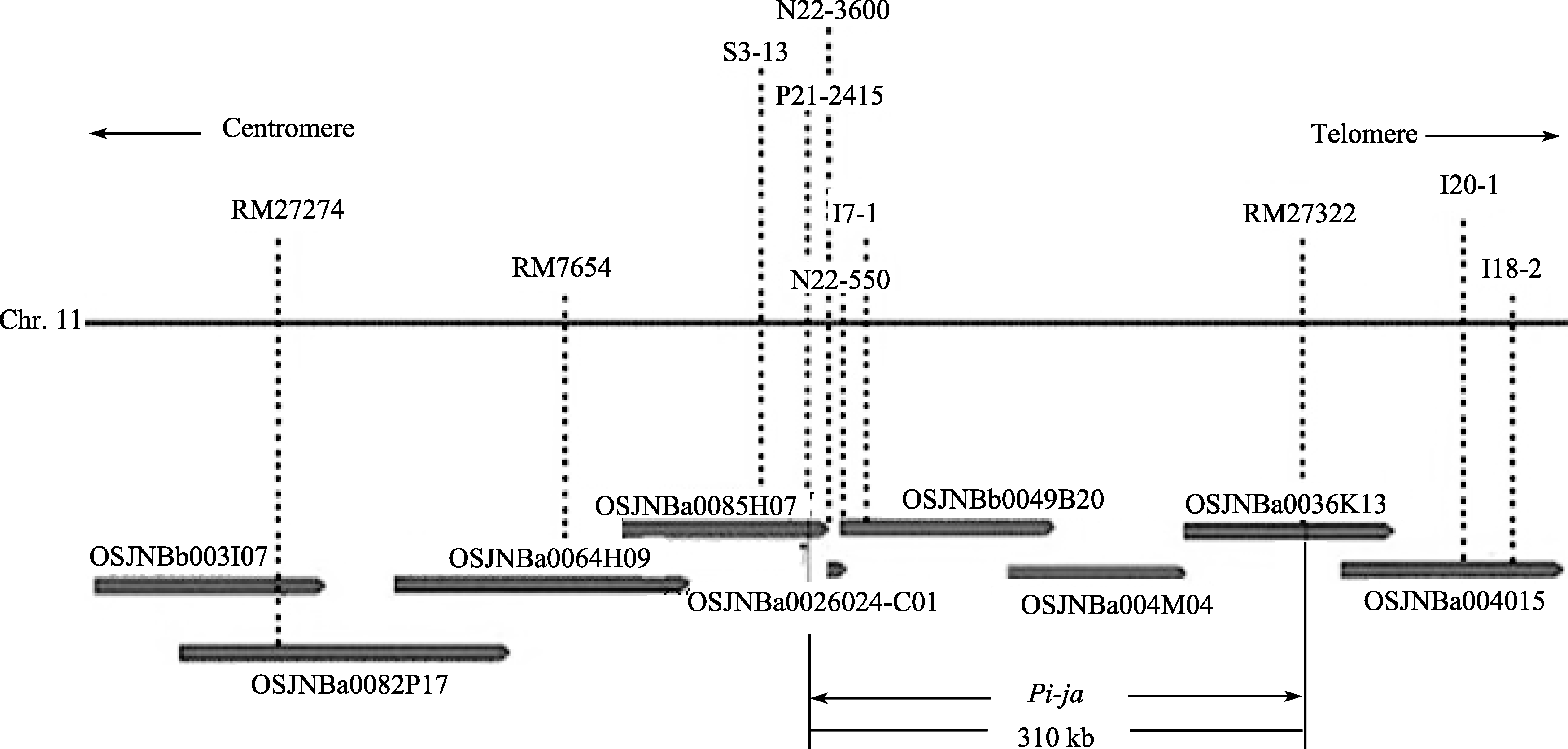
7001S is a male-sterile rice with broad-spectrum resistance to rice blast pathogens and highly resistant to 22 strains ofMagnaporthe oryzae(M. oryzae). The F2 generation of hybrid between 7001S and 80-4B showed significant resistance to rice blast pathogens. The ratio of resistant plants: susceptible plants was 3:1, indicating that the resistance of 7001S to the rice blast is controlled by one-dominant karyogene or a QTL locus. Molecular marker analysis showed that the rice blast resistance gene was located on the terminal long arm of chromosome 11 between P21-2415 and RM27322, with genetic distance of 0.27 cM and physical distance of 310 kb. Some co-segregated molecular markers were also found in this gene area and could be used for identifying candidate genes.
