
{kind=link}
{kind=link}
对中国水稻骨干亲本评定方法的探索
[孙宗修**,, *  , 鄂志国
, 鄂志国** , 王磊, 朱德峰, 张玉屏, 胡国成, 刘文真, 付亚萍* ]
, 鄂志国]
|
|


** 同等贡献(Contributed equally to this work)
亲本是作物育种的基础, 骨干亲本指同一时期用于育种的亲本中在同样的世代内衍生的品种(系)数量较多, 种植面积较大的育种材料。确定骨干亲本不但能够了解育种历史和亲本的贡献, 而且对育种也有重要指导作用。本文分析了以往确定骨干亲本时存在的主要问题, 提出评定骨干亲本应该有一定的规范, 即对同时代育成品种(系)和同衍生世代量化比较。以水稻数据库中我国育成的2398份水稻品种(系)为依据, 按亚种分类, 育成年代分组, 以连续3代衍生的品种(系)数量及其种植面积为参数, 分析育成品种在后续育种研究中的应用状况, 在此基础上, 采用量化评选的方法, 评选出我国20世纪下半叶用于水稻育种的骨干亲本, 其中籼稻26个, 粳稻23个。本文以水稻为例提出了评选骨干亲本的原则和方法, 并对今后选择育种亲本和提高育种效率提出了一些建议。
, E Zhi-Guo
Parent is the basis of crop breeding, while backbone parent refers to the breeding material that derives many varieties (lines) and owns large planting area within the same generation of breeding parents in the same period. Determining backbone parents will not only enable us to learn breeding history and parental contribution, but also play an important role in guiding practical breeding. We try to establish an objective methodology of assessing backbone parents. This paper analyzed main problems in determining backbone parents in the past, and suggested that certain norm should be followed to assess backbone parents, that is to say, carrying out quantitative comparison of improved varieties (lines) in the same period and the same derivative generations. A total of 2398 rice varieties (lines) in Chinese rice database were classified based on subspecies and grouped according to breeding time. The quantities of varieties (lines) and planting areas for three generations continuously were used as parameters to analyze the application of improved varieties in the further breeding research. On this basis, 49 backbone parents, including 26 parents of
