{kind=link}
{kind=link}
小麦新品种“山农20”抗病基因的分子检测
[李继发1, * , 邓志英1, * , 孙福来2 , 关西贞1 , 王延训1 , 田纪春1, *  ]
]
]
|
|


山农20是2011年和2012年分别通过国家黄淮南、北片审定的小麦高产多抗新品种, 在国家区试抗病性鉴定和生产中都表现出良好的抗黄淮麦区主要病害的特性。本研究利用与小麦抗白粉病、条锈病、叶锈病、纹枯病基因和抗赤霉病主效QTL紧密连锁的SSR、SCAR、STS等标记对该品种进行了分子检测, 发现山农20含有6个抗白粉病基因(
Shannong 20 is a newly released winter wheat variety in the National Regional Trials for North and South Yellow-Huai River Valleys, which exhibits excellent resistance to major diseases of wheat. In this study, the possible resistance genes in Shannong 20 were identified by using SSR, SCAR, and STS markers tightly linked to resistance genes/loci against powdery mildew, stripe rust, leaf rust, sheath blight, and scab. A total of 15 resistance genes were detected, including six (
我国从20世纪50年代开始进行小麦抗病常规育种, 主要针对条锈病。近年来, 除条锈病外, 许多原来的次要病害逐渐成为限制小麦生产的主要病害, 如白粉病、赤霉病、纹枯病和全蚀病等, 其中最主要的病害是条锈病, 其次是白粉病。由于小麦条锈病和白粉病分布广泛、病原菌生理小种复杂多变等特点, 常常导致品种抗性频繁丧失, 因此, 这些病害的防治任务十分艰巨。除了利用现有抗病基因进行常规育种外, 挖掘新抗病基因和在同一品种中聚合多种抗病基因是重要的育种策略。另外, 随着分子标记的广泛应用和标记技术的发展, 获得了越来越多的抗病基因分子标记, 为分子标记辅助选择(MAS), 加快目标基因的聚合育种提供了便利。
目前, 已有60多个小麦抗白粉病基因被定位到47个位点( Pm1~ Pm47), 分布在除3A、4D以外的各条染色体上[ 1, 2]。其中, Pm38和 Pm39为白粉病成株抗性基因[ 3], 其余抗性基因均为病原菌生理小种专化性的主效基因; 由于生理小种的不断变异, 大部分已丧失抗性[ 4]。
国际上已命名51个抗条锈病主效基因, 涉及48个位点( Yr1~ Yr48), 除 Yr11~Yr14外均被定位于特定的染色体上[ 5]。其中, Yr3位点有 Yr3a、 Yr3b和 Yr3c三个复等位基因, Yr4位点有 Yr4a和 Yr4b两个复等位基因, Yr11、 Yr12、 Yr13、 Yr14、 Yr16、 Yr18、 Yr29、 Yr30、 Yr36、 Yr39、 Yr46和 Yr48为成株抗性基因, 其他39个为苗期抗性基因[ 5]。还有72个条锈病成株抗性QTL被定位在除1A、1D、3D和7A染色体外的其余各个染色体上[ 4]。
目前已正式命名的小麦抗叶锈病基因有68个, 其中大多数为小种特异的主效抗叶锈病基因[ 6], 已报道39个分子标记与抗叶锈病基因连锁或共分离[ 7]。
近年来, 小麦纹枯病在我国特别是黄淮麦区趋向严重, 已上升为我国小麦主要病害之一, 但关于抗纹枯病的遗传研究报道较少。张小村等[ 8]利用2个重组自交系群体RIL-8和RIL-SES, 对抗纹枯病性QTL进行分析, 检测到1个SSR标记(Xgwm526)与纹枯病抗性具有关联性, 与纹枯病抗病基因 Ses1的遗传距离为27.9 cM。
小麦赤霉病主要发生在温暖潮湿和半潮湿地区, 在我国主要发生在长江中下游冬麦区和东北春麦区, 近年来随着全球气候变暖、灌溉条件的改善及耕作制度的变化, 有进一步蔓延到黄淮麦区的趋势[ 9]。目前, 除7D染色体外的其他所有的染色体上都检测到抗赤霉病QTL的存在[ 10], 其中位于3B、5A和6B染色体上的QTL在苏麦3号、望水白、Wuhan、Nyubai、Frontana、CM82036和DH181中被检测到, 并且效应较大, 是小麦抗赤霉病的主效QTL。但到目前为止, 世界上还没有发现对赤霉病完全免疫的小麦品种。