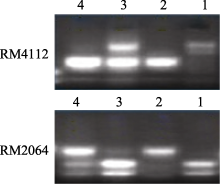
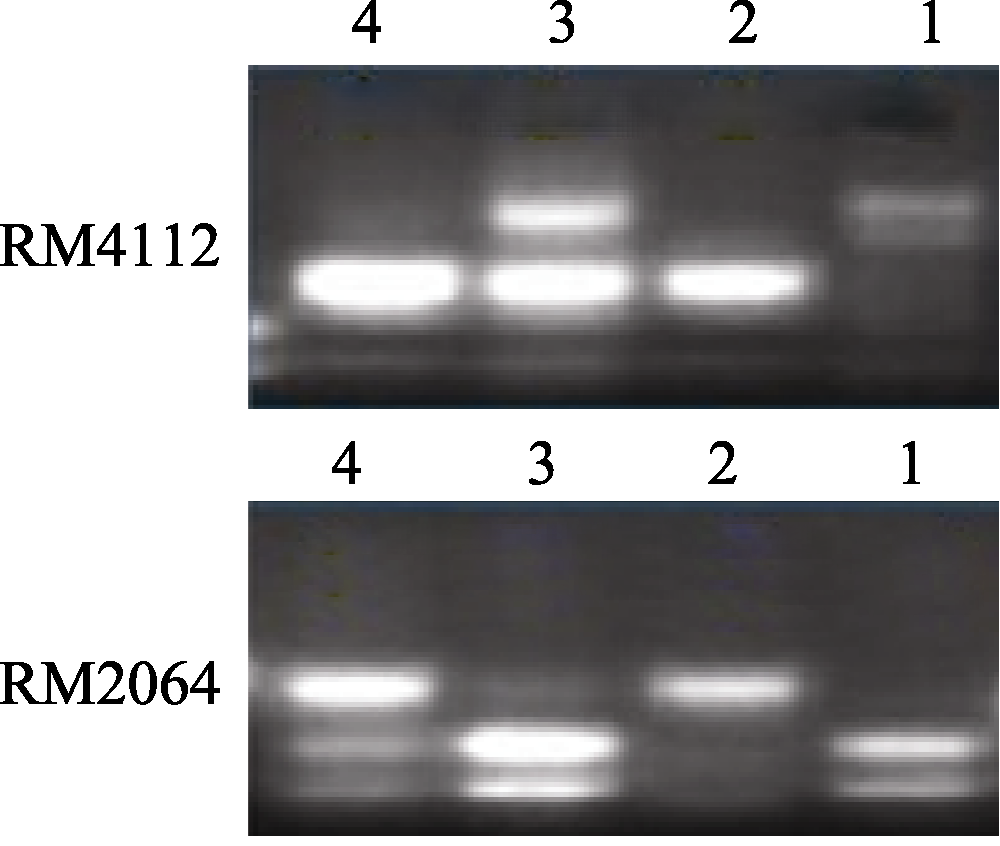
图3 SSR标记在池中的电泳分离图1: 7001S; 2: 80-4B; 3: 抗病池; 4: 感病池。Fig. 3 Segregation patterns of the SSR markers in the pool1: 7001S; 2: 80-4B; 3: resistant pool; 4: susceptible pool.
图7 7001S和80-4B中LOC_Os11g45980氨基酸序列的差异阴影部分表示抗感亲本氨基酸差异。Fig. 7 Difference of LOC_Os11g45980 amino acid sequence in 7001S and 80-4BThe shading parts indicate amino acid differences between resistant and susceptible parents.
MoffatA S. Plant genetics. Mapping the sequence of disease resistance. Science, 1994, 265: 1804-1805[本文引用:1]
[2]
SkamniotiP, GurrS J. Against the grain: safeguarding rice from rice blast disease. Trends Biotechnol, 2009, 27: 141-150[本文引用:1][JCR: 9.148]
[3]
赵国珍, 贾育林, 严宗卜, WderenC, JiaM H, 戴陆园. 一种高效便捷的水稻DNA提取法及其应用. 中国水稻科学. 2012, 26: 495-499ZhaoG Z, JiaY L, YanZ B, WderenC, JiaM H, DaiL G. An efficient, economic, and rapid rice DNA extracton method and its application. Chin J Rice Sci, 2012, 26: 495-499 (in Chinese with English abstract)[本文引用:1][CJCR: 1.4083]
[4]
雷财林, 凌忠专, 王久林. 水稻抗病育种研究进展. 生物学通报, 2004, 39(11): 4-7LeiC L, LingZ Z, WangJ L. Research advanccs in rice breeding for disease resistance. Bull Biol, 2004, 39(11): 4-7 (in Chinese)[本文引用:1][CJCR: 0.3175]
[5]
AshikawaI, HayashiN, YamaneH, KanamoriH, WuJ Z, MatsumotoT, OnoK, YanoM. Two adjacent nucleotide-binding site-leucine-rich repeat class genes are required to confer Pikm- specific rice blast resistance. Genetics, 2008, 180: 2267-2276[本文引用:2][JCR: 4.007]
[6]
BryanG T, WuK S, FarrallL, JiaY L, HersheyH P, McAdamsS A, FaulkK N, DonaldsonG K, TarchiniR, ValentB. A single amino acid difference distinguishes resistant and susceptible alleles of the rice blast resistance gene Pi-ta. Plant Cell, 2000, 12: 2033-2046[本文引用:1][JCR: 8.987]
[7]
HuaL, WuJ, ChenC, ChenC X, WuW H, HeX Y, LinF, WangL, AshikawaI, MatsumotoT, WangL, PanQ H. The isolation of Pi1, an allele at the Pik locus which confers broad spectrum resistance to rice blast. Theor Appl Genet, 2012, 125: 1047-1055[本文引用:2][JCR: 3.297]
[8]
KanzakiH, YoshidaK, SaitohH, FujisakiK, HirabuchiA, AlauxL, FournierE, TharreauD, TerauchiR. Arms race co-evolution of Magnaporthe oryzae AVR-Pik and rice Pik genes driven by their physical interactions. Plant J, 2012, 72: 894-907[本文引用:2][JCR: 6.16]
[9]
LeeS W, HanS W, SririyanumM, ParkC J, SeoY S, RonaldP C. A type I-secreted, sulfated peptide triggers Xa21-mediated innate immunity. Science, 2009, 326: 850-853[本文引用:1]
[10]
LinF, ChenS, QueZ, WangL, LiuX Q, PanQ H. The blast resistance gene Pi37 encodes a nucleotide binding site leucine-rich repeat protein and is a member of a resistance gene cluster on rice chromosome 1. Genetics, 2007, 177: 1871-1880[本文引用:1][JCR: 4.007]
[11]
LiuJ, LiuX, DaiL, WangG L. Recent progress in elucidating the structure, function and evolution of disease resistance genes in plants. J Genet Genomics, 2007, 34: 765-776[本文引用:1][JCR: 1.883][CJCR: 0.6313]
[12]
QuS D, LiuG F, ZhouB, BellizziM, ZengL R, DaiL Y, WangG L. The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-binding site-leucine-rich repeat protein and is a member of a multigene family in rice. Genetics, 2006, 172: 1901-1914[本文引用:1][JCR: 4.007]
[13]
WangW, WenY, BerkeyR, XiaoS Y. Specific targeting of the Arabidopsis resistance protein RPW8. 2 to the interfacial membrane encasing the fungal Haustorium renders broad-spectrum resistance to powdery mildew. Plant Cell, 2009, 21: 2898-2913[本文引用:1][JCR: 8.987]
ZhouB, QuS, LiuG, DolanM, SakaiH, LuG D, BellizziM, WangG. The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea. Mol Plant Microbe Interact, 2006, 19: 1216-1228[本文引用:1][JCR: 4.431]
[16]
ZhuX, ChenS, YangJ, ZhouS C, ZengL X, HanJ L, SuJ, WangL, PanQ H. The identification of Pi50(t), a new member of the rice blast resistance Pi2/Pi9 multigene family. Theor Appl Genet, 2012, 124: 1295-1304[本文引用:1][JCR: 3.297]
[17]
FukuokaS, SakaN, KogaH, OnoK, ShimizuT, EbanaK, HayashiN, TakahashiA, HirochikaH, OkunoK, YanoM. Loss of function of a proline-containing protein confers durable disease resistance in rice. Science, 2009, 325: 998-1001[本文引用:1]
