{kind=link}
{kind=link}
{kind=link}
利用SNP遗传图谱定位盐、旱胁迫下甘蓝型油菜种子发芽率的QTL
[荐红举* , 肖阳* , 李加纳, 马珍珍, 魏丽娟, 刘列钊*  ]
]
]
|
|


研究盐胁迫、干旱胁迫下甘蓝型油菜的发芽率, 寻找与发芽率相关联的分子标记, 可为油菜逆境胁迫下种子萌发的分子标记辅助育种提供理论依据。本研究以甘蓝型黄籽油菜GH06和甘蓝型黑籽油菜P174为亲本, 通过单粒传法(single seed descent, SSD)连续自交9代构建重组自交系群体。采用16 g L-1 NaCl溶液进行盐胁迫, 20%的PEG-6000溶液模拟干旱胁迫, 处理重组自交系种子并统计其发芽率。实验室构建的SNP遗传图谱, 包含2795个SNP多态性标记位点, 总长1832.9 cM, 相邻标记间平均距离为0.66 cM。利用该图谱并采用复合区间作图法(CIM)分析2种胁迫条件下第3天、第4天及累计4 d后发芽率的QTL。共检测到19个QTL, 分布于A01、A03、A06、A07、A09和C06染色体上。其中, 11个盐胁迫相关的QTL可解释的变异为4.9%~10.9%, 8个干旱胁迫相关的QTL可解释的变异为3.8%~6.9%; 并且在A03和A09染色体上, 盐胁迫和干旱胁迫下检测到的QTL有相近区段。研究结果表明油菜种子发芽率属于典型的数量性状, 受环境影响较大; 且随着胁迫时间的延长, 油菜种子启动了不同的基因来响应环境胁迫。
The objective of this study was to identify QTLs for seed germination percentage under salinity and drought stresses in
甘蓝型油菜( Brassica napus L .)是世界四大油料作物(大豆、向日葵、油菜、花生)之一, 在世界油料作物中占有重要地位[ 1]。盐害和干旱是影响油菜产量及品质的最重要的非生物胁迫因素。根据联合国教科文组织和粮农组织不完全统计, 全世界盐碱地面积约9.5亿公顷, 其中我国约有3300万公顷[ 2]。我国长江流域油菜播种期干旱最为严重, 9月至10月油菜播种后, 常常会遇到长达10~15 d的秋旱[ 3]。