亲本是育种的基础, “骨干亲本”指不仅直接培育了众多大面积推广品种, 而且还由其衍生出许多具有广泛应用价值的育种亲本材料[ 1]。农作物的骨干亲本除本身具备优良性状之外, 还具有高配合力, 即易与其他亲本杂交育成优良品种[ 2]。黎裕等[ 3]认为骨干亲本有综合性状优良、一般配合力高、适应性广3个特点。确定骨干亲本不但能够了解育种历史和亲本的贡献, 而且对指导现实的育种也有重要作用。近年来许多学者在SSR标记[ 4]、蛋白质[ 5]、染色体上特异分子标记[ 6]和QTL[ 7, 8, 9]等方面对骨干亲本做了研究, 还探讨了骨干亲本的利用方式[ 10, 11], 邵国军等注意到骨干亲本的演替[ 12]。但是对骨干亲本的选定还缺少客观的标准。
目前在确定“骨干亲本”时存在四方面问题: (1)往往只注重衍生品种的数量, 忽视衍生品种的实际种植面积。例如将育成品种接近30个或以上的小麦育种亲本界定为小麦骨干亲本[ 2], 又如根据育成或衍生品种占同期育成品种的比例认为20世纪90年代前黑龙江水稻育成品种的骨干亲本是石狩白毛、农林11、虾夷, 20世纪90年代后育成品种的骨干亲本是藤系138、上育397、富士光[ 13]。(2)骨干亲本与系谱混淆, 没有衍生世代的限制, 骨干亲本被演绎成了重要的原始亲本[ 14]。(3)由于对骨干亲本定义不够完善, 忽视亲本育成年代的影响。(4)判定骨干亲本缺乏可量化的客观标准。我们认为, 评定育种的骨干亲本, 有点像接力赛, 需要同一条起跑线, 同一个终点, 同样的赛制和同样的记分标准。时代不同, 育种水平不同, 育种目标与农业政策乃至种植区划都有很大变化, 而且不同时代育成的亲本用于育种的时间跨度不同, 因此不应该将不同时代育成的品种放在一起评比。正因为如此, 评定骨干亲本应该有一定的规范, 即同时代育成的品种(系, 下文统一简化称为品种)和对同样的衍生世代进行量化的比较。本文尝试以中国水稻品种及其系谱数据库中我国育成的2398份水稻品种为依据, 按育成年代分组, 以连续3代衍生的品种数量及其种植面积为参数, 对育成品种在后续育种研究中的应用状况进行整体分析, 在此基础上, 采用量化评选的方法, 评选出我国20世纪下半叶用于水稻育种的骨干亲本, 并对今后选择育种亲本和提高育种效率提出一些建议。
中国水稻品种及其系谱数据库(http://www.rice-data.cn/variety/)由中国水稻研究所创建, 用于收录国内外水稻品种和不育系等材料, 包括品种的亲本来源、选育单位、特征特性、适种地区和推广应用情况等, 截至2013年10月31日, 累计收录水稻种质14 330份。本文从该库中挑选衍生了后代的我国自育成品种2398份, 不包括野生稻、转基因水稻、三系与两系杂交水稻及其不育系、育成年份不详以及2011年后育成的品种, 外引品种引进的年份往往不详, 也不列入正文的分析范围。
将上述品种按籼粳分类, 籼稻1452份, 粳稻946份。每一类按育成年代分组(其中20世纪30-50年代数量少, 归为一组, 记为50S)。各组中统计每一品种(记为G0)衍生的子代(G1)、孙代(G2)和曾孙代(G3)的个数与频次(例如, A品种衍生 m个G1代品种, 则A品种衍生G1代的个数为 m, 衍生的频次为1)及该品种各衍生世代的种植面积(例如, A品种衍生 n个G2代品种, 这 n个品种的合计种植面积即为A品种G2代的种植面积)。种植面积中不包括育成品种自身的种植面积, 恢复系直接配组的杂交水稻的种植面积记入下一代的种植面积。面积数据源自农业部全国农技推广服务中心历年(1983—2010)发布的全国农作物主要品种推广情况统计表, 部分引自《中国水稻品种及其系谱》[ 15]。
由于上述2398份材料并非都能衍生G2~G3代, 分组统计各个世代出现数据的总频次, 考虑到2001—2010年育成品种衍生出G3代还需要时间, 仅计算从20世纪50年代到90年代育成品种衍生出G3代的比例。然后逐代对衍生的个数及其种植面积各自排序。以各衍生世代的频次为基数, 选取衍生个数以及种植面积最多的Top10%的育成品种作进一步的分析, 如果末位有若干数据相同的材料, 则同时入选, 因此有的分组可能多于10%。
由于取样原则规定用于分析的材料均为衍生了G1代的品种, 故G1的个数(2398)与G0相同(表1), 然而G1并非都衍生出G2代, 同样并非所有G2代都能够衍生出G3代。不同年代的籼稻衍生G2代的比例平均为70%, 粳稻约为60%, 也就是说, 有30%~40%育成品种的G1代没有衍生出G2代, 而G0能够衍生出G3代的数量更少, 不同年代平均仅约30%左右。