[18]
HayashiN, InoueH, KatoT, FunaoT, ShirotaM, ShimizuT, KanamoriH, YamaneH, SaitoY H, MatsumotoT, YanoM, TakatsujiH. Durable panicle blast-resistance gene Pb1 encodes an atypical CC-NBS-LRR protein and was generated by acquiring a promoter through local genome duplication. Plant J, 2010, 64: 498-510[本文引用:1][JCR: 6.16]
刘华招, 陈温福, 刘延. 水稻基因分子标记的物理图谱锚定. 华北农学报, 2009, 24(增刊): 5-8LiuH Z, ChenW F, LiuY. Rice Pi genes molecular markers anchored to the physics map of rice genome. Acta Agric Boreali-Sin, 2009, 24(suppl): 5-8 (in Chinese with English abstract)[本文引用:1][CJCR: 2.0855]
[21]
ZhaiC, LinF, DongZ, HeX Y, YuanB, ZengX S, WangL, PanQ H. The isolation and characterization of Pik, a rice blast resistance gene which emerged after rice domestication. New Phytol, 2011, 189: 321-334[本文引用:2][JCR: 6.645]
[22]
YuanB, ZhaiC, WangW, ZengX S, XuX K, HuH Q, LinF, WangL, PanQ H. The Pik-p resistance to Magnaporthe oryzae in rice is mediated by a pair of closely linked CC-NBS-LRR genes. Theor Appl Genet, 2011, 122: 1017-1028[本文引用:2][JCR: 3.297]
[23]
OkuyamaY, KanzakiH, AbeA, YoshidaK, TamiruM, SaitohH, FujibeT, MatsumuraH, ShentonM, GalamD C, UndanJ, ItoA, SoneT, TerauchiR. A multifaceted genomics approach allows the isolation of the rice Pia-blast resistance gene consisting of two adjacent NBS-LRR protein genes. Plant J, 2011, 66: 467-479[本文引用:1][JCR: 6.16]
[24]
BamshadM, WoodingS P. Signature of natural selection in the human genome. Nat Rev Genet, 2003, 4: 99-111[本文引用:1][JCR: 38.075]
[25]
TianD, ArakiH, StahlE, BergelsonJ, KreitmanM. Signature of balancing selection in the Arabidopsis. Proc Natl Acad Sci USA, 2002, 99: 11525-11530[本文引用:1][JCR: 9.737]
RavensdaleM, NemriA, ThrallP H, EllisJ G, DoddsP N. Co-evolutionary interactions between host resistance and pathogen effector genes in flax rust disease. Mol Plant Pathol, 2011, 12: 93-102[本文引用:1][JCR: 3.899]
[28]
RaiA K, KumarS P, GuptaS K, GautamN, SinghN K, SharmaT R. Functional complementation of rice blast resistance gene Pi-K(H)(Pi54) conferring resistance to diverse strains of Magnaporthe oryzae. Plant Biochem Biotechnol, 2011, 20: 55-65[本文引用:1][JCR: 0.523]
[29]
李成云, 陈宗麒, 陈琼珠. 稻瘟病菌的研究进展. 西南农业学报, 1995, 8(3): 107-112LiC Y, ChenZ Q, ChenQ Z. Research progress of rice blast fungus. Southwest China J Agric Sci, 1995, 8(3): 107-112 (in Chinese)[本文引用:1]
[30]
TakahashiA, HayashiN, MiyaoA, HirochikaH. Unique features of the rice blast resistance Pi-sh locus revealed by large scale retrotransposon-tagging. BMC Plant Biol, 2010, 10(175): 1-14[本文引用:1][JCR: 3.447]
[31]
刘海, 肖应辉, 唐文邦, 邓化冰, 陈立云. 水稻两用核不育系繁殖基地计算机选择系统研制与应用. 作物学报, 2011, 37: 755-763LiuH, XiaoY H, TangW B, DengH B, ChenL Y. Development and application of a computer-aided selection system for thermo-sensitive genic male sterile rice multiplying site. Acta Agron Sin, 2011, 37: 755-763 (in Chinese with English abstract)[本文引用:1][CJCR: 1.8267]
[32]
杨仕华, 程本义, 沈伟峰, 夏俊辉. 中国两系杂交水稻选育与应用进展. 杂交水稻, 2009, 24(1): 5-9YangS H, ChengB Y, ShenW F, XiaJ H. Progress of application and breeding on two-line hybrid rice in China. Hybrid Rice, 2009, 24(1): 5-9 (in Chinese with English abstract)[本文引用:1][CJCR: 0.4895]
Rice is the staple diet of more than three billion people. Yields must double over the next 40 years if we are to sustain the nutritional needs of the ever-expanding global population. Between 10% and 30% of the annual rice harvest is lost due to infection by the rice blast fungusMagnaporthe oryzae. Evaluation of genetic and virulence diversity of blast populations with diagnostic markers will aid disease management. We review theM. oryzae species-specific and cultivar-specific avirulence determinants and evaluate efforts towards generating durable and broad-spectrum resistance in single resistant cultivars or mixtures. We consider modern usage of fungicides and plant defence activators, assess the usefulness of biological control and categorize current approaches towards blast-tolerant genetically modified rice.
