



{kind=link}
{kind=link}
{kind=link}
{kind=link}
双季杂交晚粳稻超高产形成特征
[许轲1 , 张军1 , 花劲1 , 张洪程1, *  , 周培建
, 周培建2 , 程飞虎2 , 黄大山2 , 陈忠平2 , 陈国梁3 , 戴其根1 , 霍中洋1 , 魏海燕1 , 高辉1 ]
, 周培建|
|


本研究旨在明确传统双季晚籼稻地区双季杂交晚粳稻超高产产量构成及其群体特征, 阐明双季杂交晚粳稻超高产形成规律。以江西省上高县6.77 hm2连片双季杂交晚粳稻高产攻关示范方为依托, 选用杂交粳稻甬优8号为材料, 对中产(8.25~9.75 t hm-2)、高产(9.75~10.50 t hm-2)和超高产(>10.50 t hm-2) 3个产量水平群体的产量构成及群体特征进行系统比较研究。结果表明, 与中产、高产水平群体相比, 超高产水平群体表现穗数足、穗型大、群体颖花量多(50 000×104 hm-2以上)的显著特点, 但结实率和千粒重略低, 差异不显著; 群体茎蘖动态上, 群体起点较高, 可及时够苗; 够苗后增长平缓, 高峰苗数量较少、下降平缓, 成穗率高(78.0%左右)。群体叶面积指数前期增长较缓, 最大值出现在孕穗期, 为8.0左右, 此后下降缓慢, 成熟期仍保持3.5以上; 群体光合势生育前期较小, 中、后期较大, 抽穗至成熟期光合势为300×104 m2 d hm-2以上, 总光合势为560×104 m2 d hm-2以上。拔节前干物质量积累速度较慢, 拔节后积累速度较快, 至抽穗期群体干物质量为10.5 t hm-2左右, 抽穗后积累量亦高, 成熟期干物质量达19.0 t hm-2左右, 后期茎鞘物质转运率大于14.0%。超高产群体根量多、活力较强; 植株吸氮能力强、成熟期氮素累积量高, 氮素利用率40%以上。根据双季杂交晚粳稻超高产形成特征, 我们探讨了培育双季晚粳稻超高产群体的关键栽培技术。
, ZHOU Pei-JianThis study investigated the formation rule of super-high-yielding late
中国是世界上最大的水稻生产国和稻米消费国, 北方单季稻区传统种植的水稻品种以粳稻类型为主, 南方双季稻区以籼稻类型为主。随着经济的快速发展和人民生活水平的提高, 品质优、口感好的粳米深受大众喜爱, 人们对粳米的需求量日益增长; 特别是近年来, 南方“籼米改粳米”趋势明显, 已成为较大的粳米消费区[ 1, 2, 3]。我国粳稻种植面积占全国水稻总面积不足30%, 研究者提出了利用南方双季稻区充足的温光资源进行“籼稻改粳稻”, 以提高粳稻的总产量[ 4, 5]。如何充分挖掘作物品种的遗传潜力和配套建立高产栽培技术, 一直是农业研究的难点和重点[ 6, 7], 因此, 针对性研究双季晚粳稻超高产形成特征, 发展粳稻生产, 对我国南方“籼稻改粳稻”工程的实施显得十分迫切和重要, 对保障粮食安全、促进社会稳定具有重要意义。