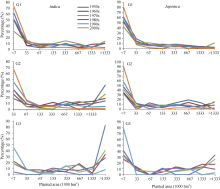
品种间衍生的G1、G2、G3代的个数差异大, 最多的是1968年育成的珍汕97[ 15]在G3代共衍生有995个品种, 其次为1980年育成的明恢63[ 16], 在G2与G3代分别衍生了709和722个品种。但同时也存在大量从上一代到下一代只衍生1个品种的现象, 在G1代尤其突出, 约46%的G0代品种在G1代只衍生1个品种。需要指出的是, 没有衍生G2代的绝大多数正是这些品种。图1显示各个年代G1衍生个数的分布频度, 可见随着衍生个数的增加其在当代所占的比例递减, 以20世纪80年代的籼稻为例, 只衍生1个品种的个数占全部G1代的40.1%, 衍生2~5个品种的个数占35.8%, 衍生6-10个品种的占12.1%, 衍生11~50个品种的占9.5%, 衍生50个以上品种的仅为2.6%, 籼粳稻年度间变化趋势基本一致。G2与G3代的变化趋势与G1代基本相同, 但是衍生多个品种的频度有所增加(图略)。
| 表1 衍生出的G1~G3代水稻品种(系)数及其占G0代的比例 Table 1 Quantity of G1-G3 generations derived from rice varieties (lines) (G0) and their respective ratios to G0 |
 | 图1 不同年代育成品种(系)衍生G1个数的分布频度Fig. 1 Distribution frequencies of G1 amount derived from bred varieties (lines) in different decades |
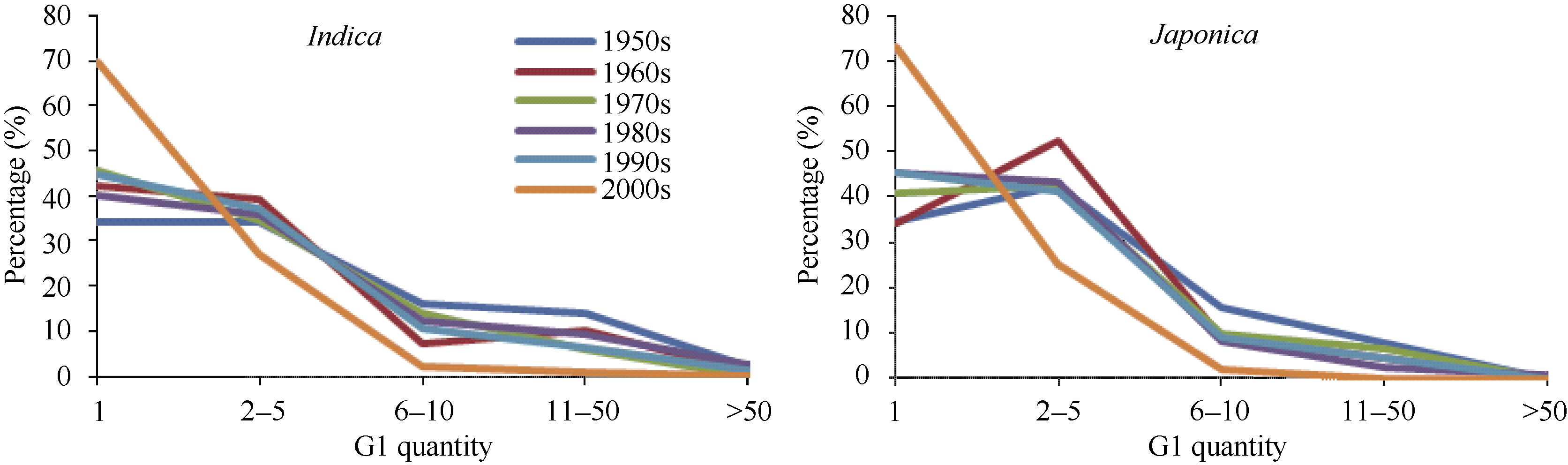
以单个育成品种为计算单位, 统计了各世代衍生品种合计种植面积。由图2可见, G1代衍生品种合计种植面积小于6600公顷的占当代总数的50%左右。另一方面, G1代种植面积较大的占当代总数比例不高, 从20世纪50年代到21世纪00年代, 籼稻平均有16.3%的G1代种植面积超过66万公顷, 粳稻仅为9.7%, 使分布图呈典型的L型。随着衍生世代的推移, 种植面积小于6600公顷的比例逐步减少, 而大于66万公顷的比例增加, 面积介于两者之间的变动不大。例如, G3代种植面积小于6600公顷的占当代的比例下降到40%以下(个别除外), 而超过132万公顷的品种数, 籼稻与粳稻分别平均达38.9%与17.9%, 结果使种植面积的L型图逐渐演变成为明显的U型图。
后代种植面积少于6600公顷的品种往往与上代只衍生1个后代的现象有关, 但是也有例外, 即并非衍生个数少的品种种植面积一定很少, 衍生多个品种也有合计种植面积少于6600公顷的现象。究其原因, 一是衍生的为多个不育系, 因此没有面积; 二是衍生的品种数虽多, 但是每个品种的种植面积都小于6600公顷, 而农业部统计方法是单个品种种植面积不到6600公顷的不列入统计, 所以也没有面积。
综上所述, 所分析的2398份育成品种中, 40%只衍生1个G1代, 约50%衍生的G1代种植面积在6600公顷以下, 两者大多数重叠表现。约2/3的籼稻与粳稻能够衍生G2代。各世代中衍生品种数达到或者超过50个以及种植面积超过132万公顷的比例很低。育成年代对后代衍生的数量影响不大。
分组统计3代(G1~G3)的衍生品种数与种植面积(共6个参数)的Top10%, 有时接近末尾的几个品种的数据相同而同时入选, 因此会出现入选个数超过10%的现象。由于入选Top10%品种中, 只有极少数所有6个参数能够全部达标, 有的只有1项符合Top10%的要求, 因此实际入选的品种数超过育成品种总数的10%, 为方便计, 入选品种依然通称为总数Top10%。统计上述各组含1、2直到6个Top10%数据的品种数, 由于基数不同, 计算了相应的百分比以资比较。结果表明达标1项Top10%数据的比例籼稻为45%左右, 粳稻为60%左右, 随着达标项数增加, 所占比例逐渐减少, 同时达标4~6项Top10%数据的品种约占实际入选品种的15% (籼稻)与10% (粳稻), 同时达标5~6项Top10%数据的品种约占4% (籼稻)与8% (粳稻)(表3)。