ZHAO Guozhen1, JIA Yulin2,* , YAN Zongbu3 , Christopher W DEREN3, Melissa H JIA2 , DAI Luyuan1,*
赵国珍1,贾育林2,* ,严宗卜3,Christopher W DEREN3 ,Melissa H JIA2 ,戴陆园1,*
1Institute of Food Crops, Yunnan Academy of Agricultural Sciences, Kunming 650205, China;2USDAARS Dale Bumpers National Rice Research Center, Stuttgart AR 72160, USA;3 University of Arkansas,Rice Research and Extension Center, Stuttgart AR 72160, USA;
A method for DNA extraction from rice leaf, root and seed was developed, and the extracted DNA was used as a template to successfully amplify the rice blast resistance gene Pita. Profiles of Pita in 165 breeding lines detected by DNA markers were verified using differential blast races. This method involved three steps: 1) Plant tissue was placed into a 200 μL 96well plate; 2) A total of 70 μL of Buffer A containing NaOH and Tween 20 was added to each sample and incubated at 95℃ for 10 min in a PCR machine; and 3) A total of 70 μL of Buffer B containing TrisHCl and EDTA was added to each well and the resulting solution used for PCR amplification. Results demonstrated that this method of DNA extraction has the following advantages: 1) It is economical and only 4 common chemicals totaling 140 μL were used; 2) It is easy to perform, consists of only three steps, and one technician can extract hundreds of samples per day; 3) It requires a standard PCR machine; and 4) It can efficiently extract DNA from as little as a half dried seed, 5 to 20 mg of leaf tissue, or 20 mg root tissue. The resulting DNA quality was good enough to detect a single copy of the rice blast resistance gene Pita which was verified in breeding lines with the expected disease reactions. This method has proven to be highly efficient for evaluating a large number of samples.
Ashikawa I , Hayashi N , Yamane H , Kanamori H , Wu J Z , Matsumoto T , Ono K , Yano M
The rice blast resistance genePikm was cloned by a map-based cloning strategy. High-resolution genetic mapping and sequencing of the gene region in thePikm-containing cultivar Tsuyuake narrowed down the candidate region to a 131-kb genomic interval. Sequence analysis predicted two adjacently arranged resistance-like genes,Pikm1-TS andPikm2-TS, within this candidate region. These genes encoded proteins with a nucleotide-binding site (NBS) and leucine-rich repeats (LRRs) and were considered the most probable candidates forPikm. However, genetic complementation analysis of transgenic lines individually carrying these two genes negated the possibility that eitherPikm1-TS orPikm2-TS alone wasPikm. Instead, it was revealed that transgenic lines carrying both of these genes expressed blast resistance. The results of the complementation analysis and an evaluation of the resistance specificity of the transgenic lines to blast isolates demonstrated thatPikm-specific resistance is conferred by cooperation ofPikm1-TS andPikm2-TS. Although these two genes are not homologous with each other, they both contain all the conserved motifs necessary for an NBS–LRR class gene to function independently as a resistance gene.
Hua L , Wu J , Chen C , Chen C X , Wu W H , He X Y , Lin F , Wang L , Ashikawa I , Matsumoto T , Wang L , Pan Q H
We report the isolation ofPi1, a gene conferring broad-spectrum resistance to rice blast (Magnaporthe oryzae). Using loss- and gain-of-function approaches, we demonstrate thatPi1 is an allele at thePik locus. Like other alleles at this locus,Pi1 consists of two genes. A functional nucleotide polymorphism (FNP) was identified that allows differentiation ofPi1 from otherPik alleles and other non-Pik genes. A extensive germplasm survey using this FNP reveals thatPi1 is a rare allele in germplasm collections and one that has conferred durable resistance to a broad spectrum of pathogen isolates.
Kanzaki H , Yoshida K , Saitoh H , Fujisaki K , Hirabuchi A , Alaux L , Fournier E , Tharreau D , Terauchi R
Attack and counter-attack impose strong reciprocal selection on pathogens and hosts, leading to development of arms race evolutionary dynamics. Here we show thatMagnaporthe oryzae avirulence geneAVR-Pik and the cognate rice resistance (R) genePik are highly variable, with multiple alleles in which DNA replacements cause amino acid changes. There is tight recognition specificity of theAVR-Pik alleles by the variousPik alleles. We found that AVR-Pik physically binds the N-terminal coiled-coil domain of Pik in a yeast two-hybrid assay as well as in anin planta co-immunoprecipitation assay. This binding specificity correlates with the recognition specificity betweenAVR andR genes. We propose thatAVR-Pik andPik are locked into arms race co-evolution driven by their direct physical interactions.
Sang-Won Lee*, Sang-Wook Han*, Malinee Sririyanum, Chang-Jin Park, Young-Su Seo, Pamela C. Ronald†
Department of Plant Pathology, University of California, Davis, CA 95616, USA. ↵†To whom correspondence should be addressed. E-mail:pcronald@ucdavis.edu
Plants employ multifaceted mechanisms to fight with numerous pathogens in nature. Resistance (R) genes are the most effective weapons against pathogen invasion since they can specifically recognize the corresponding pathogen effectors or associated protein(s) to activate plant immune responses at the site of infection. Up to date, over 70R genes have been isolated from various plant species. Most R proteins contain conserved motifs such as nucleotide-binding site (NBS), leucine-rich repeat (LRR), Toll-interleukin-1 receptor domain (TIR, homologous to cytoplasmic domains of theDrosophila Toll protein and the mammalian interleukin-1 receptor), coiled-coil (CC) or leucine zipper (LZ) structure and protein kinase domain (PK). Recent results indicate that these domains play significant roles in R protein interactions with effector proteins from pathogens and in activating signal transduction pathways involved in innate immunity. This review highlights an overview of the recent progress in elucidating the structure, function and evolution of the isolatedR genes in different plant-pathogen interaction systems.