尽管已获得一些主要抗病性的QTL及其分子标记, 但在实际生产中应用却很少。目前生产上应用的小麦品种基本上都是常规育种方法培育而成, 由于其抗病机制及其携带的抗病基因都不明确, 致使很难进一步研究和利用很多优良品种。因此, 利用分子标记分析重要品种的抗病基因, 不仅为优异抗病基因的鉴定和利用MAS技术进行多基因聚合提供技术基础, 而且有助于跟踪重要或骨干亲本所含抗病基因的来源和遗传, 为在育种中有效利用不同抗病基因及其合理布局奠定基础。
本课题组培育的山农20属于高产多抗小麦新品种, 其亲本为PH82-2-2和954072。2010年和2011年分别通过国家黄淮南片和黄淮北片试验程序, 是目前很少见的广适性“双国审”小麦新品种。该品种抗逆性好、抗病性强、品质优、高产、稳产、适应性强, 适宜在包括苏北、皖北、河南、山西、陕西、山东、河北7个省份和地区的黄淮麦区推广种植。在近几年的推广种植中, 通过田间接种鉴定, 该品种表现出对多种小麦主要病害的优良抗性, 但是其抗病机制尚不清楚。本研究针对小麦抗白粉病、抗条锈病、抗叶锈病、抗纹枯病和抗赤霉病等相关性状, 选取已开发的与有关抗性基因或主效QTL连锁距离相对较近和稳定的SSR、STS、SCAR等分子标记, 对山农20及其亲本进行抗病性和抗病基因的分子鉴定, 旨在明确该品种抗病基因及其抗性机制, 为对其进一步研究和利用奠定基础, 也为利用MAS方法培育具抗性和稳产小麦新品种提供参考。
山农20及其亲本PH82-2-2 (母本)和954072 (父本)均由本课题组保存。
2008—2010年, 由中国农业科学院植物保护研究所根据全国农业技术推广服务中心《农技种函[2008]301号》文件要求, 分别对国家冬小麦品种区域试验品种进行人工接种抗病性鉴定, 其中供试条锈菌、叶锈菌、白粉菌均为混合优势小种, 赤霉菌和纹枯病菌以强致病力菌株接种。条锈病鉴定菌系包括CY29、CY30、CY31、CY32等生理小种; 叶锈病接种菌系以PHT、THT、PHK生理小种为主, 同时混有THP和FHT; 白粉病菌菌系包括V1、V3a、V3b、V3c、V3d、V3e、V3f、V4a、V4b、V5、V6、V7、V8、V17、V19、V25和V35; 赤霉菌菌株为F15、F34、F0301和F0609; 纹枯病菌菌株为R0301。各年度均按《小麦抗病虫性评价技术规范》(NY/T 1443-2007)进行接种和抗病性鉴定, 在4月上中旬分别进行田间和室内接种, 待对照品种充分发病后调查病情。
为了更准确地鉴定山农20在生产种植中的抗病性, 2010—2013年分别在山东的济宁、泰安、菏泽, 安徽的宿州、濉溪、阜阳, 河南的商丘、郑州、濮阳及江苏的徐州、新沂、连云港安排了0.67 hm2生产示范种植, 并按照《小麦白粉病测报调查规范》(NY/T 613-2002)、《小麦条锈病测报技术规范》 (GB/T 15795-2011)、《小麦叶锈病测报调查规范》(NY/T 617-2002)、《小麦纹枯病测报调查规范》(NY/T 614-2002)和《小麦赤霉病测报技术规范》(GB/T 15796-2011)调查田间病害, 反应型分为免疫(0)、高抗(1)、中抗(2)、中感(3)和高感(4)。每年每省选3点鉴定抗病性, 按抗性最弱点的抗病表现记载结果。
采用改良CTAB法(http://www.triticarte.com.au/)提取山农20及其两个亲本的基因组DNA, 用1%琼脂糖凝胶电泳检测DNA的浓度和质量。选用与小麦抗白粉病、抗条锈病、抗叶锈病、抗纹枯病和抗赤霉病基因或QTL紧密连锁或共分离的47个分子标记(见附表)进行分子鉴定。所有标记引物均由英潍捷基(上海)贸易有限公司合成。
PCR体系为20 μL, 含10×buffer (含Mg2+) 2 μL、dNTP 1.6 μL (10 mmol L-1)、每条引物0.4 μL、模板DNA 50 ng、1 U Taq聚合酶。PCR扩增程序为 95℃预变性5 min; 95℃变性50 s; 退火1 min (退火温度因具体引物而定), 72℃延伸1 min, 35个循环; 72℃延伸
国家区试中, 山农20对5种病害的抗病性在黄淮南片和北片表现不尽相同, 年度间也有差异, 但总体呈现对白粉、条锈和叶锈病有良好抗性, 多为免疫或高抗、或慢锈病, 而对赤霉病和纹枯病抗性较低, 甚至高感(表1)。
