



{kind=link}
{kind=link}
{kind=link}
{kind=link}
籼、粳超级稻品种根系形态及若干生理特征的差异
[龚金龙 , 邢志鹏, 胡雅杰, 张洪程
, 邢志鹏, 胡雅杰, 张洪程* , 戴其根, 霍中洋, 许轲, 魏海燕, 高辉, 郭保卫]
, 邢志鹏, 胡雅杰, 张洪程, 戴其根, 霍中洋, 许轲, 魏海燕, 高辉, 郭保卫]
|
|


第一作者联系方式: E-mail:gongjinlong0729@126.com
为阐明籼、粳超级稻根系干物质积累与分布特征和主要形态生理性状指标的差异及其与产量形成的关系, 以当地主体且具有代表性的2个超级杂交籼稻组合和2个常规粳型超级稻品种为试验材料, 对麦稻两熟制条件下籼、粳超级稻单茎和群体根干重、总根长、根数、根体积、根系吸收面积、发根力、抽穗后根系伤流强度及根冠比、每条根长、根直径、根密度、颖花根流量、抽穗期根系在土层中的分布和产量构成等方面进行了系统的比较研究。结果表明: (1)整个生育期, 粳稻的根冠比、每条根长、发根数、发根体积、发根干重及颖花根流量、穗数、群体颖花量、结实率和实收产量均高于籼稻, 而根直径、每穗粒数和千粒重低于籼稻, 其中根冠比、每条根长、颖花根流量、穗数、每穗粒数、结实率和实收产量差异达显著或极显著水平; (2)粳稻抽穗前(含抽穗期)单茎根干重、总根长、根数、根体积和根系总吸收表面积及根密度均低于籼稻, 但差异不显著, 而成熟期这6个指标粳稻均显著或极显著高于籼稻; (3)粳稻拔节前单茎活跃吸收表面积和活跃吸收表面积比均小于籼稻, 而拔节后(含拔节期)两者差异趋势与之相反, 差异显著或极显著; (4)除拔节期群体根干重、拔节期和抽穗期群体根数外, 其他群体形态生理特征指标粳稻均显著或极显著高于籼稻; (5)无论是单茎还是群体, 粳稻抽穗后0~35 d根系伤流量均显著或极显著高于籼稻; (6)粳稻0~10 cm土层根系干重所占比例极显著低于籼稻, 10 cm以下土层根系干重占根系总干重的比例极显著高于籼稻, 粳稻扎根更深, 进一步强化了植株抗倒防早衰能力。与超级杂交籼稻相比, 常规粳型超级稻抽穗后根系生长优势不断加大, 特别是群体生长优势, 成熟期粳稻所有根系形态生理特征指标均优于籼稻, 是粳稻高产形成的重要原因和保障。
, XING Zhi-Peng, HU Ya-Jie, ZHANG Hong-Cheng, DAI Qi-Gen, HUO Zhong-Yang, XU Ke, WEI Hai-Yan, GAO Hui, GUO Bao-Wei
The objective of this study was to reveal the difference of dry matter accumulation and its distribution, and the main morphological and physiological characteristics in roots between
植株根系既是水分和养分吸收的主要器官, 又是多种激素、有机酸和氨基酸合成的重要场所, 其形态和生理特性与地上部的生长发育、产量和品质的形成等均有密切的关系[ 1, 2, 3]。作为水稻科学的一个重要组成部分, 水稻根系研究也一直受到作物栽培耕作、遗传育种、营养生理、土壤肥料、生态环境和农田灌溉等相关学科的专家和学者的高度重视, 并在根系形态建成与研究方法、根系形态结构与生理活性和地上部株型及产量的关系、根系对环境胁迫的响应、根系遗传育种与生长调控等方面作了大量建设性工作[ 4, 5, 6]。但由于根系生长在地下, 受研究方法和技术手段的限制, 对其研究的深度和广度均滞后于地上部。关于水稻基因型间根系形态生理特征的差异, 前人也作了一些相关研究[ 7, 8, 9], 但基本上都以单个亚种为对象, 或比较研究亚种内杂交稻与常规稻的差异, 亦或两种类型品种仅用作试验重复, 而截至2013年农业部认定的101个超级稻品种中, 籼型超级稻以杂交稻为主, 粳型超级稻全部为常规稻。因此, 系统比较研究高产栽培条件下常规粳型超级稻品种与超级杂交籼稻组合根系形态生理特征的差异及其与高产形成的关系, 具有重要的理论意义与生产实践价值。