Kioumars Ghamkhar(1)(6),Sachiko Isobe(2),Phillip G. H. Nichols(3)(4),Troy Faithfull(1),Megan H. Ryan(4),Richard Snowball(3),Shusei Sato(2),Rudi Appels(5)
1.Centre for Legumes in Mediterranean Agriculture, The University of Western Australia, 35 Stirling Highway, Crawley, WA, 6009, Australia
6.Department of Primary Industries, Victorian AgriBiosciences Centre, 1 Park Drive, Bundoora, VIC, 3083, Australia
2.Kazusa DNA Research Institute, Kazusa Kamatri 2-6-7, Kisarazu, Chiba, 292-0818, Japan
3.Department of Agriculture and Food Western Australia, Locked Bag 4, Bentley Delivery Centre, Bentley, WA, 6983, Australia
4.School of Plant Biology, Faculty of Natural and Agricultural Sciences, The University of Western Australia, 35 Stirling Highway, Crawley, WA, 6009, Australia
5.Centre for Comparative Genomics, Murdoch University, 90 South Street, Murdoch, WA, 6150, Australia
Abstract This study reports on the construction of the first genetic maps of subterranean clover (Trifolium subterraneum L.), a diploid, inbreeding annual pasture legume, and alignment of its linkage groups with those of red clover (T. pratense L.) andMedicago truncatula Gaertn. Transferability of red and white clover (T. repens L.) simple sequence repeat (SSR) markers to subterranean clover was observed. A total of 343 SSR loci were mapped into eight subterranean clover linkage groups, with 6–31 loci per linkage group and 27 loci with similar locations between two distinctF2 mapping populations. Phenotypic data obtained for flowering time, content of three isoflavonoids (formononetin, genistein and biochanin A), hardseededness, leaf markings, calyx pigmentation and hairiness of stems were analyzed, together with genotypic data. Genomic intervals influencing each trait were assigned to one to three chromosome regions, accounting for 5.5–59.8% of the phenotypic variance. Syntenic relationships were observed among subterranean clover, red clover andMedicago truncatula genomes. Comparisons of loci shared between the three species indicated that at least two chromosomal regions have undergone duplications in the subterranean clover genome. Candidate genes for isoflavone content were identified usingM. truncatula as a reference genome. Synteny-based segmentation observed in Brassicaceae chromosomes helped to account for the apparent segmental-based relationship between the clover genomes, particularly within the subterranean clover lines. The proposed segmental nature of clover genome could account for the extensive variation observed between the parental genotypes, while not preventing production of fertile intercrosses.
1.Laboratory of Plant Resistance and Genetics, College of Resources and Environmental Sciences, South China Agricultural University, Guangzhou, 510642, China 2.Plant Protection Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou, 510640, Guangdong, China 3.Rice Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou, Guangdong, China
The deployment of broad-spectrum resistance genes is the most effective and economic means of controlling blast in rice. The cultivar Er-Ba-Zhan (EBZ) is a widely used donor of blast resistance in South China, with many cultivars derived from it displaying broad-spectrum resistance against blast. Mapping in a set of recombinant inbred lines bred from the cross between EBZ and the highly blast-susceptible cultivar Liangjiangxintuanheigu (LTH) identified in EBZ a blast resistance gene on each of chromosomes 1 (Pish), 6 (Pi2/Pi9) and 12 (Pita/Pita-2). The resistance spectrum and race specificity of the allele atPi2/Pi9 were both different from those present in other knownPi2/Pi9 carriers. Fine-scale mapping based on a large number of susceptible EBZ × LTH F2 and EBZ × LTH BC1F2 segregants placed the gene within a 53-kb segment, which includesPi2/Pi9. Sequence comparisons of the LRR motifs of the four functional NBS-LRR genes withinPi2/Pi9 revealed that the EBZ allele is distinct from other knownPi2/Pi9 alleles. As a result, the gene has been given the designationPi50(t).
1QTL Genomics Research Center, National Institute of Agrobiological Sciences, Kannondai 2-1-2, Tsukuba, Ibaraki 305-8602, Japan.2Mountainous Region Agricultural Research Institute, Aichi Agricultural Research Center, Inahasi, Toyota, Aichi 441-2513, Japan.3Bioproduction Sciences, Ishikawa Prefectural University, Suematsu 1-308, Nonoichi-machi, Ishikawa 921-8836, Japan.4Institute of the Society for Techno-Innovation of Agriculture, Forestry and Fisheries, Ippaizuka, Tsukuba, Ibaraki 305-0854,Japan.5Division of Plant Sciences, National Institute of Agrobiological Sciences, Kannondai 2-1-2, Tsukuba, Ibaraki 305-8602, Japan.6Division of Genome and Biodiversity Research, National Institute of Agrobiological Sciences, Kannondai 2-1-2, Tsukuba, Ibaraki305-8602, Japan.7Graduate School of Life and Environmental Sciences, University of Tsukuba, Tennohdai 1-1-1, Tsukuba, Ibaraki 305-8572, Japan. ↵*To whom correspondence should be addressed. E-mail:fukusan@affrc.go.jp
1 Plant Disease Resistance Research Unit, Division of Plant Sciences, National Institute of Agrobiological Sciences, Kannondai 2-1-2, Tsukuba, Ibaraki 305-8602, Japan 2 Field Crop Division, Aichi Agricultural Research Center, Nagakutecho, Aichi 480-1103, Japan 3 Institute of the Society for Techno-Innovation of Agriculture, Forestry and Fisheries, Ippaizuka, Tsukuba,Ibaraki 305-0854, Japan 4 National Agricultural Research Center for Hokkaido Region, National Agriculture and Food Research Organization, Hitsujigaoka, Sapporo, Hokkaido 062-8555, Japan * (fax +81 29 838 8383; e-mailtakatsuh@affrc.go.jp).