连续3个生长季在山东、安徽、河南和江苏田间自然感病条件下, 对山农20进行抗病性鉴定。总体来看, 白粉病由免疫到轻度发生; 条锈和叶锈病由免疫至高抗; 纹枯病由中抗至中感; 赤霉病发病较重, 表现为中抗至高感。2012年病害发生较重, 赤霉病表现最重, 其次是纹枯病。山东的抗病表现最好, 安徽、河南和江苏抗病性依次减弱(表2)。
| 表1 山农20在国家区试中的抗病性鉴定结果 Table 1 Resistance evaluation of Shannong 20 to main diseases of wheat in China national region trials |
| 表2 自然感病条件下山农20对5种主要病害的抗病性鉴定 Table 2 Resistance of Shannong 20 to five major wheat diseases under natural disease fields |
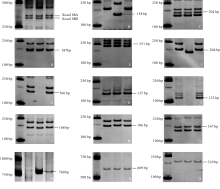
2.2.1 抗白粉病基因 利用与抗白粉病基因紧密连锁的分子标记, 对15个基因位点检测, 结果山农20在6个位点( Pm12、 Pm24、 Pm30、 Pm31、 Pm35和 Pm36)有扩增产物(图1), 且这些特异带在其一个亲本或双亲中也被检测到(表3), 说明山农20可能携带这6个抗白粉病基因。在 Pm16位点, 仅PH82-2-2有特异扩增, 而在山农20和954072中未检测到该片段(表3), 可能 Pm16基因未能从PH82-2-2传递到山农20。而 Pm36基因位点则是通过亲本954072传递给山农20的; Pm12、 Pm24、 Pm30、 Pm31、 Pm35基因位点在2个亲本和山农20中都含有; 其余8个基因位点在2个亲本及山农20中都不存在。
2.2.2 抗条锈病基因 利用与抗条锈病基因紧密连锁的分子标记, 对12个基因位点检测, 结果在6个位点( Yr5、Yr9、Yr15、Yr24、Yr26和 YrTp1)有扩增产物(图1), 且这些特异带在其一个亲本或双亲中均被检测到(表4), 说明山农20可能携带这6个抗条锈病基因。其中, Yr5和 Yr15基因位点由亲本954072传给山农20; Yr9、Yr24、Yr26和 YrTp1基因位点在2个亲本和山农20都含有, 而在2个亲本和山农20中都未检测到其余6个基因位点的特异片段。
 | 图1 不同抗病基因分子标记的扩增结果各泳道自左向右依次为DL2000分子量标记、山农20、PH82-2-2和954072。A~F: 抗白粉病基因 Pm12(A)、 Pm16(B)、 Pm24(C)、 Pm30(D)、 Pm35(E)和 Pm36(F)的分子标记带型; G~L: 抗条锈病基因 Yr5(G)、 Yr9(H)、 Yr15(I)、 Yr24(J)、 Yr26(K)和 YrTp1(L)的分子标记带型; M~O: 抗叶锈病基因 Lr1(M)、 Lr21(N)和 Lr26(O)的分子标记带型。Fig. 1 PCR profiles of molecular markers for different resistance genesFrom left to right, the four lanes in each panel were loaded with DL2000 marker and genome DNA templates extracted from Shannong 20, PH82-2-2, and 954072, respectively. A-F: Banding patterns of markers for powdery mildew resistance genes Pm12(A), Pm16(B), Pm24(C), Pm30(D), Pm35(E), and Pm36(F); G-L: Banding patterns of markers for stripe rust resistance genes Yr5(G), Yr9(H), Yr15(I), Yr24(J), Yr26(K), and YrTp1(L); M-O: Banding patterns of markers for leaf rust resistance genes Lr1(M), Lr21(N), and Lr26(O). |