众多研究者们已经较系统研究了南方稻区籼稻高产特征与配套的栽培技术, 并提出了多项高产栽培技术模式[ 8, 9, 10, 11, 12]。但目前粳稻超高产的相关研究主要集中在单季稻区。在粳稻超高产研究上, 凌启鸿[ 13]提出了群体质量栽培理论与技术, 杨建昌等[ 14]在江苏淮北明确了常规中熟粳稻产量11.0 t hm-2以上的群体生育特性, 李刚华等[ 15]在苏中地区研明了粳型超级稻超高产群体特征及其对氮素的响应, 吴桂成等[ 16]阐明了粳型超级稻产量构成因素协同规律及超高产特征, 张洪程等[ 17]系统研究了长江中下游杂交粳稻13.5 t hm-2超高产群体动态特征及其形成机制, 并提出了水稻“精苗稳前、控蘖优中、大穗强后”超高产定量化栽培模式[ 18]。徐正进等[ 19]综合评述了北方粳稻超高产育种理论与实践, 讨论了与粳稻超高产有关的生理与遗传基础问题, 为粳稻超高产栽培提供了重要途径, 寒地水稻三超栽培技术在东北粳稻区得到了较大面积的推广应用[ 20]。同时, 各稻区在高产栽培模式的指导下, 创造了一系列的超高产纪录。综合表明, 目前粳稻超高产栽培研究仅局限于单季稻地区, 或仅针对双季稻地区的一季粳稻(中稻)[ 21, 22], 尚未见系统阐明双季晚粳稻超高产形成特征方面的研究报道。本研究以江西省上高县双季杂交晚粳6.77 hm2连片高产攻关方为对象, 根据土壤地力、群体起点、田间肥水情况等, 追踪调查代表性田块, 划定出中产(8.25~9.75 t hm-2)、高产(9.75~10.50 t hm-2)和超高产(>10.50 t hm-2) 3个产量等级群体, 研究它们的产量及其构成、群体茎蘖动态、叶面积和光合势动态、粒叶比、干物质生产积累与转运、根系活性和氮素吸收利用等若干特征, 提出双季杂交晚粳稻超高产群体质量指标, 以期为双季杂交晚粳稻超高产栽培与育种提供理论依据。
试验材料杂交粳稻甬优8号, 主茎总叶片数(N) 15~16叶, 伸长节间数(n) 5~6个, 全生育期150~ 155 d。2011—2012年, 种植于江西省上高县泗溪镇曾家村(115°09′E, 28°31′N), 设置双季晚粳稻高产精确定量栽培攻关试验方, 2年分别连片7.2 hm2和 7.8 hm2。试验田前茬为早籼稻, 晚稻季后茬为空闲田或种植绿肥作物紫云英。土壤类型为沙壤土, 地力中上等(前茬早稻产量为6.23~7.45 t hm-2), 土壤含有机质20.75~24.02 g kg-1、速效氮70.59~78.24 mg kg-1、速效磷26.65~32.13 mg kg-1、速效钾62.89~70.46 mg kg-1。6月20日播种, 湿润育秧, 7月20日移栽, 行株距为26.4 cm×13.2 cm, 每穴3~4粒种子苗。