>
Rice blast is one of the most widespread and destructive plant diseases worldwide. Breeders have used disease resistance (R) genes that mediate fungal race-specific ‘gene-for-gene’ resistance to manage rice blast, but the resistance is prone to breakdown due to high pathogenic variability of blast fungus.Panicle blast 1 (Pb1) is a blast-resistance gene derived from theindica cultivar ‘Modan’.Pb1-mediated resistance, which is characterized by durability of resistance and adult/panicle blast resistance, has been introduced into elite varieties for commercial cultivation. We isolated thePb1 gene by map-based cloning. It encoded a coiled-coil–nucleotide-binding-site–leucine-rich repeat (CC–NBS–LRR) protein. The Pb1 protein sequence differed from previously reported R-proteins, particularly in the NBS domain, in which the P-loop was apparently absent and some other motifs were degenerated.Pb1 was located within one of tandemly repeated 60-kb units, which presumably arose through local genome duplication.Pb1 transcript levels increased during the development ofPb1+ cultivars; this expression pattern accounts for their adult/panicle resistance. Promoter:GUS analysis indicated that genome duplication played a crucial role in the generation ofPb1 by placing a promoter sequence upstream of its coding sequence, thereby conferring aPb1-characteristic expression pattern to a transcriptionally inactive ‘sleeping’ resistance gene. We discuss possible determinants for the durability ofPb1-mediated blast resistance.
1 Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China, 2 Rice Research Institute, Sichuan Agricultural University, Chengdu 611130, China 3 Institute of Crop Research, Chinese Academy of Agricultural Sciences, Beijing 100081, China, and 4 Graduate School of Chinese Academy of Sciences, Beijing 100101, China * (fax +86 28 8272 2918 or +86 10 6487 3428; e-maillishigui_sc@263.net orlhzhu@genetics.ac.cn).
>
Rice blast, caused by the fungal pathogenMagnaporthe grisea, is one of the most devastating diseases in rice worldwide. The dominant resistance gene,Pi-d2 [previously namedPi-d(t)2], present in the rice variety Digu, confers gene-for-gene resistance to the Chinese blast strain, ZB15.Pi-d2 was previously mapped close to the centromere of chromosome 6. In this study, thePi-d2 gene was isolated by a map-based cloning strategy.Pi-d2 encodes a receptor-like kinase protein with a predicted extracellular domain of a bulb-type mannose specific binding lectin (B-lectin) and an intracellular serine–threonine kinase domain.Pi-d2 is a single-copy gene that is constitutively expressed in the rice variety Digu. Transgenic plants carrying thePi-d2 transgene confer race-specific resistance to theM. grisea strain, ZB15. The Pi-d2 protein is plasma membrane localized. A single amino acid difference at position 441 of Pi-d2 distinguishes resistant and susceptible alleles of rice blast resistance genePi-d2. Because of its novel extracellular domain,Pi-d2 represents a new class of plant resistance genes.
Zhai C , Lin F , Dong Z , He X Y , Yuan B , Zeng X S , Wang L , Pan Q H
The rice–rice blast pathosystem is of great interest, not only because of the damaging potential of rice blast to the rice crop, but also because both the pathogen and its host are experimentally amenable. The rice blast resistance genePik, which is one of the five classical alleles located at thePik locus on the long arm of chromosome 11, confers high and stable resistance to many Chinese rice blast isolates. The isolation and functional characterization ofPik were performed in the present study through genetic and genomic approaches. A combination ofPik-1 andPik-2 is required for the expression ofPik resistance. BothPik-1 andPik-2 encode coiled-coil nucleotide binding site leucine-rich repeat (NBS-LRR) proteins, and each shares a very high level of protein identity with corresponding proteins encoded by thePik-m andPik-p alleles.Pik could be distinguished from otherPik alleles, includingPik-m andPik-p, by the allele-specific, single-nucleotide polymorphism T1-2944G. The coupled genes probably did not evolve as a result of a duplication event, and are far from any NBS-LRRR gene characterized.Pik is a younger allele at the locus that probably emerged after rice domestication.
Yuan B , Zhai C , Wang W , Zeng X S , Xu X K , Hu H Q , Lin F , Wang L , Pan Q H
The blast resistance genePik-p, mapping to thePik locus on the long arm of rice chromosome 11, was isolated by map-basedin silico cloning. Four NBS-LRR genes are present in the target region of cv. Nipponbare, and a presence/absence analysis in thePik-p carrier cv. K60 excluded two of these as candidates forPik-p. The other two candidates (KP3 andKP4) were expressed in cv. K60. A loss-of-function experiment by RNAi showed that bothKP3 andKP4 are required forPik-p function, while a gain-of-function experiment by complementation test revealed that neitherKP3 norKP4 on their own can impart resistance, but that resistance was expressed when both were introduced simultaneously. BothPikp-1 (KP3) andPikp-2 (KP4) encode coiled-coil NBS-LRR proteins and share, respectively, 95 and 99% peptide identity with the two alleles,Pikm1-TS andPikm2-TS. ThePikp-1 andPikp-2 sequences share only limited homology. Their sequence allowedPik-p to be distinguished fromPik,Pik-s,Pik-m andPik-h. BothPikp-1 andPikp-2 were constitutively expressed in cv. K60 and only marginally induced by blast infection.
1 Iwate Biotechnology Research Center, Kitakami, Iwate 024-0003, Japan 2 Tsukuba Botanical Garden, National Museum of Nature and Science, Tsukuba 305-0005, Japan 3 Iwate Agricultural Research Center, Kitakami, Iwate 024-0003, Japan 4 Graduate School of Agriculture, Hokkaido University, Sapporo 060-8589, Japan * (fax +81 197 68 3881; e-mailterauchi@ibrc.or.jp).