江苏地处长江下游经济发达地区, 属麦(油)稻两熟制的籼、粳兼作地区, 历史上曾几度开展“籼稻改粳稻”和“粳稻改籼稻”的制度改革和布局调整; 改革开放以后, 在全省农业行政推广和科研、教学等多单位的共同努力之下, 江苏粳稻生产取得重要进展, 从20世纪80年代初籼稻比例占85%以上发展到目前粳稻所占比例接近90%, 成为南方粳稻种植面积最大的主产省, 粳稻年种植面积、总产分别占全国的20%和30%左右; 2012年江苏粳稻年种植面积近200万公顷, 且多年来江苏水稻单产一直稳居8.25 t hm-2以上, 部分高产县(市、区)平均单产已超过9 t hm-2 [ 10, 11, 12]。其中, 江苏“籼改粳”工作的推进是以常规粳稻替代杂交籼稻并配套粳稻高产栽培技术而实现的, 但针对这两种类型水稻品种的产量差异及粳稻高产形成机制的研究尚不系统。因此, 本研究立足于苏中地区, 在麦稻两熟制籼、粳同季兼作条件下, 选用能安全齐穗成熟和充分利用当地温光资源的偏迟熟[ 13]高产当家籼、粳超级稻品种各2个, 配以各自相应的高产栽培管理措施, 以充分发挥其产量水平。在此基础上, 系统深入分析籼、粳超级稻根系形态生理特征的差异, 从根系层面上阐明粳稻高产形成机制, 以期为江苏以及同类生态稻区乃至整个南方稻区实施新一轮“籼改粳”提供理论依据和技术支撑。
扬州大学农学院校外试验基地江苏省兴化市钓鱼镇科研基地(33°05' N, 119°57' E)位于江淮之间, 江苏里下河腹部, 属北亚热带湿润气候区, 年平均温度15℃左右, 年降水量1024.8 mm左右, 全年日照时数2305.6 h左右, 无霜期227 d左右。地势低洼, 平均地面海拔仅2.4 m, 境内湖泊众多, 河流纵横, 属水网圩区。试验地前茬为小麦(实收产量6.1 t hm-2)。土壤地力中等, 勤泥土, 质地黏性, 含有机质22.3 g kg-1、全氮1.54 g kg-1、速效磷14.5 mg kg-1、速效钾111.9 mg kg-1, pH 7.2, 0~20 cm耕作层土壤容重1.29 g cm-3。
供试籼型超级杂交稻组合为中熟中籼稻扬两优6号和两优培九, 其全生育期在145~150 d之间; 常规粳型超级稻品种为早熟晚粳稻镇稻11和武运粳24, 其全生育期在160~165 d之间。扬两优6号、两优培九、镇稻11和武运粳24分别由江苏省里下河农业科学研究所、江苏省农业科学院粮食作物研究所、江苏省镇江市农业科学院、江苏省(武进)水稻研究所选育和提供。所有品种主茎总叶片数(N)均为18叶, 伸长节间数(n)均为6个。
在长江下游麦-稻两熟制条件下, 根据籼、粳超级稻高产特征和要求, 分别设置能充分发挥两种类型水稻品种产量潜力的密、肥、水等高产栽培管理措施。应用精确定量栽培原理设计, 2011年和2012年5月15日将催好芽的稻种用旱育保姆拌种后均匀撒播, 旱育壮秧, 6月15日移栽, 栽插密度25.1万穴 hm-2(13.3 cm × 30 cm), 常规粳稻每穴2苗, 杂交籼稻每穴1苗。杂交籼稻总施氮量225 kg hm-2, 常规粳稻300 kg hm-2, 氮肥运筹比例均为基蘖肥∶穗肥 = 5∶5, 基肥∶分蘖肥 = 6∶4, N∶P2O5∶K2O= 1.0∶0.5∶1.0。分蘖肥于移栽后一个叶龄一次性施入, 穗肥于倒四叶和倒三叶分两次施用, 磷肥全部做基肥, 钾肥分基肥和拔节肥两次等量施用。在有效分蘖临界叶龄的前一个叶龄, 当茎蘖数达到预期穗数的80%时, 自然断水搁田, 坚持轻搁、多次搁的原则; 拔节至成熟期实行湿润灌溉, 干干湿湿, 直至成熟收获前1周断水。其他栽培管理措施均按照高产栽培要求实施。