 | 图2 一定种植面积的衍生品种(系)数占当代品种总数的百分比Fig. 2 Distribution frequencies of derivational quantity from bred varieties (lines) for three generations with different amount of planted area in different decades |
| 表2 20世纪90年代籼稻和粳稻3个世代各参数总数Top10%的理论与实际入选数 Table 2 Theoretical and actual quantities of Top10% percent of each parameter from three generations of indica and japonica in 1990 |
进一步分析入选Top10%的育成品种在3个世代中的实际分布状况(表4), 仅在G1代拥有Top10%数据而在G2与G3代没有数据列入前10%的品种约占全部入选品种的50%左右, 年代之间差异不大; 仅在G2代拥有Top10%数据的约占15%~20%; 只在G3代含有Top10%数据的比例明显减少, 不少年代降为0。同时含2代Top10%数据的约占10%~20% (粳稻波动大些), 其中绝大多数为含G1和G2代的品种, 含G2和G3明显减少, 同时含G1和G3的更少。同时含3代的约占15% (粳稻波动大些), 且随着年代推移比例逐步下降, 这可能与第三代的衍生以及推广需要时间有关。
| 表3 满足Top10%品种(系)要求的参数数量的比例 Table 3 Percentages of parameter quantity met the requirement of Top10% varieties (lines) (%) |
| 表4 入选Top10%育成品种(系)的衍生后代在3个世代中的分布 Table 4 Distribution of Top10% varieties (lines) in three generations |
需要指出的是表4中的G1、G2与G3都可能存在2种情况, 即同一世代内衍生个数与种植面积两者都入选Top10%或者只有其中1个参数满足入选要求, 因此, 同时含2代的品种可能有2~4个参数同时达标, 同时含3代的品种可能有3~6个参数达标。同样, 表3中除了同时含有5~6个Top10%数据的品种肯定在3个世代都有分布外, 有2~4个Top10%数据的品种可能分布在1~3个世代。由表5可见, 不同年代育成品种中, 含有3个及以上满足Top10%要求的约占育成品种总数的5%~6%, 而含有4个及以上满足Top10%要求的锐减一半, 只有3%左右; 另一方面, 籼稻与粳稻3个世代都出现达标数据的品种分别为2.2%和3.2%, 与含有4个及以上满足Top10%要求的比例(3.3%和3.0%)接近, 而且大多数是重叠的, 将2项合并, 入选籼稻与粳稻占育成品种数的比例分别为3.5%与3.8%, 我们认为这些品种应该是上世纪下半叶我国水稻的骨干亲本。表6列出在此范围的26个籼稻与23个粳稻品种各代的衍生个数及种植面积, 这49个品种都是生产上发挥重大作用的品种或者杂交水稻亲本, 例如1980年育成的籼型恢复系明恢63[ 16], 不但由其直接配组的杂交稻(汕优63等)累计种植面积超过8400万公顷, 而且其衍生的G2和G3的个数和种植面积都居同组之首。但是, 同样在水稻生产上作出重大贡献的一些常规籼稻, 如广陆矮4号、特青与浙辐802等只能进入5%的范围, 未能入围3.5%。
由于衍生代数不够, 未对2000年代的育成品种评选骨干亲本。
| 表5 有3~4项参数及3个世代入选Top10%的育成品种(系) Table 5 Quantity of varieties (lines) with three to four parameters and three generations of selected Top10% |