因此, 如何筛选出耐盐、抗旱的油菜新品种是育种家们亟待解决的问题。迄今为止, 各实验室已构建了至少20张甘蓝型油菜的遗传图谱[ 4], 并且定位了大量与芥酸[ 5, 6]、硫苷[ 7, 8]、含油量[ 6, 7, 8, 9]、菌核病抗性[ 10]等相关的基因, 包括一些重要基因。因此, 分子标记遗传图谱对于改良作物性状具有重要意义。
种子发芽率是作物育种中一个重要的衡量指标。国内外对番茄[ 11]、白菜[ 12]和拟南芥[ 13]等发芽率受环境因素的影响、发芽率与其他生理指标的关系的报道较多, 但对胁迫环境下种子发芽率的QTL定位研究相对较少。许耀照等[ 14]、原小燕等[ 15]和谢小玉等[ 16]用不同的方法探讨了盐害及干旱对甘蓝型油菜生长、发育的影响。Nguyen等[ 17]定位了甘蓝型黄籽油菜中所有与 AtPirin1同源的基因, AtPirin1在拟南芥中降低发芽率, 并抑制幼苗生长。通过比较控制发芽率、苗期生长、活力和与植物生长相关的激素的QTL的位置, 开发与 AtPirin1基因转录活性拷贝连锁的分子标记用于分子标记辅助育种。
植物耐盐性、抗旱性是受多基因控制的数量性状, 传统遗传学方法无法定位到单个抗性基因。在我国油菜抗逆性研究中, 重点还停留在生理生化研究的层面, 功能基因的深入研究还很少, 很多抗逆性问题还没有得到重视[ 18]。本文利用已构建的甘蓝型油菜重组自交系高密度SNP遗传图谱, 研究在盐和干旱两种胁迫条件下油菜种子发芽率, 讨论其间的相关性, 定位影响种子发芽的数量性状座位(QTL), 期望能够找到与甘蓝型油菜发芽率QTL紧密连锁的分子标记, 为油菜种子发芽的分子标记辅助选择提供依据。
黄籽母本GH06来源于甘蓝型黄籽油菜自交后代, 黑籽父本P174为油研2号经定向自交选出的后代。两亲本杂交后代通过单粒传法,连续自交至F10代,组成重组自交系群体RIL, 选取其中172个材料构建遗传连锁图谱, 收获相应种子(F11为本试验材料)。所有材料由重庆市油菜工程技术研究中心提供。
1.2.1 盐胁迫处理 以Hoagland营养液为母液, 配制质量浓度为16 g L-1的NaCl溶液。
选取大小一致、健康饱满的油菜种子, 置于铺