氮肥施用总量为纯氮0.270 t hm-2, 基肥∶分蘖肥∶穗肥为4∶3∶3, 分蘖肥于移栽后7 d一次性施用, 穗肥于倒四叶、倒二叶抽出时分2次等量施入。氮∶磷∶钾比例为1.0∶0.5∶0.5。磷肥一次性基施, 钾肥分别于耕翻前、拔节期等量施入。移栽至有效分蘖期, 田面保持3 cm水层; 群体80%够苗后自然断水搁田, 搁田至土壤沉实后, 每次灌入3 cm水层, 待田间丰产沟(沟深15 cm)不见水时复灌, 周而复始直至成熟前1周。按粳稻超高产栽培措施统一进行其他管理措施。
同时在试验田中进行小面积的不同品种杂交晚粳稻连片超高产攻关田试验。根据前茬早稻产量和2010年预备试验结果分别在10块田田头作埂预留 20 m2左右小区不施用氮肥, 作为对照处理。
1.2.1 茎蘖动态测定 选择代表性田块, 定点20穴作为一个观察点, 每块田3个观察点。在移栽期、有效分蘖临界叶龄期、拔节期、抽穗期、乳熟期(抽穗后20 d)和成熟期观察茎蘖数消长动态。
1.2.2 干物质及叶面积测定 于有效分蘖临界叶龄期、拔节期、抽穗期、乳熟期(抽穗后20 d)、蜡熟期(抽穗后35 d)、成熟期(收获前1 d), 从各类型田块取10穴为1个样本, 将植株样本分解为绿叶、枯叶、茎、鞘和穗(抽穗以后), 测定叶面积和干物质重。在抽穗期采用长×宽×0.75法测定叶面积, 包括总茎蘖叶面积(总叶面积)、有效茎蘖叶面积(有效叶面积)和有效茎蘖顶三叶叶面积(高效叶面积)。每次测定重复3次。
1.2.3 根干重和伤流液测定 于拔节期、抽穗期、成熟期, 三类型产量等级每块田以5穴为1个样本, 重复3次, 剪去地上部分后以每穴根茬为中心, 挖取根土块(长30 cm、宽10 cm、高20 cm), 置40目尼龙网袋中, 用流水冲洗, 将根烘干后称重。在抽穗期、乳熟期和蜡熟期, 选择各产量等级平均茎蘖数的植株进行测定; 每一田块以5穴为1个样本, 重复3次, 于18:00在各茎距离地面12 cm处剪去地上部分植株(取样前排干田间水层), 将预先称重的脱脂棉放于茎剪口处, 套上伤流管, 第2天8:00取回带有伤流液的脱脂棉球并称重, 计算伤流液量。
1.2.4 产量测定 水稻收获前不同产量水平群体每一田块取5点, 每点取样50穴, 测定单位面积穗数, 并将稻穗装进塑料窗纱口袋内, 风干后, 脱粒、去除杂质(不包括空瘪粒), 求出50穴总粒数和每穗粒数; 采用水漂法去除空瘪粒, 求取结实率; 以1000粒实粒样本(风干种子)称重, 重复3次(误差不超过0.05 g)求取千粒重。成熟期各田块割取一定面积测定实际产量, 每块田5点, 每点20 m2。在连片高产示范方中选择代表性田块验收实际产量, 其中3块田由专家组进行实际产量验收。
1.2.5 计算与统计方法