育种的基础是亲本, 我国一贯重视亲本在水稻育种中的作用, 并提出术语“骨干亲本”。评定育种的骨干亲本离不开系谱, 我国对水稻育成品种的系谱研究相当深入, 除了研究报告, 还有专著[ 15, 17]并建立了较为完整的数据库[ 18]。这些研究重点分析了一些重要种质资源衍生的后代情况, 对当前生产上推广的水稻品种正本溯源起了积极作用。但是骨干亲本是指同一时期用于育种的亲本中在同样的世代内衍生的品种数量较多、种植面积较大的育种材料, 因此骨干亲本不同于系谱中的“共同祖先”。例如, 矮仔占在矮化育种的关键时期发挥了巨大作用[ 19], 毫无疑问是当时重要的骨干亲本(本研究也证实这一
点), 但是时至今日, 几乎所有亲本都带有 sd1基因, 谁也不会再用原始材料矮仔占作育种亲本, 因此认为它还是当今的骨干亲本显然不符合事实。同时, 遴选骨干亲本时界定衍生代数相当重要, 以石狩白毛为例, 该品种从上世纪40年代引进东北, 1956年开始作为亲本选育新品种, 到2010年, 共衍生6代115个品种, 但如果截止到第3代, 则为62个品种[ 1]。因此, 如果对20世纪90年代的亲本与上世纪50、60年代的亲本简单类比衍生的后代数, 由于前者比后者“年轻”30~40年, 衍生后代数量没有后者多而被否认也有失公允。正因为如此, 需要强调评定骨干亲本应该有一定的规范, 即同时代育成的品种和对同样的衍生代数量化比较。刘化龙等[ 13]将时间分为2段(1950—1990和1990—2010)遴选骨干亲本, 时间跨度太长。本研究将数据库中2398份材料按籼粳稻分类后, 按年代(10年)和3个衍生世代分组, 以衍生后代个数及其种植面积为参数, 从中选出TOP10%的品种, 最终选出至少有3个世代或者有4个参数同时符合Top10%的育成品种, 这些品种约占总数的3.5%左右, 符合骨干亲本应该是“同一时期内用于育种的亲本中能够在连续3代中衍生较多的品种并有较大种植面积”的设想。本研究之所以界定连续3代(G1~G3)作为统计的范围是考虑到新品种的选育一般历时4年(8代), 有一定种植面积需要3年(以上), 3代共计历时21年, 对于评定亲本的历史贡献是比较合适的。
| 表6 对我国水稻育种作出重大贡献的部分品种(系)(面积单位为千公顷) Table 6 Some varieties (lines) contributing significantly to the rice breeding in China (Area: 1000 hm2) |
| (续表6) 对我国水稻育种作出重大贡献的部分品种(系)(面积单位为千公顷) Table 6 Some varieties (lines) contributing significantly to the rice breeding in China (Area: 1000 hm2) |
利用农业部全国农技推广服务中心历年(1983— 2010)发布的全国农作物主要品种推广情况统计表, 我们量化分析了我国水稻的品种更替(待发表), 并根据本文提出的方法(年代除外)测算了各更替年代(1983—1987和1988—1997)育成品种中的骨干亲本(1998—2002因缺少足够的G3数据, 无法测算)。结果表明有25个品种入选为骨干亲本, 表6中在1983—1997年间育成并入选的骨干亲本都是测算中的骨干亲本, 此外, 还有3个1988年左右育成的籼稻常规品种(浙江嘉兴农业科学院的嘉育293, 湖南衡阳农业科学研究所的To974与四川省农业科学院作物所的CDR22)是表6中没有的, 究其原因是按品种更替年代划分时, 1980年代初育成的明恢63、测64-7、协青早与II-32B等不在1983—1987年的统计范围内, 从而为嘉育293提供了上升的空间。比较这两种按不同年代遴选骨干亲本的结果说明, 两者的符合率达90%左右, 从而说明本文提出的方法是可靠的。按理说, 采用品种更替的年代遴选骨干亲本更切合实际, 但是由于1983年前没有逐年的面积数据, 因此1950—1982的品种更替无法量化分析, 故本文还是采用以年代为基础遴选骨干亲本。