试验重复3次, 共12个小区, 随机区组排布, 每个小区33.3 m2。不同类型和品种间作埂隔离, 并用塑料薄膜覆盖埂体, 各处理间设约0.5 m间隔沟, 试验区四周设约1 m排灌沟, 保证各小区单独排灌。
移栽时, 采用改进的根系研究方法, 在每个小区预先埋入特制的圆形营养袋, 营养袋材料为黑色耐氧化聚乙烯膜, 口径18 cm, 高25 cm, 袋底和侧面有8个口径为0.5 cm的滤水透气孔。各品种每小区按S形共埋入32个营养袋(实际测定需要28个)。每个营养袋中间栽插1穴, 用于测定各处理关键生育时期单茎根系相关形态生理性状指标, 并根据茎蘖追踪和普查结果得到的群体茎蘖(穗)数计算群体根系性状的相关指标(单茎根系相关指标×群体茎蘖数或穗数)。
1.3.1 根系形态、根冠比和根系吸收表面积 分别于有效分蘖临界叶龄期(N-n)、拔节期、抽穗期和成熟期, 从每小区各取埋入土中的营养袋4个, 剥去袋体, 置40目尼龙网袋中用流水冲洗获得完整根系, 剪取所有根系, 其中2穴用于测定根长、根数、根粗等形态性状指标, 然后将鲜根及地上部稻株置恒温箱内, 105℃杀青30 min, 80℃烘干至恒重, 称取根系干重和地上部干重, 计算根冠比; 另外2穴用于测定根系体积、根系总吸收表面积和活跃吸收表面积。采用排水法测定根体积, 甲烯蓝蘸根法[ 14]测定根系吸收表面积。
1.3.2 发根力 分别于有效分蘖临界叶龄期、拔节期、抽穗期和成熟期按每小区平均茎蘖(穗)数取代表性植株3穴, 洗去根部土壤, 细心剪去所有不定根, 放入蒸馏水中培养10 d后测定发根数、发根体积和发根干重。
1.3.3 根系在土层中的分布 于抽穗期, 从每个小区各取埋入土中的营养袋3个, 剥去袋体, 从上至下将圆柱形土体切割为0~5、5~10、10~15、>15 cm四个层次, 分别置40目尼龙网袋中用流水冲洗干净, 105℃杀青30 min, 80℃烘干至恒重, 测定并计算各层根系干重及占总干重的比例。
1.3.4 根系伤流强度 分别于抽穗后0、7、14、21、28、35 d, 按每小区茎蘖数的平均值取代表性植株3穴, 于18:00在各茎离地面约12 cm处(在测定前排干田间水)剪去其上部植株, 然后将预先称重的脱脂棉放在茎的剪口处, 包上塑料薄膜, 于第2天早上8:00取回带有伤流液的脱脂棉并称重, 计算伤流量。
1.3.5 产量 成熟期每个小区取5个观察点, 每点取10穴, 共50穴测定穗数; 各小区取20穴装进塑料窗纱口袋内, 风干后, 脱粒、去杂质(不去空瘪粒), 计算20穴的总粒数, 进而求得每穗粒数; 用水漂法去除空瘪粒, 求结实率; 以1000实粒样本(干种子)称重, 重复3次(误差≤0.05 g)求取千粒重。以联合收割机实收产量, 测定水分, 去除杂质, 折算实产。
根密度(g cm-3) = 根干重/根体积;
根系活跃吸收表面积比(%) = 根系活跃吸收表面积/根系总吸收表面积×100%;
颖花根流量(mg h-1或mg g-1 h-1) = 抽穗后0~ 35 d基部节间平均伤流量/颖花(实粒或粒重)。
采用Microsoft Excel 2003计算数据并作图; 运用SPSS数据处理系统[ 15]统计分析, 采用LSD法多重比较数据。