| 表3 抗白粉病基因分子检测结果 Table 3 Molecular detection results of powdery mildew resistance genes |
| 表4 抗条锈病基因分子检测结果 Table 4 Molecular detection results of stripe rust resistance genes |
2.2.3 抗叶锈病基因 利用与抗叶锈病基因紧密连锁的分子标记, 对12个基因位点检测, 结果在2个位点( Lr21和 Lr26)有扩增产物(图1), 且这些特异带在双亲中也被检测到(表5), 说明山农20可能携带这2个抗叶锈病基因。对于 Lr1基因位点只在PH82-2-2扩增出特异性片段(见附图), 但未传递给山农20。在山农20和2个亲本均未扩增出其余9个
基因位点的特异性片段。
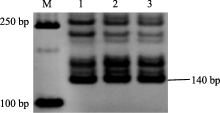
2.2.4 抗纹枯病基因 对于与抗纹枯病基因Ses1紧密连锁的SSR标记Xgwm526, 在山农20及其双亲中均扩增出约140 bp的特异性片段(图2), 推测山农20可能含有 Ses1基因。
| 表5 抗叶锈病基因分子检测结果 Table 5 Molecular detection results of leaf rust resistance genes |
 | 图2 Ses1基因的分子标记在山农20及其亲本中的扩增结果M: DL2000; 1: 山农20; 2: PH82-2-2; 3: 954072。Fig. 2 Banding patterns on Ses1locus in Shannong 20 and its parents amplified with specific primersM: DL2000; 1: Shannong 20; 2: PH82-2-2; 3: 954072. |
2.2.5 抗赤霉病主效QTL 利用5个已开发SSR标记, 对山农20、PH82-2-2和954072进行抗赤霉病QTL检测, 结果均未扩增出相应标记的特异性片段, 说明山农20及其亲本没有这些主效QTL。
小麦品种抗病性鉴定最经典的方法是使用抗病菌株的田间鉴定后进行基因推导, 但这一方法易受外界环境条件、遗传背景及基因×环境和基因×基因互作的影响, 而且有些基因没有相对应的致病菌株, 因此, 这一方法在检测某些新的高抗品种时, 难以确定是否含有新的抗病基因。利用与抗病基因/QTL紧密连锁的分子标记, 可以弥补基因推导的不足, 而且随着标记开发, 可及时鉴定和检测新的抗病基因及其遗传传递。山农20是一个农艺性状优良的冬小麦新品种, 除产量和品质好以外, 其突出的特点是对多种小麦主要病害表现广谱抗病性, 在多年的国家区试鉴定和生产鉴定中, 表现对白粉病和条锈病免疫、抗叶锈病、中感纹枯病和赤霉病。利用抗病性鉴定和分子标记检测方法, 明确山农20可能携带的抗病基因, 将对该品种的推广应用和选育抗病性更好的新品种有重要意义。
目前, Pm12、 Pm13、 Pm16、 Pm20、 Pm21、 Pm30在中国对小麦白粉病表现高抗至免疫, 而 Pm1、 Pm2、 Pm3、 Pm4a、 Pm4b、 Pm5、 Pm6、 Pm7、 Pm8等已丧失抗性, 只是在聚合状态下有较好的抗病性[ 11]。在已知的抗白粉病基因中, 来自簇毛麦的 Pm21抗性强, 其载体品种无明显的不良性状, 在不同小麦遗传背景下抗病性均表现稳定, 也是目前最有效的抗病基因之一[ 12]。 Pm22、 Pm25、 Pm26、 Pm27、 Pm28、 Pm29、 Pm31、 Pm32、 Pm33、 Pm34、 Pm35、 Pm36、 Pm37、 Pm38、 Pm39、 Pm40、 Pm41、 Pm42和 Pm43在中国的利用价值有待进一步鉴定[ 11]。本研究发现, 山农20可扩增出 Pm12、 Pm24、 Pm30、 Pm31、 Pm35和 Pm36等6个抗病基因的特异片段, 多个抗病基因的聚合可能是其对多个白粉病混合菌株免疫的重要原因。
从抗性基因在当前生产品种中出现的频率看, Yr9所占比例最大, Yr2和 Yr1次之[ 13]。李在峰等[ 14]发现 Yr5、 Yr10、 Yr15、 Yr26和 Yr31对当前流行的条锈病生理小种具有良好的抗性, 并认为 Yr16、 Yr18和 Yr29为条锈病持久抗性基因。殷学贵等[ 15]在小麦与十倍体长穗偃麦草的杂交后代材料A-3中鉴定出抗条锈病新基因 YrTp1, 对条锈病有较好抗性。本研究检测结果显示, 山农20可能聚合了 Yr5、 Yr9、 Yr15、 Yr24、 Yr26和 YrTp1基因, 田间抗条锈病鉴定结果暗示这些抗病基因的聚合可使品种表现出对条锈病免疫或高抗。