光合势(×104 m2 d hm-2)=1/2(L1+L2)×(t2-t1), 式中, L1和L2为前后2次测定的叶面积(m2 hm-2), t1和t2为前后2次测定的时间(d);有效叶面积率(%)=有效LAI/最大LAI×100;高效叶面积率(%)=高效LAI/最大LAI×100;颖花/叶(cm2)=总颖花数/孕穗期叶面积;实粒/叶(cm2)=总实粒数/孕穗期叶面积;粒重(mg)/叶(cm2)=籽粒产量/孕穗期叶面积;群体生长率=(W2-W1)/(t2-t1), 式中, W1和W2为2次测定的干物质量, t1和t2为2次测定的时间(d);净同化率(g m-2 d-1)=[ln (LAI2)-ln (LAI1)]/ (LAI2-LAI1)×(W2-W1)/(t2-t1), LAI1和LAI2为前后2次测定的叶面积指数, t1和t2为前后2次测定的时间(d), W1和W2为前后2次测定的植株干物质量(g);表观输出率(%)=(抽穗期单茎茎鞘重-成熟期单茎茎鞘重)/抽穗期单茎茎鞘重×100;最大输出率(%)=(抽穗期单茎茎鞘重-齐穗后 15 d单茎茎鞘重)/抽穗期单茎茎鞘重×100;颖花(或实粒或粒重)根流量=茎秆伤流量(抽穗至蜡熟期平均值)/颖花数(或实粒数或粒重);氮肥当季利用率(%)=(施氮区植株总吸氮量-空白区植株总吸氮量)/总施氮量×100;氮肥生理利用率(%)=(施氮区产量-空白区产量)/(施氮区植株总吸氮量-空白区植株总吸氮量);氮肥农学效率=(施氮区产量-空白区产量)/总施氮量;氮肥表观生产力=籽粒产量/总施氮量;运用Microsoft Excel软件进行数据计算与作图; 运用DPS进行统计分析。
甬优8号2年3个产量水平(中产、高产、超高产)群体平均实际产量分别为9.34、10.59和10.98 t hm-2, 超高产较高产增加3.77%, 高产较中产增加13.37%, 差异显著或极显著(表1)。从产量构成因素分析, 3个产量水平群体颖花量(×104 hm-2)平均值分别为 41 330.93、46 921.55和50 101.43, 超高产较高产、高产较中产分别增加6.79%和13.52%, 差异均极显著; 平均结实率分别为86.54%、86.26%、85.95%, 超高产较高产、高产较中产分别减少0.36%和0.32%, 不同产量等级间差异不显著; 千粒重分别为26.69、26.66和26.24 g, 不同产量等级间差异不显著。从总颖花量构成的两个因素来看, 两年超高产较高产穗数平均增加1.24%、每穗粒数平均增加5.16%, 高产田的穗数较中产田增加8.95%, 每穗粒数增加4.95%。
综上分析可知, 双季晚粳稻从中产到高产水平需显著增加单位面积穗数和穗粒数, 保持较高的结实率和千粒重, 从高产向超高产水平发展, 适当增加穗数, 主攻大穗, 并保持较高的结实率和千粒重。综合两年研究结果表明, 双季晚粳稻超高产群体的适宜产量构成应以足量穗数与较大穗型协调产出足量的群体颖花量(50 000×104 hm-2以上), 同时保持较高的结实率(86.0%左右)和千粒重(26 g左右)。
| 表1 不同产量水平杂交晚粳稻产量及其构成 Table 1 Grain yield and its components of late japonica hybrid rice under different yield levels |