有2层定性滤纸的直径为9 cm的培养皿内, 每皿60粒, 每株系3次重复, 分别加入上述NaCl溶液, 第1天加入3 mL, 第2天起每天加入1.5 mL, 放入恒温培养箱, 设定培养箱25℃, 光照16 h, 黑暗8 h。
1.2.2 干旱胁迫处理 以去离子水为母液, 配制20% (w/w)的PEG-6000溶液, 作为模拟干旱胁迫试验处理条件。
挑选健康饱满的种子, 置于铺有2层定性滤纸的直径为9 cm的培养皿内, 每皿60粒, 每株系3次重复,分别加入上述PEG-6000溶液, 第1天3 mL, 自第2天起每天1.5 mL, 放入恒温培养箱(25℃, 光照16 h, 黑暗8 h)。
以芽长为种子一半作为发芽标准统计发芽种子数。从胁迫处理第2天开始,每天计数已发芽的种子,计算发芽率; 至胁迫处理后第6天结束。
选用西南大学重庆市油菜工程技术研究中心刘列钊等[ 19]于2013年利用SNP分子标记方法构建RIL (含172个株系)的遗传连锁图谱。该图谱共有2795个SNP多态性标记位点, 图谱总长1832.9 cM, 相邻标记间平均距离为0.66 cM。
参与图谱构建的172个株系中, 由于部分株系在生育期出现倒伏、病害等情况导致种子不够健康、饱满, 故仅对该群体中健康饱满无病害的163个株系的种子进行发芽试验。并对发芽率进行统计, 采用QTL分析软件Windows QTL Cartographer 2.5[ 20]及复合区间作图(composite interval mapping, CIM)法进行甘蓝型油菜种子发芽率的QTL定位及效应检测[ 21]。进行CIM分析时, 选取1 cM的步长(walking speed), 按照假定检测10和Zmapqtl模型3, 选取参数为1000次回归, 显著水平为0.01。LOD≥2.5时, 即认为该区间可能存在一个QTL。运行软件的结果可同时给出性状QTL的加性效应和解释的表型变异。按照McCouch等[ 22]的方法对检测到的QTL命名, 斜体小写字母“ q”加上性状的名字, 即用S (salinity的首字母)、D (Drought的首字母)分别代表盐胁迫及干旱胁迫, 紧接着的是染色体所在位置, 最后是QTL的序号。如 qS4-A03-1代表RIL群体与盐胁迫累积4 d发芽率相关的位于A03染色体上的第1个QTL。用软件Microsoft Excel及SPSS18.0统计分析表型数据。