我国小麦品种资源中主要利用的抗叶锈基因有 Lr1、 Lr10、 Lr26、 Lr28、 Lr34和 Lr35, 其中 Lr1、 Lr10和 Lr26因病原菌新致病小种的产生或者次要小种上升为流行小种而“丧失”抗叶锈性[ 7], 但Kolmer[ 16]和German等[ 17]发现, 将 Lr1、 Lr3、 Lr11、 Lr26等基因与 Lr13或 Lr34聚合后, 可提高小麦成株期的抗病性, 抗病性一般都大于基因单独存在时。 Lr34为慢锈基因, 在成株期表达, 表现持久抗性, 同时兼抗条锈、秆锈和白粉病[ 18]。 Lr9、 Lr19、 Lr24和 Lr38具有全生育期抗病性, 但可能是典型的垂直抗病性基因, 应慎重应用[ 19]。本研究检测结果显示, 山农20可能携带 Lr21和 Lr26。在国家黄淮南片区试鉴定中, 山农20连续两年表现慢叶锈病, 在黄淮北片区试鉴定中表现高抗和中抗叶锈病; 在多年多点的生产种植鉴定中, 抗病反应为免疫和高抗。这是抗性基因聚合的效应还是存在新的抗(慢)叶锈基因, 有待深入研究。
关于小麦纹枯病抗性基因定位和分子标记的报道较少。张小村等[ 8]利用118个RIL-SES系, 发现1个与纹枯病抗性连锁的SSR标记, 即Xgwm526。该标记在山农20及其双亲中有相同的扩增带型, 结合田间纹枯病抗病性鉴定的结果, 推测山农20可能含有 Ses1基因。
小麦的赤霉病抗性是数量性状, 已在3B、4B和5A染色体上定位出抗赤霉病的主效QTL[ 20, 21]。本研究利用这些QTL的双侧标记对山农20及其2个亲本进行检测, 均未扩增出相应标记的特异片段。田间的抗病性鉴定显示山农20对赤霉病表现中感, 个别环境下为高感。因此, 在对山农20进行抗病性遗传改良时, 应着重考虑改善赤霉病抗性。
结合分子标记检测已知抗病基因/QTL和田间鉴定结果, 我们初步推测了山农20可能携带的抗病基因, 为解析该品种的优异抗性的遗传基础提供了初步资料。鉴于本试验田间抗病鉴定采用混合接种方法, 并且分子检测没有用原始载体品种作对照, 因而尚不能完全肯定候选基因的存在, 并且抗性鉴定结果是否完全是基因聚合的效应, 需作进一步验证或设计新的试验进行深入剖析。
初步推测山农20可能聚合了 Pm12、 Pm24、 Pm30、 Pm31、 Pm35和 Pm36共6个抗白粉病基因, Yr5、 Yr9、 Yr15、 Yr24、 Yr26和 YrTp1共6个抗条锈病基因, Lr21和 Lr26两个抗叶锈病基因, 以及抗纹枯病基因 Ses1; 未检测到抗赤霉病的主效QTL。该结果从某种程度上部分解释了山农20的优良抗病性。
附表 用于抗病性基因检测的分子标记 Supplementary Table Molecular markers used for the marker-assisted selection of disease resistance genes
| 基因/QTL Gene/QTL | 标记类型 Marker type | 引物 Primer | 距离 Distance (cM) | 引物序列 Primer sequence (5'-3') | 退火温度 Annealing temp. (℃) | 片段大小 Fragment size (bp) | 参考文献 Reference |
|---|---|---|---|---|---|---|---|
| 抗白粉病 Powdery mildew resistance | |||||||
| Pm2 | STS | Whs350-F | — | AGCTGTTTGGGTACAAGGTG | 60 | 498 | [22] |
| Whs350-R | GCCATCGTTTTCTACTAG | ||||||
| Pm8 | STS | IAG95-F | — | AGCAACCAAACACACCCATC | 60 | 1050 | [23] |
| IAG95-F | ATACTACGAACACACACCCC | ||||||
| Pm12 | SSR | Xcau108-F | co-segregating | CTGATCGTCCGTCTCTGTCC | 62 | [24] | |