2.1.1 根系干重和根冠比 根干重是根系生长状况的集中体现, 而根冠比是表征植株地下部和地上部是否协调生长的重要指标[ 16]。从表1可以看出, 随生育进程, 所有品种单茎根干重和群体根干重在抽穗期达到最大值, 成熟期有所下降; 而地上部干重, 无论是单茎还是群体, 均随着生育进程而不断增加, 根冠比则不断降低。两种类型品种之间, 籼稻有效分蘖临界叶龄期、拔节期和抽穗期单茎根干重均高于粳稻, 但差异不显著, 而成熟期粳稻单茎根干重则显著高于籼稻, 2011年和2012年差异幅度分别达47.11%和48.80%; 群体根干重方面, 除拔节期外, 其他生育时期均表现粳稻>籼稻的趋势, 其中抽穗期和成熟期差异达极显著水平, 这可能与拔节期籼稻无效分蘖多、分蘖结构不合理[ 17]有关。就2个品种平均值而言, 2011—2012年粳稻有效分蘖临界叶龄期、拔节期、抽穗期和成熟期根冠比分别为0.244、0.220、0.185、0.089、0.250、0.224、0.189和0.090, 分别较籼稻高5.90%、11.14%、25.77%、69.16%、5.95%、11.26%、26.12%和71.89%, 差异显著或极显著, 说明随水稻生长发育, 粳稻根系生长优势越来越明显。由此可见, 生育后期(抽穗至成熟期)较大的根系干重及根冠比是粳稻根系物质积累的重要特征之一。
2.1.2 抽穗期根系在土层中的分布 抽穗期根系 形态及分布是反映水稻生育中后期群体质量的重要指标之一, 它是水稻营养生长状况的集中表现, 并在一定程度上影响着水稻抽穗后光合物质生产与积累、矿质营养吸收与转移、籽粒灌浆与充实、抗倒支撑能力以及高产超高产的构建与形成[ 18]。由图1可以发现, 籼、粳超级稻抽穗期根系在土层中的分布明显不同。以2012年数据为例, 镇稻11和武运粳24在0~10 cm土层根系干重所占比例分别为69.64%和67.95%, 而扬两优6号和两优培九却高达77.31%和76.01%, 2个品种平均值籼稻高出粳稻7.86%。0~ 5 cm和5~10 cm土层表现的趋势与之一致(图1), 就其平均值而言, 粳稻抽穗期0~5 cm和5~10 cm土层根系干重占根系总干重的比例分别较籼稻高8.18%和16.94%。10 cm以下土层, 无论是10~15 cm, 还是>15 cm, 以及>10 cm, 均表现粳稻>籼稻的趋势, 粳稻10~15 cm和>15 cm土层2个品种根系干重所占比例的平均值分别较籼稻高7.91%和124.61%, 这可能也是粳稻氮素吸收能力和抗倒伏能力均强于籼稻的重要原因之一。籼、粳超级稻抽穗期各土层根系干重占根系总干重的比例的差异均达到显著或极显著水平, 且两年试验的重复性较好, 表现趋势一致(图1)。由此可见, 根系干重的优化分布及深扎根性是粳稻根系分布的重要特征之一。
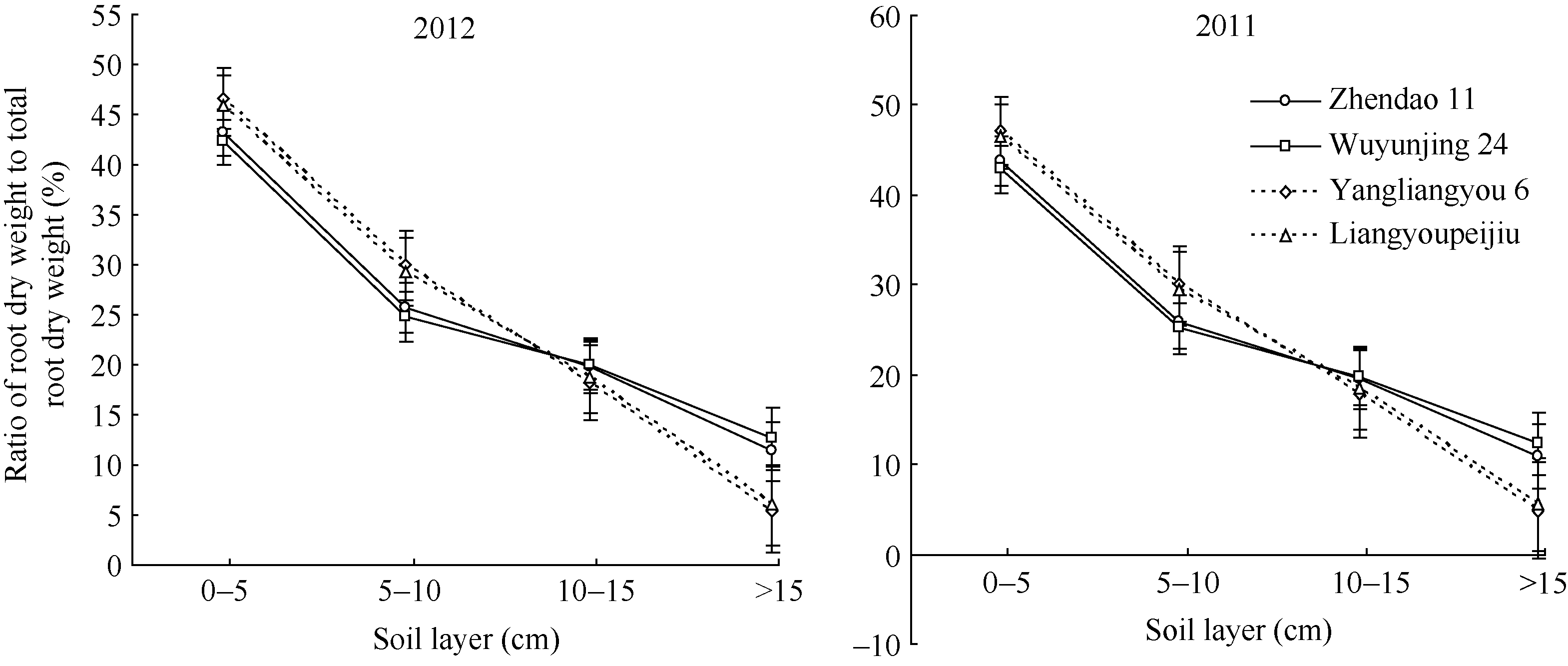 | 图1 籼、粳超级稻抽穗期各土层根系干重占根系总干重的比例的差异Fig. 1 Difference of ratio of root dry weight in each soil layers to total root dry weight at heading between indica and japonica super rice |
| 表1 籼、粳超级稻主要生育期根系干重和地上部干重及根冠比的差异 Table 1 Difference of dry weight of root and shoot at the main growth stages and their root-shoot ratios between indica and japonica super rice |