亲本在去离子水中的发芽率分别为98.70%和96.20%。母本GH06在盐胁迫和干旱胁迫下发芽率分别为75.00%和80.00%, 父本P174的发芽率分别
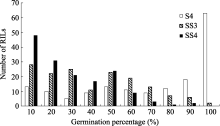
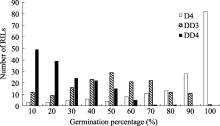
为50.00%和70.00%。考察了亲本及群体累计4 d盐胁迫及干旱胁迫下发芽率, 由于种子发芽受胁迫持续时间的影响, 故同时对第3和第4天的发芽情况统计分析, 结果显示种子发芽率呈现偏态连续分布(表1、图1和图2), 说明该性状受多基因控制, 群体符合QTL定位要求。
| 表1 亲本和重组自交系群体在不同胁迫环境下的发芽率 Table 1 Germination percentage under different stress environments in parents and RIL population |
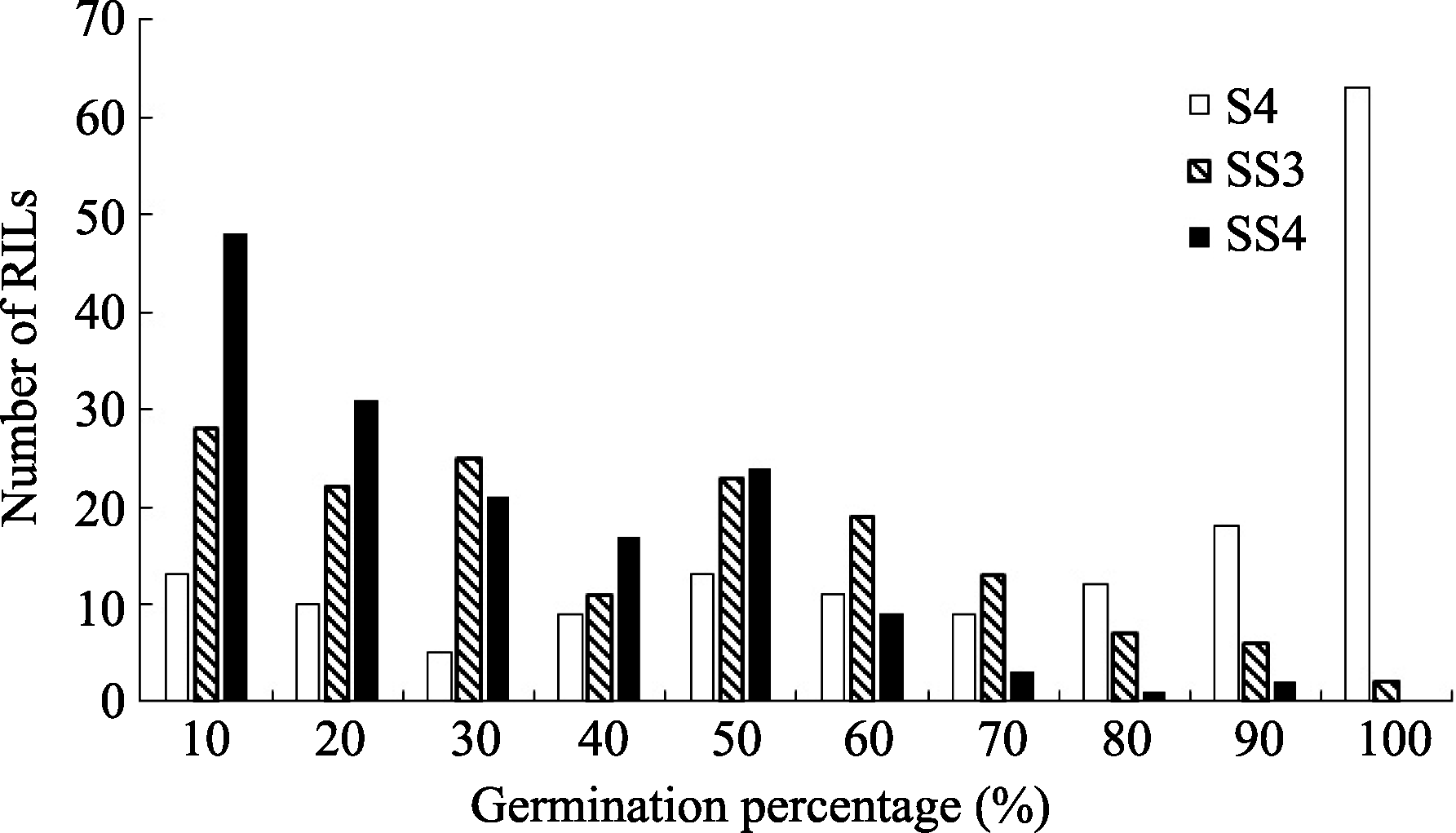 | 图1 盐胁迫下重组自交系群体发芽率的频率分布S4: 盐胁迫累计4 d; SS3和SS4: 盐胁迫第3和第4天。Fig. 1 Frequency distribution of the germination percentage of RILs under salinity stressS4: 4 days under salinity stress; SS3 and SS4: the third and fourth day under salinity stress. |
利用软件Windows QTL Cartographer 2.5及复合区间作图法对盐胁迫条件下累计4 d (S4)及第3天(SS3)、第4天(SS4)的种子发芽率进行QTL定位分析, 共检测到11个QTL, 其中与4 d发芽率相关的QTL有5个, 与盐胁迫第3天、第4天相关的有6个, LOD值在2.57~5.30之间, 单个QTL解释性状表现的变幅为4.90%~10.92% (表2)。这些QTL分别位于A01、A03、A07、A09和C06, 其中位于A07和