本研究评出26个籼稻与23个粳稻品种为上世纪50年代以来我国育成品种中的骨干亲本, 这些品种本身绝大多数是耳熟能详, 在生产中发挥了重要作用的, 有的则是颇有特色的材料(如广亲和材料02428[ 20])。有意思的是籼稻骨干亲本中大多数是杂交水稻的恢复系或者保持系, 常规品种很少, 尤其是20世纪70年代杂交水稻取得突破后, 除了广东农业科学院水稻研究所育成的丰矮占1号外, 同样在水稻生产中做出重大贡献的一些常规籼稻, 如广陆矮4号、特青与浙辐802等只能进入5%的范围, 未能入围3.5%的骨干亲本行列。究其原因, 一方面是杂交稻的恢复系和保持系竞争能力强, 其配合力也略强于这些常规籼稻, 上文中以品种更替为划分年代依据的测试结果也印证了这一观点, 另一方面, 种子公司热衷于发展杂交稻以获取更多利润可能也有重要影响。
根据我国稻作区划[ 17]对表6中49个骨干亲本的育成地归类(表7)表明, 有30个骨干亲本(13籼与17粳)出自长江中下游稻区, 其次为华南稻区(9籼), 西南和东北稻区各占5个。这一结果与我国各稻区的水稻种植面积、稻区生态与发展趋势是一致的。
我国水稻的生产和发展与国外品种的引进与利用有密切关系[ 1, 21], 我们的分析还发现外引品种对我国水稻骨干亲本的形成也至关重要。例如, 矮仔占4号的亲本矮仔占是广西容县的地方品种, 有资料表明矮仔占最先是由华侨从东南亚传入的[ 19], 辽粳5号的亲本之一是日本粳稻丰锦[ 15], 合江20、辽粳326和松粳3号等也都有日本粳稻的血缘, 至于籼型杂交稻的恢复系则全部与外引品种有缘。由于外引品种在我国的应用时间往往不详, 未能参与本文对骨干亲本的统一评定, 但也进行了类似国内品种的初步测试。结果表明籼稻IR8、IR30、圭630、IR24、IR36、IR26和密阳46以及粳稻农垦58、藤坂5号、丰锦、秋光、屉锦、喜峰、下北、金南风和Lemont等在我国水稻品种的选育中作出了重要贡献, 例如IR30的G1~G3代累计衍生1206个品种, 种植面积超过17 500万公顷, 农垦58不仅衍生大量常规品种, 而且衍生出光温敏雄性不育系农垦58S, 开创了我国两系杂交水稻的先河[ 22]。因此, 今后应当继续充分重视外引品种的开发和利用。
| 表7 表7 我国各稻区产生的骨干亲本 Table 7 Backbone parents bred from different rice regions in China |
对我国水稻育成品种在品种选育中作用的分析发现一个耐人寻味的现象, 即2398份材料中有40%在G1代只衍生了一个后代, 有50%在G1代的种植面积小于6600公顷而不能列入农业部的统计表, 同时还有30%~40%的材料只衍生了G1代, 没有G2代与G3代。结合我们对1983—2010年农业部发布的全国农作物主要品种推广情况统计表的水稻部分的分析, 在列入统计的2968个水稻品种(包括杂交水稻)中, 有1437份(48.4%)只有1~2次达到或者超过6600公顷, 犹如昙花一现(结果另行发表)。四川省对近10年育成品种的分析也发现, 只配1个组合的恢复系与不育系分别占78.6%和54.9%[ 23]。这些数据明白无误地反映出目前水稻育种的效率亟待提高。本研究表明, 虽然所有育成品种都可以作为育种亲本, 但是, 能够成为骨干亲本的往往是在水稻生产中作出重大贡献的品种。诚然, 也有一些生产上有很大面积的品种(如广陆矮4号等)由于配合力或品质等原因, 未能入围骨干亲本, 但其衍生后代的数量与种植面积远大于其他品种。因此精心选择亲本, 聚焦于生产上大面积应用的品种, 可能是明智的选择。当然, 充分利用骨干亲本的同时还需要根据各地的生态条件恰当选择与之配对的其他亲本, 既符合具体的育种目标又保持遗传多样性。
由表6可见, 省级农业科学院与地市级农业科学院(所)既是大品种的育成单位, 也是培育骨干亲本的主体, 这与两系杂交水稻选育单位的情况基本一致[ 24], 因此重视和加强对省级和地市级育种单位的支持十分重要。
探讨了评选我国农作物骨干亲本的方法, 提出应该在对同时代育成品种和同衍生世代量化比较基础上遴选骨干亲本。就水稻而言, 以连续3代衍生的品种数量及其种植面积为参数量化评选是可行和合理的。在不同稻区评选当地的骨干亲本需要考虑当地的生态条件和主栽品种等实际情况。文中提出的评选原则对其他作物骨干亲本的评选也有一定的参考价值。
致谢: 中国水稻研究所图书馆许杰先生提供许多宝贵文献资料, 特此致谢。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|