根长、根数、根直径及根体积和根密度等是衡量根系形态性状的几个重要指标[ 19]。表2表明, 随生育进程, 除扬两优6号和两优培九抽穗期群体根体积外, 所有品种的其他形态性状指标均在抽穗期达到最大值, 以后逐渐下降, 这可能与籼稻拔节后群体茎蘖消减迅速[ 17]有关。2012年镇稻11有效分蘖临界叶龄期、拔节期、抽穗期和成熟期每条根长分别为15.9、15.9、16.5和11.8 cm, 武运粳24则依次为16.0、16.0、17.4和12.0 cm, 粳稻2个品种每条根长的平均值分别较籼稻高25.54%、25.59%、25.75%和51.41%, 差异极显著, 这与粳稻10 cm以下土层根系干重所占比例极显著高于籼稻(图1)相一致。2011年籼、粳超级稻每条根长差异趋势与2012年一致(表2)。进一步分析单茎根系形态特征时可以发现(表2), 抽穗前粳稻单茎总根长、单茎根数和单茎根体积均小于籼稻, 而成熟期粳稻这3个指标均大于籼稻, 差异显著或极显著; 群体形态特征方面, 整个生育期粳稻群体总根长和群体根体积均显著或极显著大于籼稻, 而粳稻有效分蘖临界叶龄期和成熟期群体根数多于籼稻, 拔节期和抽穗期粳稻群体根数少于籼稻, 其中拔节期和成熟期群体根数达到极显著水平, 这可能与粳稻群体苗(穗)数多、籼稻生育中期(拔节至抽穗期)无效分蘖保持有一些小须根、但长度和体积都比较小[ 17]有关。无论哪个生育时期, 粳稻根直径都小于籼稻, 但差异不显著; 2011— 2012年镇稻11和武运粳24成熟期根密度分别为0.177、0.187、0.175和0.186 g cm-3, 均极显著高于籼稻, 而其他生育时期粳稻根密度均低于籼稻, 但两者差异均未达到显著水平, 说明粳稻根系不粗、充实不足, 还有待于强化粳稻根系育种。由此可见, 根系较长且群体生长优势明显, 尤其是生育后期(抽穗至成熟期), 是粳稻根系形态的重要特征之一。
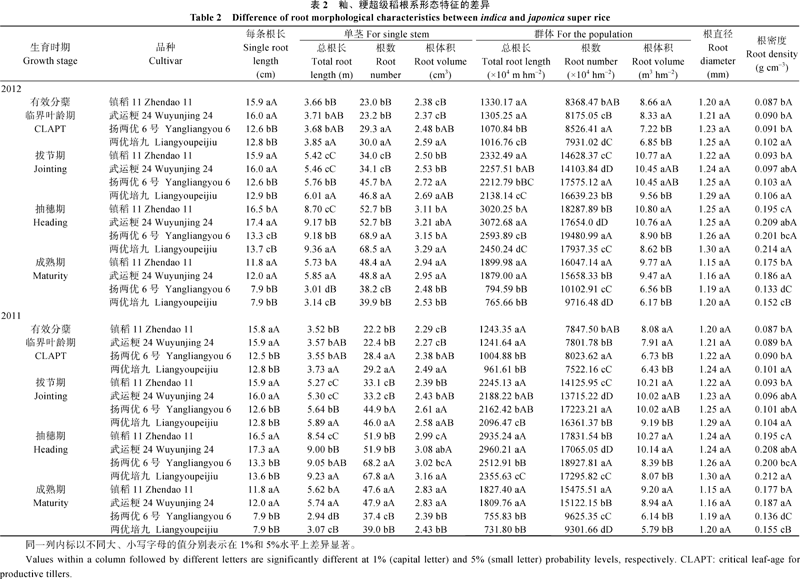 | 表2 籼、粳超级稻根系形态特征的差异Table 2 Difference of root morphological characteristic between indica and japonica super rise |
2.3.1 根系吸收面积 由表3可知, 籼、粳超级稻单茎和群体根系总吸收表面积和活跃吸收表面积及根系活跃吸收表面积比均随着生育进程呈先升后降的变化趋势, 抽穗期最大。粳稻抽穗前根系单茎总吸收表面积均小于籼稻, 但差异不显著; 成熟期, 2011—2012年镇稻11和武运粳24根系单茎总吸收表面积分别为1.19、1.25、1.21和1.27 m2, 均显著高于籼稻; 粳稻有效分蘖临界叶龄期根系单茎活跃吸收表面积略低于籼稻, 而拔节期粳稻根系单茎活跃吸收表面积略高于籼稻, 抽穗期和成熟期粳稻根系单茎活跃吸收表面积均极显著高于籼稻, 说明大田生长期随着水稻生长发育的持续, 粳稻根系单茎吸收优势越来越明显。群体根系吸收面积方面, 就2个品种平均值而言, 2012年粳稻有效分蘖临界叶龄期、拔节期、抽穗期和成熟期根系群体总吸收表面积分别为521.29、703.67、1326.64和404.87, 分别较籼稻高23.05%、8.77%、24.27%和48.14%, 群体活跃吸收表面积分别为210.19×104、342.28×104、795.28×104和123.44×104 m2 hm-2, 分别较籼稻高18.94%、22.39%、42.83%和209.07%, 差异极显著, 说明粳稻根系群体吸收优势更为突出。2011年籼、粳超级稻根系群体吸收特征所表现的趋势与2012年一致(表3)。粳稻有效分蘖临界叶龄期根系活跃吸收表面积比显著低于籼稻, 而拔节期、抽穗期和成熟期粳稻根系活跃吸收表面积比均极显著高于籼稻, 说明拔节后粳稻根系吸收活力更强, 有利于光合积累和物质代谢, 这与籼稻移栽后分蘖力较强、粳稻生育中后期(拔节至成熟期)生长势较强[ 17]是相一致的。由此可见, 群体吸收面积大、活跃吸收表面积比高, 生育中后期(拔节至成熟期)吸收能力强, 是粳稻根系吸收生理的重要特征之一。