| Xcau108-R | GGGTGAAACAGAATGTGGCG | ||||||
| Pm13 | STS | Xutv14-F | — | CGCCAGCCAATTATCTCCATGA | 62 | 564 | [25] |
| Xutv14-R | AGCCATGCGCGGTGTCATGTGAA | ||||||
| Pm16 | SSR | Xbarc004-F | 2.2 | GCGTGTTTGTGTCTGCGTTCTA | 52 | 158 | [26] |
| Xbarc004-R | CACCACACATGCCACCTTCTTT | ||||||
| Pm21 | SCAR | Pm21D | — | GTTTGTTCACGTTGAATGAATTC | 55 | 1265 | [27] |
| Pm21E | CACTCTCCTCCACTAACAGAGG | ||||||
| Pm24 | SSR | Xgwm337-F | 2.4±1.2 | CCTCTTCCTCCCTCACTTAGC | 55 | 204 | [28] |
| Xgwm337-R | TGCTAACTGGCCTTTGCC | ||||||
| Pm30 | SSR | Xgwm159-F | 5.7 | GCAGAAGCTTGTTGGTAGGC | 60 | 187 | [2] |
| Xgwm159-R | GGGCCAACACTGGAACAC | ||||||
| Pm31 | SSR | Xpsp3071-F | 2.5 | CGTGCCCTACACCTCCTTTTCTCTC | 60 | 153 | [3] |
| Xpsp3071-R | TCCGTACATACTCCGGGAGACC | ||||||
| Pm32 | SSR | Xgwm408-F | 1.3 | GTATAATTCGTTCACAGCACGC | 56 | 187 | [29] |
| Xgwm408-R | TCGATTTATTTGGGCCACTG | ||||||
| Pm33 | SSR | Xwmc317-F | 1.1 | TGCTAGCAATGCTCCGGGTAAC | 60 | 132 | [30] |
| Xwmc317-R | TCACGAAACCTTTTCCTCCTCC | ||||||
| Pm34 | SSR | Xbarc144-F | 2.6 | GCGTTTTAGGTGGACGACATAGATAGA | 62 | 235 | [31] |
| Xbarc144-R | GCGCCACGGGCATTTCTCATAC | ||||||
| Pm35 | SSR | Xcfd7-F | 10.3 | AGCTACCAGCCTAGCAGCAG | 60 | 251 | [32] |
| Xcfd7-R | TCAGACACGTCTCCTGACAAA | ||||||
| Pm36 | SSR | Xwmc75-F | 10.0 | GTCCGCCGCACACATCTTACTA | 61 | 206 | [33] |
| Xwmc75-R | GTTTGATCCTGCGACTCCCTTG | ||||||
| Pm43 | SSR | Xbarc11-F | 4.2 | GCGATGCGTGTAAAGTCTGAAGATGA | 62 | 299 | [34] |
| Xbarc11-R | GCGTCCATGGAGCTCTGTTTTATCTGA | ||||||
(续附表)
| 基因/QTL Gene/QTL | 标记类型 Marker type | 引物 Primer | 距离 Distance (cM) | 引物序列 Primer sequence (5'-3') | 退火温度 Annealing temp. (℃) | 片段大小 Fragment size (bp) | 参考文献 Reference |
|---|---|---|---|---|---|---|---|
| 抗条锈病 Stripe rust resistance | |||||||
| Yr2 | SSR | Xwmc364-F | 5.6 | ATCACAATGCTGGCCCTAAAAC | 58 | 207 | [35] |
| Xwmc364-R | CAGTGCCAAAATGTCGAAAGTC | ||||||
| Yr5 | SSR | Xgwm501-F | 10.5 | GGCTATCTCTGGCGCTAAAA | 57 | 166 | [36] |