2.2.1 群体茎蘖动态 由图1可见, 与中产和高产田相比, 超高产田移栽期生长前期茎蘖数增长缓慢, 有效分蘖临界叶龄期(N-n)、拔节期、抽穗期、乳熟期群体茎蘖均较低, 直至成熟期略高于高产田, 极显著高于中产田。超高产田移栽期茎蘖数约为最终成穗数的46%, 中产和高产田分别约为最终穗数的44%和42%, 超高产群体生育前期增长速度较高产和中产群体慢, 至有效分蘖临界叶龄期(N-n)不同产量等级群体均达到预期穗数; 各群体均于拔节期达到高峰苗数, 超高产田茎蘖数约为成熟期穗数的1.2倍, 高产田为1.3倍, 中产田为1.4倍左右; 此后超高产群体平稳下降, 抽穗期穗数等于或略多于成熟期穗数; 分蘖成穗率超高产田为77.12%~78.14%, 高产田为74.14%~75.05%, 中产田仅为66.26%~68.11%。
2.2.2 叶面积指数与光合势动态 不同产量等级水稻群体叶面积指数动态与各生育期茎蘖动态趋势相似(图2)。与中产和高产群体相比, 超高产群体叶面积指数(LAI)在移栽期较高(差异不显著), 之后中产和高产田群体叶面积增长速度较快, 叶面积指数高, 直至拔节期均高于超高产田。不同产量等级水稻最大LAI均出现于孕穗期, 其中超高产田(8.02~8.04)群体极显著或显著高于中产(7.06~7.11)和高产田(7.49~7.83); 此后, 超高产田群体叶面积下降略缓于中产和高产田(图2), 至成熟期超高产田群体LAI仍保持3.5以上。
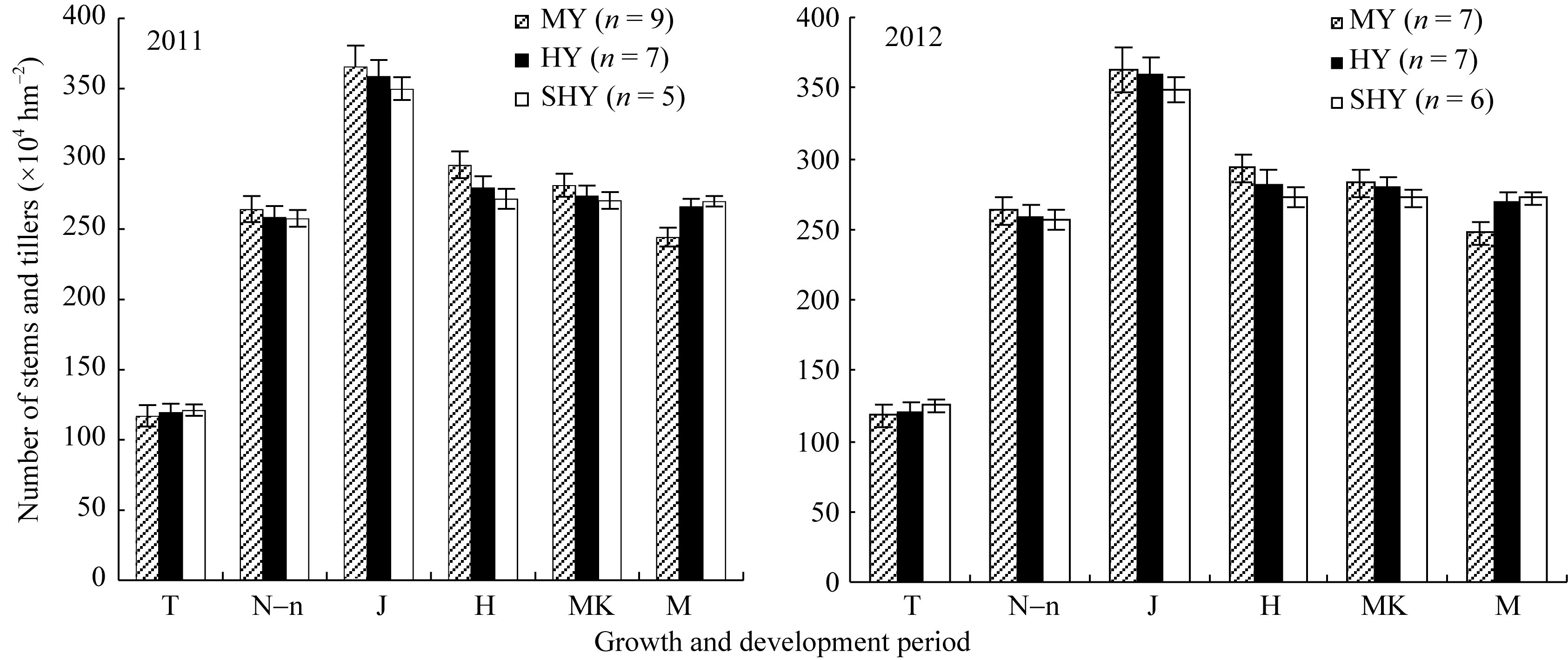 | 图1 不同产量水平杂交晚粳稻不同生育时期的茎蘖数Fig. 1 Number of stems and tillers of late japonica hybrid rice at each growth period (×104hm-2) |
MY: 中产; HY: 高产; SHY: 超高产; T: 移栽期; N-n: 有效分蘖临界叶龄期; J: 拔节期; H: 抽穗期; MK: 乳熟期; M: 成熟期。
MY: medium yield; HY: high yield; SHY: super-high yield; T: transplanting stage; N-n: critical leaf-age for productive tillers; J: jointing; H: heading; MK: milky stage; M: maturity.
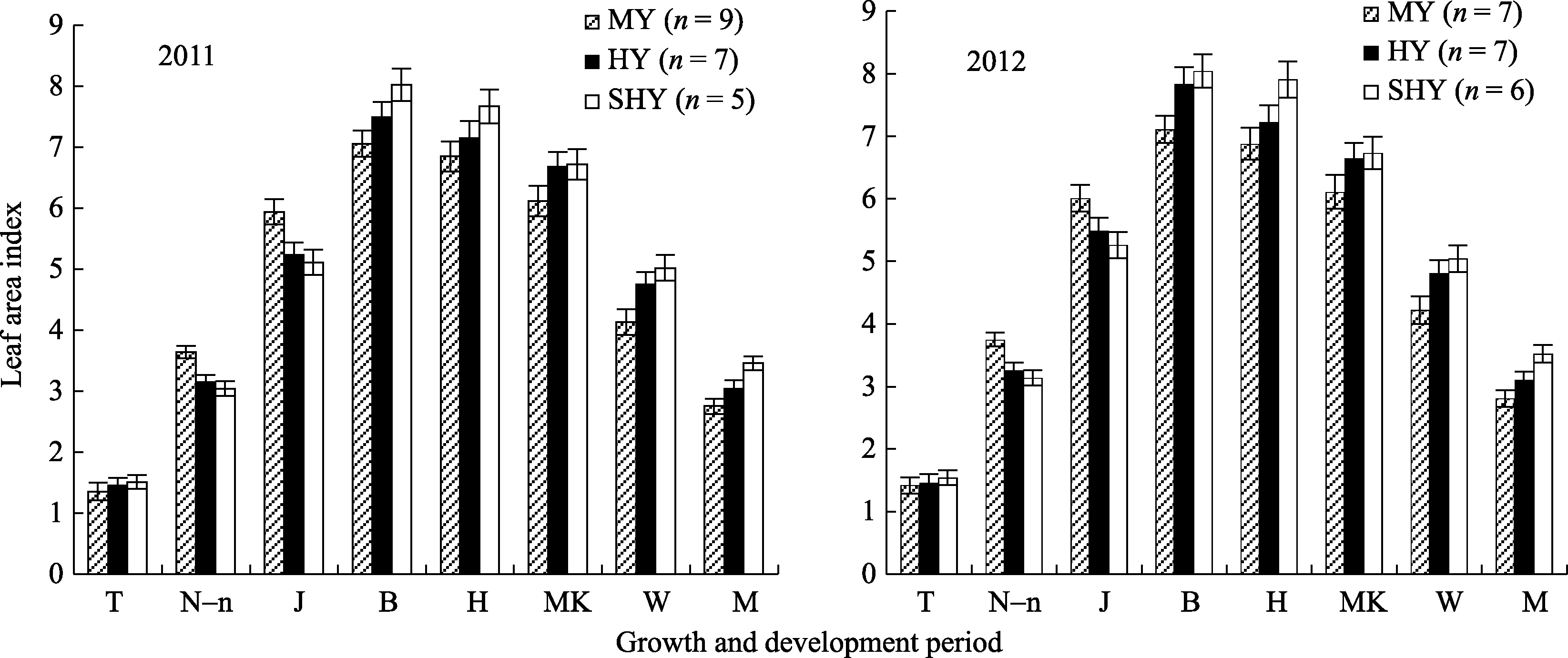 | 图2 不同产量水平杂交晚粳稻不同生育时期叶面积指数缩写同图1。B: 孕穗期; W: 蜡熟期。Fig. 2 Leaf area index of late japonica hybrid rice at each growth periodAbbreviations are the same as given in Figure 1. B: booting; W: waxy stage. |