 | 表3 籼、粳超级稻根系吸收表面积的差异Table 3 Difference of root absorbing surface between indica and japonica super rise |
2.3.2 发根力 发根力表征稻株萌发新根的潜能, 是根系及地上部植株生理生化代谢强弱的外在表现, 也是衡量植株健壮与否的重要标志之一[ 20]。从表4可以看出, 所有品种的发根数、发根体积和发根干重均随生育进程而呈先升后降的趋势, 抽穗期最大; 籼稻成熟期3个发根指标均为0, 说明籼稻生育后期根系活力趋弱, 有明显的早衰现象。两种类型品种之间, 整个生育期粳稻发根数均多于籼稻, 2011—2012年粳稻有效分蘖临界叶龄期、拔节期和抽穗期2个品种发根数的平均值分别为每穴7.1、12.5、18.8、7.2、12.8、19.2条, 分别较籼稻高5.38%、11.44%、18.69%、5.38%、11.44%、18.69%, 其中抽穗期发根数差异达到极显著水平, 说明与籼稻相比, 粳稻生理生化代谢及发根力的优势均越来越强。发根体积和发根干重所表现的趋势与发根数一致(表4)。由此可见, 较高的发根数量及质量是粳稻根系生理活性的另一重要特征。
| 表4 籼、粳超级稻发根力的差异 Table 4 Difference of root germination ability between indica and japonica super rice |
2.3.3 抽穗后根系伤流强度 根系伤流强度是反映植株根系活力的另一重要指标[ 21]。由表5可知, 籼、粳超级稻单茎和群体根系伤流量均随着抽穗灌浆的持续而不断降低。两种类型品种之间, 无论是镇稻11还是武运粳24, 粳稻抽穗后0、7、14、21、28和35 d单茎根系伤流量均显著或极显著高于籼稻; 就2个品种平均值而言, 2011—2012年粳稻抽穗后0~ 35 d根系单茎平均伤流量分别达83.53 mg h-1和86.09 mg h-1, 分别较籼稻高47.55%和46.34%, 差异极显著; 群体根系伤流量表现趋势与单茎根系伤流量一致(表5), 且抽穗后0~35 d群体根系伤流量及其平均值的差异均达到极显著水平。由此可见, 生育后期(抽穗至成熟期)较强的根系伤流强度是根系代谢生理的又一重要特征。从籼、粳超级稻颖花根流量的差异来分析(表6), 本试验采用颖花、实粒和粒重3个指标来表示颖花根流量, 2011年和2012年镇稻11三个参数值分别为0.627 mg h-1、0.678 mg h-1、26.218 mg g-1 h-1和0.639 mg h-1、0.693 mg h-1、26.511 mg g-1 h-1, 武运粳24三个参数值分别为0.624 mg h-1、0.689 mg h-1、25.599 mg g-1 h-1和0.639 mg h-1、0.710 mg h-1、26.266 mg g-1 h-1, 粳稻2年2个品种3个指标值的平均值分别较籼稻高79.92%、66.11%和64.53%, 差异极显著, 这在实收产量上也有较好的反映。就平均值而言, 2011年和2012年粳稻实收产量分别达10.84 t hm-2和11.12 t hm-2, 分别较籼稻高12.84%和11.79%, 差异显著。再从产量构成因素来看(表6), 粳稻穗数极显著高于籼稻, 每穗粒数极显著低于籼稻, 最终两年2个品种群体颖花量的平均值粳稻较籼稻高2.75%, 差异不显著; 库容充实性状上, 粳稻结实率极显著高于籼稻, 千粒重则略低于籼稻, 说明穗多、库足且籽粒灌浆充实量多是粳稻高产形成的重要特征与途径。
| 表5 籼、粳超级稻抽穗后根系伤流强度的差异 Table 5 Difference of root bleeding intensity after heading between indica and japonica super rice |