| Xgwm501-R | TCCACAAACAAGTAGCGCC | ||||||
| Yr9 | SSR | Xgwm582-F | 3.7 | AAGCACTACGAAAATATGAC | 51 | 135 | [37] |
| Xgwm582-R | TCTTAAGGGGTGTTATCATA | ||||||
| Yr10 | SCAR | SC-200-F | — | CTGCAGAGTGACATCATACA | 60 | 200 | [38] |
| SC-200-R | TCGAACTAGTAGATGCTGGC | ||||||
| Yr15 | SSR | Xgwm33-F | 5.0 | GGAGTCACACTTGTTTGTGCA | 60 | 123 | [39] |
| Xgwm33-R | CACTGCACACCTAACTACCTGC | ||||||
| Yr17 | SCAR | SC-372-F | 0.8±0.7 | ATGTCCGCCCTTCCACAACTC | 60 | 372 | [40] |
| SC-372-R | CACTTGCCTATAAGCACAGAG | ||||||
| Yr18 | STS | csLV34-F | — | GTTGGTTAAGACTGGTGATGG | 57 | 150 | [41] |
| csLV34-R | TGCTTGCTATTGCTGAATAGT | ||||||
| Yr24 | SSR | Xgwm273-F | 6.1 | AGCAGTGAGGAAGGGGATC | 58 | 168 | [42] |
| Xgwm273-R | ATTGGACGGACAGATGCTTT | ||||||
| Yr26 | SSR | Xgwm18-F | 1.9 | TGGCGCCATGATTGCATTATCTTC | 59 | 186 | [43] |
| Xgwm18-R | GGTTGCTGAAGAACCTTATTTAGG | ||||||
| Yr36 | SSR | Xbarc101-F | co-segregating | GCTCCTCTCACGATCACGCAAAG | 62 | 125 | [44] |
| Xbarc101-R | GCGAGTCGATCACACTATGAGCCAATG | ||||||
| YrV23 | SSR | Xwmc356-F | 9.4 | GCCGTTGCCCAATGTAGAAG | 60 | 251 | [45] |
| Xwmc356-R | CCAGAGAAACTCGCCGTGTC | ||||||
| YrTp1 | SSR | Xwmc477-F | 0.4 | CGTCGAAAACCGTACACTCTCC | 61 | 167 | [46] |
| Xwmc477-R | GCGAAACAGAATAGCCCTGATG | ||||||
| 抗叶锈病 Leaf rust resistance | |||||||
| Lr1 | STS | WR003-F | — | GGGACAGAGACCTTGGTGGA | 55 | 760 | [47] |
| WR003-R | GACGATGATGATTTGCTGCTGG | ||||||
| Lr9 | STS | J13/1 | — | TCCTTTTATTCCGCACGCCGG | 66 | 1100 | [48] |
| J13/2 | CCACACTACCCCAAAGAGACG | ||||||
| Lr10 | STS | Lrk10D1 | — | GAAGCCCTTCGTCTCATCTG | 58 | 282 | [49] |
| Lrk10D2 | TTGATTCATTGCAGATGAGATCACG | ||||||
| Lr19 | STS | Lr19Gb-F | — | CATCCTTGGGGACCTC | 56 | 130 | [50] |
| Lr19Gb-R | CCAGCTCGCATACATCCA | ||||||
| Lr20 | STS | STS638-F | — | ACAGCGATGAAGCAATGAAA | 60 | 542 | [51] |
| STS638-R | GTCCAGTTGGTTGATGGAAT | ||||||
| Lr21 | STS | Lr21-F | — | CGCTTTTACCGAGATTGGTC | 57 | 669 | [52] |