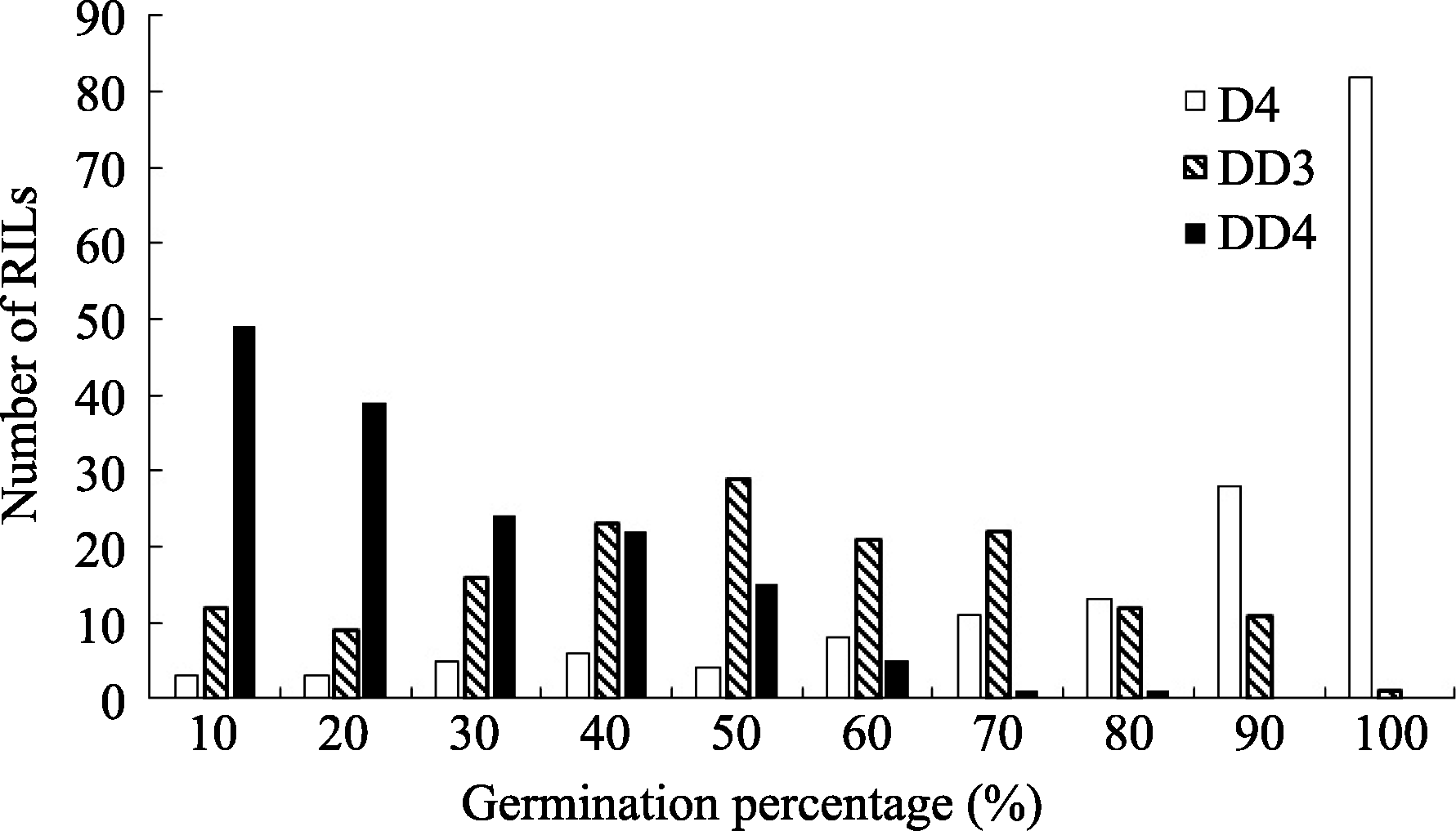 | 图2 干旱胁迫下重组自交系群体发芽率的频率分布D4: 干旱胁迫累计4 d; DD3和DD4: 干旱胁迫第3和第4天。Fig. 2 Frequency distribution of the germination percentage of the RILs under drought stressD4: 4 days under drought stress; DD3 and DDR: the third and fourth day under drought stress. |
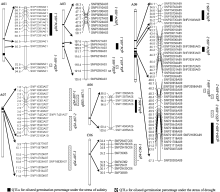
A09染色体较多(图3), 分别有4个和3个QTL, S4、SS3、SS4分别解释42.23%、24.29%和9.94%的表型变异。位于同一区域, 对于同一性状, 等位基因的增效作用既有来自父本的又有来自母本的。从加性效应值来看, 这11个QTL中仅有 qS4-A03-1和 qSS4- A01-1为正值, 其增效作用来自母本GH06的等位基因; 其余均为来自父本P174的等位基因, 起增效作用, 这说明在盐胁迫下种子发芽率的增效基因在亲本中有一定的基因离散。
| 表2 从重组自交系群体鉴定的油菜种子发芽率QTL Table 2 QTLs for germination percentage detected from RIL population of oilseed |
利用软件Windows QTL Cartographer 2.5及复合区间作图法对干旱胁迫条件下累计4 d (D4)及第3天(DD3)、第4天(DD4)的种子发芽率进行QTL定位分析, 共检测到8个QTL, 其中与累计胁迫4 d发芽率相关的QTL有2个, 与干旱胁迫第3天、第4天相关的有6个, LOD值在1.86~4.24之间, 单个QTL解释性状表现的变幅为3.15%~6.90% (表2), 为微效基因, 这些QTL分别位于A01、A03、A06和A09, 且位于染色体A03和A09较多(图3), 分别有2个和3个QTL, 其中 qDD3-19-1无法定位到具体染色体上, 仅知道它位于标记SNP28433附近。D4、DD3、DD4分别解释8.51%、3.15%及28.41%的表型变异。从加性效应值来看, 这8个QTL中 qD4-A06-1、 qDD4- A01-1和 qDD4-A03-1位点的加性效应来源于父本P174; 其余5个QTL位点的加性效应来自母本GH06。