 | 表6 籼、粳超级稻颖花根流量及产量的差异Table 6 Difference of root bleeding per spikelet and grain yield between indica and japonica super rise |
有关根系形态结构与活性及其与产量的关系, 一直是水稻根系研究的热点, 也是争论的一个焦点[ 4]。其核心问题是高产水稻应具有怎样的根系形态生理特征?根量是不是越大越好?根系活力是不是越强越好?目前大多数研究均较为一致地认为[ 22, 23, 24], 根重、根数、根系吸收表面积、颖花根活量、根系伤流强度等与产量均呈显著或极显著的正相关; 但也有研究结果表明[ 25, 26, 27], 根量或根冠比过大会造成无效消耗而对产量产生不利影响, 并据此提出了“根系冗余生长[ 28]”的观点。笔者认为这不仅与供试品种、耕作制度、肥水管理措施等有关, 还受到大田生长条件下根系取样方法的影响。关于籼、粳亚种间根系性状的差异, 陈健[ 29]以籼型、粳型、偏籼型、偏粳型的18个水稻品种为主要试材, 采用盆栽与田间栽培相结合的方法, 研究发现籼型、偏籼型品种的根干重和根体积分别高于粳型、偏粳型; 籼粳交育成种根数有优势; 根活性吸收面积, 偏籼>籼, 偏粳>粳, 并与单株产量密切相关; 孕穗期单株伤流强度为偏粳>偏籼>籼>粳; α-萘胺氧化力品种间差异极显著。董桂春等[ 30]在群体水培条件下, 选用包括常规籼稻、常规粳稻、广亲和品种、三系杂交籼稻、杂交粳稻、两系法杂交稻在内的92个水稻品种(组合)表明, 多数根系形态性状及根系吸收面积均为两系法杂交稻>杂交籼稻>杂交粳稻>常规籼稻>常规粳稻>广亲和品种, 根系活性为三系杂交籼稻>杂交粳稻>常规籼稻>常规粳稻>两系法杂交稻>广亲和品种; 杂交水稻的每株根干重、每株不定根总长、每条不定根长、最长根长、每株根系活性、根系活跃吸收面积显著或极显著高于常规水稻, 单位长度根重则显著低于常规水稻; 籼型水稻的每株根干重、最长根长、每条不定根粗、每条不定根重、单位长度根重、每株根系活性、根系活跃吸收面积显著或极显著大于粳型水稻。代贵金等[ 31]采用盆栽的方式, 对北方不同类型水稻品种的根系特征比较发现, 与粳稻相比, 籼稻主要表现为根数较少, 根体积较大, 根粗长, 根系吸收面积和比表面积都较小, 根冠比小, 后期衰退早且快, 这可能与籼稻不耐低温、籽粒灌浆充实速度快等[ 17]有关。本研究采用改进的根系研究方法, 可更真切地反映根系生长情况。试验结果表明, 粳稻有效分蘖临界叶龄期、拔节期、抽穗期和成熟期根冠比、每条根长、发根数、发根体积、发根干重及颖花根流量、穗数、群体颖花量、结实率和实收产量均高于籼稻, 而整个生育期根直径以及每穗粒数和千粒重粳稻低于籼稻, 其中根冠比、每条根长、颖花根流量、穗数、每穗粒数、结实率和实收产量达到显著或极显著水平。可见粳稻根系性状总体好于籼稻, 这可能与粳稻产量构成合理、光合物质生产优势明显、冠层结构优化、群体质量较高等有关, 还有待进一步研究证实。
关于根系在土层中的分布与产量的关系, 凌启鸿等[ 32]研究发现, 上层根量(上部3个发根节位长出的根)与产量呈极显著正相关, 下层根量与产量的关系随着栽培条件的不同而改变, 这可能是因为上层根的发生与穗分化同步。郑景生等[ 33]以产量12 t hm-2左右的超高产水稻为对象研究表明, 各根层对超高产形成的贡献率, 0~5 cm土层的上层根占65%左右, 5~20 cm土层的下层根占35%左右, 这与“上层根”是水稻生殖生长期的主要功能根系的论点一致。而蔡昆争等[ 34]研究认为, 上层根(0~10 cm)质量与产量之间没有显著的相关, 而下层根质量(10 cm以下)与产量之间呈显著正相关, 相关系数达0.7258。