| Lr21-R | TCTGGTATCTCACGAAGCCTT | ||||||
| Lr24 | STS | J09/1 | — | TCTAGTCTGTACATGGGGGC | 57 | 350 | [53] |
| J09/2 | TGGCACATGAACTCCATACG | ||||||
| Lr26 | STS | Lr26-F | — | GCAAGTAAGCAGCTTGATTTAGC | 60 | 210 | [54] |
| Lr26-R | AATGGATGTCCCGGTGAGTGG | ||||||
| Lr29 | SCAR | OPY10/1 | — | GTGACCTCAGGCAATGCA | 60 | 850 | [55] |
| OPY10/2 | GTGACCTCAGAACCGATG | ||||||
| Lr34 | STS | csLV34-F | — | GTTGGTTAAGACTGGTGATGG | 57 | 150 | [41] |
| csLV34-R | TGCTTGCTATTGCTGAATAGT | ||||||
| Lr35 | SCAR | Sr39 F2 | — | AGAGAGAGTAGAAGAGCTGC | 57 | 900 | [55] |
| Sr39 R3 | AGAGAGAGAGCATCCACC | ||||||
| Lr38 | SCAR | SCAR982-F | — | GCTGAATCTGCGTATCGTCCC | 68 | 982 | [56] |
| SCAR982-R | GACTTGTTCTTCGGCGTGTTG | ||||||
(续附表)
| 基因/QTL Gene/QTL | 标记类型 Marker type | 引物 Primer | 距离 Distance (cM) | 引物序列 Primer sequence (5'-3') | 退火温度 Annealing temp. (℃) | 片段大小 Fragment size (bp) | 参考文献 Reference |
|---|---|---|---|---|---|---|---|
| 抗纹枯病Sheath blight resistance | |||||||
| Ses1 | SSR | Xgwm526-F | 27.9 | CAATAGTTCTGTGAGAGCTGCG | 58 | 140 | [8] |
| Xgwm526-R | CCAACCCAAATACACATTCTCA | ||||||
| 抗赤霉病 Scab resistance | |||||||
| Qfhs.ndsu -3BS | SSR | Xgwm533-F | 4 | AAGGCGAATCAAACGGAATA | 60 | 316 | [20] |
| Xgwm533-R | GTTGCTTTAGGGGAAAAGCC | ||||||
| SSR | Xgwm493-F | 4 | TTCCCATAACTAAAACCGCG | 60 | 175 | ||
| Xgwm493-R | GGAACATCATTTCTGGACTTTG | ||||||
| Qfhs.ifa -5A | SSR | Xgwm293-F | 2 | TACTGGTTCACATTGGTGCG | 55 | 205 | [20] |
| Xgwm293-R | TCGCCATCACTCGTTCAAG | ||||||
| SSR | Xgwm304-F | 2 | AGGAAACAGAAATATCGCGG | 55 | 202 | ||
| Xgwm304-R | AGGACTGTGGGGAATGAATG | ||||||
| Qfhi.nau -4B | SSR | Xgwm149-F | 1 | CATTGTTTTCTGCCTCTAGCC | 55 | 152 | [21] |
| Xgwm149-R | CTAGCATCGAACCTGAACAAG | ||||||
| Qfhi.nau -5A | SSR | Xwmc96-F | 1 | TAGCAGCCATGCTTAGCATCAA | 61 | 280 | [21] |
| Xwmc96-R | GTTTCAGTCTTTCACGAACACG | ||||||
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|