在两种环境胁迫下QTL定位分析, 检测到6个(2种胁迫下各3个) QTL位于A09染色体, 4个(各2个)位于A03染色体, 2个(各1个)位于A01。其中位于A09染色体上密集检测到6个(盐胁迫和干旱胁迫各3个) QTL位于34.9~53.2 cM, 可解释的表型变异为5.1%~6.9%, 均为微效基因。
随着重要农作物基因组测序的进行或完成, 作物分子遗传育种进入高通量、大规模的全基因组水平分子设计平台。SNP分子标记因为分布广泛、多样性高、数量大等特点, 越来越受到分子育种家和系统进化研究者的重视[ 23, 24, 25]。利用SNP标记可以进行分子标记辅助育种、基因和QTL遗传定位、研究群体进化以及关联性分析等工作。
植物在生长发育过程中, 一般要同时应对多种生物或非生物胁迫, 尤其是干旱和盐碱通常重叠交叉发生[ 26]。植物的耐盐及抗旱性不仅与植物的种类、基因型以及生理生化反应等有关, 而且受干旱发生的时期、强度及持续时间的影响[ 27]。植物的耐盐及抗旱性受复杂的数量性状调控。在适应环境过程中, 植物演化形成了应对不同逆境的相同或特异的响应
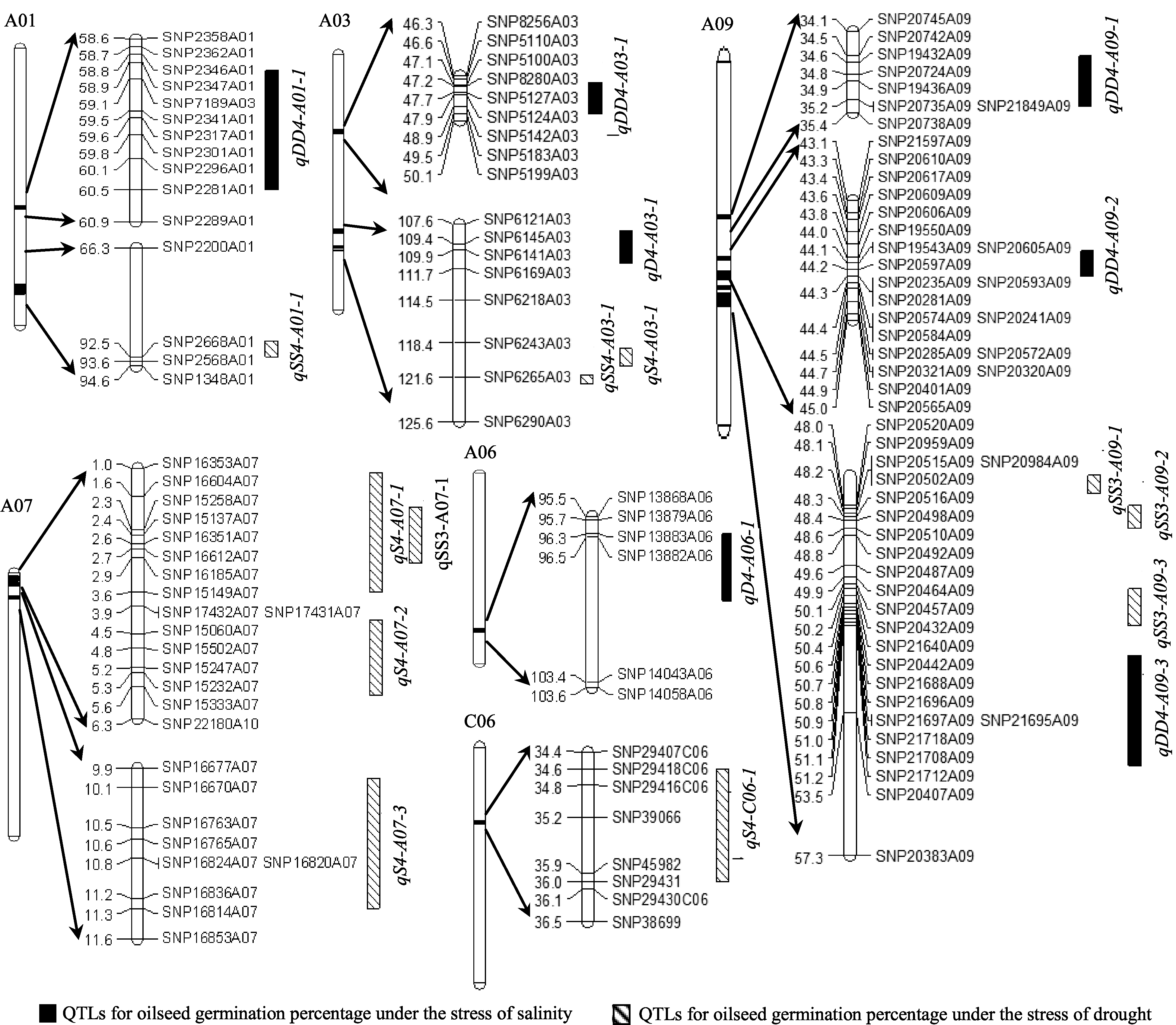 | 图3 甘蓝型油菜种子发芽率与盐胁迫、干旱胁迫相关的QTL在遗传图谱的分布Fig. 3 Putative QTLs of oilseed germination percentage under salinity and drought stresses in the genetic linkage map |
机制[ 28]。在现有的植物抗性研究中, 干旱胁迫与盐胁迫呈现正相关, 但在本文中并未得到类似的结果, 故还需要进一步的研究。由于尚没有在盐胁迫及干旱胁迫下甘蓝型油菜发芽率QTL的相关报道, 故本研究结果无法与公开发表的QTL进行比较。