Yoshida[ 35]调查了1081个水稻品种的株高、分蘖和根系生长的关系, 认为根系分布较深的品种, 植株较高、分蘖较少, 主茎及早分蘖节上的根较长且多。穗数型品种的根纤细并多分布于土壤表层, 穗重型品种根系较粗, 深层根比例大, 抽穗期下位根比重大且活性强; 多穗型品种比大穗型品种根干重大, 矮秆多穗型品种根较少而短, 高秆大穗型品种发根多而长, 根系氧化力与叶角呈正相关。本研究结果表明, 粳稻0~10 cm土层根系干重所占比例极显著低于籼稻, 10 cm以下土层根系干重占根系总干重的比例极显著高于籼稻, 粳稻根系分布更深、结构趋优, 这与吴伟明等[ 36]的研究结果相一致, 他认为与籼型水稻相比, 粳型水稻具有明显的深扎根性。至于籼、粳超级稻根系空间形态分布特征的差异及其与高产形成的关系还有待进一步研究。因此, 在栽培调控措施上应适当减少基蘖肥、中期断氮、增施穗粒肥和中期烤田后实行干湿交替灌溉, 促进根系深扎, 提高深层根比例、培育发达上层根和塑造优质根型; 同时, 注重理想型根系基因型的筛选和遗传改良, 控制根系数量, 增加根系分布深度和根系分枝, 是未来根系育种及水稻高产更高产乃至超高产的基础性条件。
在整个生育时期中, 水稻根系形态结构及生理活力是不断变化的; 从重量、数量和形态来说, 根系的生长在开花时最高, 而后根系活性不断降低并开始衰亡[ 37, 38]。本研究结果表明, 随生育进程, 除扬两优6号和两优培九抽穗期群体根体积外, 籼、粳超级稻其他根系形态生理指标均呈先升后降的变化趋势, 抽穗期表现最大; 而地上部干重, 无论是单茎还是群体, 均随着生育进程而不断增加, 这也是根冠比不断降低的重要原因之一。此外, 抽穗期及抽穗前粳稻单茎根干重、总根长、根数、根体积和根系总吸收表面积及根密度均低于籼稻, 但差异不显著, 而成熟期这6个指标粳稻均显著或极显著高于籼稻; 粳稻拔节前单茎活跃吸收表面积和活跃吸收表面积比均小于籼稻, 而拔节后(含拔节期)两者差异趋势与之相反, 差异显著或极显著。群体根系形态生理特征方面, 除拔节期群体根干重、拔节期和抽穗期群体根数外, 其他指标均粳稻显著或极显著高于籼稻。根系伤流强度, 无论是单茎还是群体, 粳稻抽穗后0~35 d根系伤流量均显著或极显著高于籼稻。可见随生育进程而不断加强的根系生长优势及群体生长优势是粳稻根系形态生理的重要特征, 而籼稻生育后期(抽穗至成熟期)根系活力衰减快、易早衰。至于籼、粳超级稻根系衰老的器官形态、生理生化、基因表达等不同水平上的代谢机理和叶片衰老特性的差异及其形成的影响因素(如温光水等气候因子和栽培管理措施)还有待进一步深化研究。因此, 种植籼稻时, 应注重采用肥水精确管理等栽培措施和遗传育种等改良手段, 塑造良好的根系构型, 提高植株抽穗后根系活力, 延缓根系衰老消亡速度, 并注重个体与群体的协调发展。目前生产实践上, 调控水稻生育后期(抽穗至成熟期)根系生长的措施主要有[ 39]: ①利用转基因技术培育抗早衰材料; ②湿润灌溉或间歇灌溉; ③依据群体叶色合理叶面追肥; ④坚持病虫害无公害化防治; ⑤因苗科学使用植物激素或生长调节剂。采用“喷施宝”、“九二零”等防早衰剂改善根系及地上部生理生化代谢环境, 促进根系分化、延缓根系衰老、优化根冠结构, 提高籽粒灌浆充实量和经济产量。
与超级杂交籼稻相比, 常规粳型超级稻抽穗后根系生长优势, 特别是群体生长优势不断加大, 成熟期所有根系形态生理指标均较优, 这些是粳稻保持较高矿质营养元素吸收速度及高产形成的重要保障。粳稻抽穗期群体根系干重大且扎根深, 进一步强化植株抗倒防早衰能力。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|