>
TheOryza sativa (rice) resistance genePia confers resistance to the blast fungusMagnaporthe oryzae carrying theAVR-Pia avirulence gene. To clonePia, we employed a multifaceted genomics approach. First, we selected 12R-gene analog (RGA) genes encoding nucleotide binding site-leucine rich repeats (NBS-LRRs) proteins from a region on chromosome 11 that shows linkage toPia. By using seven rice accessions, we examined the association between Pia phenotypes and DNA polymorphisms in the 10 genes, which revealed three genes (Os11gRGA3–Os11gRGA5) exhibiting a perfect association with the Pia phenotypes. We also screened ethyl methane sulfonate (EMS)-treated mutant lines of the rice cultivar ‘Sasanishiki’ harboringPia, and isolated two mutants that lost the Pia phenotype. DNA sequencing ofOs11gRGA3–Os11gRGA5 from the two mutant lines identified independent mutations of major effects inOs11gRGA4. The wild-type ‘Sasanishiki’ allele ofOs11gRGA4 (SasRGA4) complementedPia function in both mutants, suggesting thatSasRGA4 is necessary forPia function. However, when the rice cultivar ‘Himenomochi’ lackingPia was transfected withSasRGA4, the Pia phenotype was not recovered. An additional complementation study revealed that the two NBS-LRR-typeR genes,SasRGA4 andSasRGA5, that are located next to each other and oriented in the opposite direction are necessary forPia function. A population genetics analysis ofSasRGA4 andSasRGA5 suggests that the two genes are under long-term balancing selection.
1 Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, USA. 2 Present address: Laboratory of Plant Genetics and Breeding, Akita Prefectural University, Akita, Japan. 3 State Key Laboratory of Rice Biology, China National Rice Research Institute, Chinese Academy of Agricultural Sciences, Hangzhou, China. 4 Department of Plant Biology, Michigan State University, East Lansing, Michigan, USA. 5 Institute for Genomic Diversity, Cornell University, Ithaca, New York, USA. 6 National Center for Soybean Improvement, State Key Laboratory of Crop Genetics and Germplasm Enhancement, College of Agriculture, Nanjing Agricultural University, Nanjing, China. 7 US Department of Agriculture–Agricultural Research Service, Ithaca, New York, USA. 8 National Key Laboratory of Crop Genetic Improvement, National Center for Plant Gene Research (Wuhan), Huazhong Agricultural University, Wuhan, China. 9 National Center for Plant Gene Research, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China.
Dacheng Tian, Hitoshi Araki, Eli Stahl, Joy Bergelson, and Martin Kreitman*
Department of Ecology and Evolution, University of Chicago, 1101 East 57th Street, Chicago, IL 60615
Natural selection and genetic linkage cause DNA segments to have genealogical histories resembling those of the selected sites.When a polymorphism maintained by selection is old, it will have an island of enhanced sequence variability surrounding it,which represents a detectable “signature of selection.” We investigate the structure of single-nucleotide polymorphisms (SNPs)in a 20-kb interval containing theArabidopsis thaliana disease resistance geneRPS5, a locus containing common alleles for the presence/absence of the entire locus. The alleles are considerably diverged atsurrounding sites, indicative of an old polymorphism maintained by selection. The island of “enhanced” variability extendsseveral kilobases to either side of theRPS5 deletion junction, and these SNPs are in nearly complete linkage disequilibrium with theRPS5 insertion/deletion. At a distance of 10 kb to either side of the locus, however, we find low levels of polymorphism and theabsence of linkage disequilibrium between individual SNPs andRPS5 alleles. Our results show that the interval of enhanced variability surrounding this balanced polymorphism inArabidopsis is large enough to be readily detected, but small enough to span the focal gene and few others. For this species it shouldbe possible to identify the complete set of genes with long-lived polymorphisms, a potentially important subset of genes segregatingfor functional variants.
1 Institute of Plant Biology, University of Zürich, Zollikerstrasse 107, CH-8008 Zürich, Switzerland 2 Agroscope Reckenholz-Tänikon Research Station ART, Reckenholzstrasse 191, CH-8046 Zürich, Switzerland 3 Max Planck Institute for Informatics, Campus E1.4, D-66123 Saarbrücken, Germany * (fax +41 44 634 82 04; e-mailbkeller@botinst.uzh.ch).
>
Some plant resistance genes occur as allelic series, with each member conferring specific resistance against a subset of pathogen races. In wheat, there are 17 alleles of thePm3 gene. They encode nucleotide-binding (NB-ARC) and leucine-rich-repeat (LRR) domain proteins, which mediate resistance to distinct race spectra of powdery mildew. It is not known if specificities from different alleles can be combined to create resistance genes with broader specificity. Here, we used an approach based on avirulence analysis of pathogen populations to characterize the molecular basis ofPm3 recognition spectra. A large survey of mildew races for avirulence on thePm3 alleles revealed thatPm3a has a resistance spectrum that completely contains that ofPm3f, but also extends towards additional races. The same is true for thePm3b andPm3c gene pair. The molecular analysis of these allelic pairs revealed a role of the NB-ARC protein domain in the efficiency of effector-dependent resistance. Analysis of the wild-type and chimericPm3 alleles identified single residues in the C-terminal LRR motifs as the main determinant of allele specificity. Variable residues of the N-terminal LRRs are necessary, but not sufficient, to confer resistance specificity. Based on these data, we constructed a chimericPm3 gene by intragenic allele pyramiding ofPm3d andPm3e that showed the combined resistance specificity and, thus, a broader recognition spectrum compared with the parental alleles. Our findings support a model of stepwise evolution ofPm3 recognition specificities.