在本研究盐胁迫中, 位于A07染色体的2.7 cM处在S4、SS3同时检测出QTL, 说明该区域很可能存在此浓度盐胁迫下控制发芽的基因, 但最大的表型解释变异仅为10.9%, 需进一步试验验证; 并且加性效应均为负值, 说明该胁迫下对发芽率起正向调节作用的等位基因来自父本P174, 这在一定程度上说明父本P174皮厚耐盐胁迫而可能具有高发芽率; 在A09染色体42.4~50.8 cM之间检测到4个QTL, 可解释表型变异4.60%~6.12%, 其余QTL分布在A01、A03、C06染色体上, 可解释的表型贡献率在4.9%~7.9%之间, 均低于10%, 这在一定程度上说明在此浓度盐胁迫下油菜种子发芽率受微效多基因控制。本研究干旱胁迫中, 分别在A01、A03、A06、A09染色体上检测到QTL, 其中在A03染色体上检测到2个QTL, 但由于相距太远, 且能够解释的表型变异较低(分别为4.67%和5.70%, 表2), 故两者的功能需要进一步研究; 在A09染色体34.9~53.2 cM之间检测出3个QTL, 且相距较近, 但由于能够解释的表型变异过低, 故该区域的功能还需要进一步的分析。在干旱胁迫和盐胁迫中, 在A09染色体34.9~53.2 cM之间, 检测出6个QTL (各3个, 见表2), 说明该区域存在参与响应胁迫的相关基因, 而且QTL位点存在一定的交叉。然而, 干旱胁迫和盐胁迫第3天、第4天, 没有检测出相同或者相近的QTL区段, 可能是随着胁迫持续时间的延长, 油菜种子启动了不同的基因参与调控, 以响应不同阶段的胁迫环境。
由植物的抗性理论得知, 盐胁迫和干旱胁迫都能够引起植物类似的反应以应对不利的环境, 但统计分析两者却呈现负相关, 说明种子在盐胁迫、干旱胁迫下发芽是个复杂的生理过程, 该研究结果可为以后的甘蓝型油菜在胁迫环境下发芽的QTL定位提供参考, 今后的研究重点将是在此基础上对连锁群上较密的QTL精细定位, 以期找到胁迫条件下的主效基因, 进而为植物的抗盐、抗旱育种提供理论依据。
在A01、A03、A06、A07、A09、C06染色体上共定位了19个影响盐胁迫、干旱胁迫下油菜种子发芽率的QTL, 效应值都较小, 表现为微效多基因控制的数量遗传, 基因表达受环境影响较大; 第3天与第4天没有检测到相同或相近QTL区段, 可能是随着胁迫持续时间的延长, 植物启动了不同的胁迫基因以响应外界的胁迫环境。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|