Plant–pathogen co-evolutionary selection processes are continuous, complex and occur across many spatial and temporal scales. Comprehensive studies of the flax–flax rust pathosystem have led to the postulation of the gene-for-gene model, a genetic paradigm describing recognition events between host disease resistance proteins and pathogen effector proteins. The identification of directly interacting fungal effector proteins and plant disease resistance proteins in this pathosystem has facilitated the study of both the physical nature of these interactions and the evolutionary forces that have resulted in a molecular arms race between these organisms. The flax–flax rust pathosystem has also been detailed on the scale of interacting populations, and the integration of molecular- and population-scale datasets represents a unique opportunity to further our understanding of many poorly understood facets of host–pathogen dynamics. In this article, we discuss recent developments and insights in the flax–flax rust pathosystem and their implications for both long-term co-evolutionary dynamics in natural settings, as well as short-term co-evolutionary dynamics in agro-ecosystems.
Amit Kumar Rai(1) Satya Pal Kumar(1) Santosh Kumar Gupta(1) Naveen Gautam(1) Nagendera Kumar Singh(1) Tilak Raj Sharma(1)
1.National Research Centre on Plant Biotechnology, Indian Agricultural Research Institute, New Delhi, 110012 India
Blast disease of rice, caused byMagnaporthe oryzae is an explosive disease that can spread rapidly in conducive conditions. R-gene mediated resistance offers an environmentally sustainable solution for management of this important disease of rice. We have earlier identified a unique R-gene of rice, on chromosome 11 ofOryza sativa ssp.indica cultivar Tetep. In this study we report functional validation of thePi-kh (Pi54) gene using complementation assay. The blast resistance candidate genePi-kh(Pi54) was cloned into a plant transformation vector and the construct was used to transform ajaponica cultivar of rice Taipei 309, which is susceptible toM. oryzae. Transgenic lines containingPi-kh(Pi54) gene were found to confer high degree of resistance to diverse isolates ofM. oryzae. The callose deposition was analyzed and compared between the transgenic and non-transgenic rice plants and widespread deposition was observed at the infection sites in plants showing incompatible interaction. Successful complementation ofPi-kh(Pi54) gene confirmed that the gene is responsible for resistance toM. oryzae in transgenic lines developed during this study. Expression analysis of the gene in resistant plants revealed that the gene is pathogen inducible in nature and is not expressed constitutively. Detection of callose deposition in resistant plants containingPi-kh(Pi54) gene implicates its involvement in the initiation of defense response cascade.
LIU Hai, XIAO Ying-Hui, TANG Wen-Bang, DENG Hua-Bing,CHEN Li-Yun*
刘海,肖应辉,唐文邦,邓化冰,陈立云*
Rice Research Institute, Hunan Agricultural University, Changsha 410128, China
Great success has been achieved in the utilization of two-line hybrid rice in China. Nevertheless, the seed production of two-line hybrid rice is still a key technology that limited the extension of two-line hybrid rice on large scale. Seed multiplication of the thermo-sensitive genic male sterile (TGMS) rice, which with a narrow range of temperature for fertility restoration, is hard to be success by using the most common multiplying technologies currently. In order to improve the yield stability and security in TGMS rice multiplication, we developed a practical computer selection system for choosing an optimal multiplying site. This system using the java language was able to select suitable multiplying sites with optimal ecological factors which meet the requirement of the fertility sensitive stage (FSS) and heading-flowering stage (HFS) for TGMS rice multiplication. By using this computer-aided selection system, 24, 29, 20, 21, and 22 ecological areas were selected ,which were superior to the most common seed producing area, Sanya of Hainan province, where the suitable probabilities of FSS and HFS are 35.0%, 42.5%, 55.0%, 65.0%, and 75.0% for the TGMS with the critical temperature of 22.0℃, 22.5℃, 23.0℃, 23.5℃, and 24.0℃, respectively, while the suitable probability of the optimal multiplying site selected by the computer-aided selection system were 83%, 93%, 100%, 100%, and 100% in both FSS and HFS for TGMS with different critical temperature, respectively, showing much higher values than those in Sanya, Hainan Province. In 2010, we tried to produce the seed of C815S, a TGMS rice with the critical temperature of 22.0℃ in Baoshan, Yunnan Province selected by the computer-aided selection system. The grain yield reached 8 437.5 kg ha-1, and created the highest yield record of seed multiplication of the TGMS rice. These results suggested that the problems of unstable yield and low security for the TGMS rice seed multiplication should be solved by this computer aided selection system, and we believe the optimal multiplying site selected by the computer-aided selection system will provide a useful technical support for healthy and sustainable development in the two-line hybrid rice.
为了解决水稻两用核不育系繁殖产量不高不稳、种子质量差、效益低的问题,利用全国740个气象站点50年的气象资料,应用计算机处理技术,开发了水稻两用核不育系繁殖基地计算机选择系统,并利用该系统筛选最适宜的水稻两用核不育系繁殖基地。该系统以同时满足不育系育性敏感安全期和抽穗扬花安全期光温条件为依据,采用Java语言编制而成。针对不育起点温度为22.0℃、22.5℃、23.0℃、23.5℃和24.0℃的水稻两用核不育系,分别筛选到24、29、20、21和22个安全系数优于海南三亚冬繁的基地,以上5种不育起点温度的水稻两用核不育系在最佳繁殖基地繁种成功概率分别可达83%、93%、100%、100%和100%。2010年,在应用本系统筛选到的云南保山繁种基地进行了不育起点温度为22.0℃的水稻两用核不育系C815S的繁殖,单产达8 437.5 kg hm-2,创造了我国水稻两用核不育系繁殖产量最高记录,证实采用该计算机系统选择两用不育系繁殖基地是有效的。因此,采用本系统筛选到的繁殖基地进行水稻两用核不育系繁殖,可望解决海南冬季繁殖风险大、产量不高、种子质量差以及冷水串灌繁殖产量不高不稳和效益低下的问题,值得推广应用。














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 , 邓元宝
, 邓元宝