两年不同产量水平杂交晚粳稻的光合势(单位土地面积绿叶面积与光合时间乘积)与叶面积变化趋势基本一致(图3)。在移栽至有效分蘖临界叶龄期和有效分蘖临界叶龄期至拔节期2个生育阶段, 超高产群体光合势略小于高产群体, 显著小于中产群体; 在拔节至抽穗期、抽穗至乳熟期、乳熟至蜡熟期和蜡熟至成熟期4个生育阶段, 超高产和高产群体的光合势均显著高于中产群体。超高产群体不同生育期的光合势(×104 m2 d hm-2): 移栽至有效分蘖临界叶龄期(N-n)为40左右, (N-n)至拔节期为60左右, 拔节期至抽穗期为160左右, 抽穗期至乳熟期为130左右, 乳熟期至蜡熟期为100左右, 蜡熟期至成熟期为70左右; 全生育期总光合势>560; 其中抽穗至成熟期的光合势2年平均为304.94, 占总光合势的54%左右, 显著或极显著高于高产(占总光合势的50%)和中产(占总光合势的48%)群体。
2.2.3 群体干物质生产与运转特征
(1) 群体干物质积累 对2年不同产量水平水稻群体不同生育期干物质积累动态分析可知(图4), 在有效分蘖临界叶龄期(N-n)和拔节期, 超高产田与高产田干物质积累量差异较小, 略低于中产田; 此后, 不同生育时期三类产量水平间干物质积累量差异显著, 且呈超高产>高产>中产的趋势。超高产群体干物质积累量(t hm-2), (N-n)叶龄期为1.6左右, 拔节期为3.5左右, 抽穗期为10.5左右, 乳熟期为14.5左右, 蜡熟期为15.5左右, 成熟期为19.0左右。收获指数, 超高产田为0.490, 略高于高产田(0.480), 显著高于中产田(0.460)。
(2) 抽穗期叶面积组成与粒叶比 表2表明, 超高产田群体抽穗期有效叶面积率、高效叶面积率及粒叶比均显著高于高产和中产水平, 2年超高产群体的有效叶面积率和高效叶面积率平均分别为94.40%、73.72%, 粒叶比[颖花/叶(cm2)、实粒/叶(cm2)、粒重(mg)/叶(cm2)]平均分别为0.634、0.545、
 | 图3 不同产量水平杂交晚粳稻不同生育阶段光合势Fig. 3 Photosynthetic potential of late japonica hybrid rice at each growth stage (×104m2 d hm-2) |
MY: 中产; HY: 高产; SHY: 超高产; T: 移栽期; N-n: 有效分蘖临界叶龄期; J: 拔节期; H: 抽穗期; MK: 乳熟期; W: 蜡熟期; M: 成熟期.
MY: medium yield; HY: high yield; SHY: super-high yield; T: transplanting stage; N-n: critical leaf-age for productive tillers; J: jointing; H: heading; MK: milky stage; W: waxy stage; M: maturity.
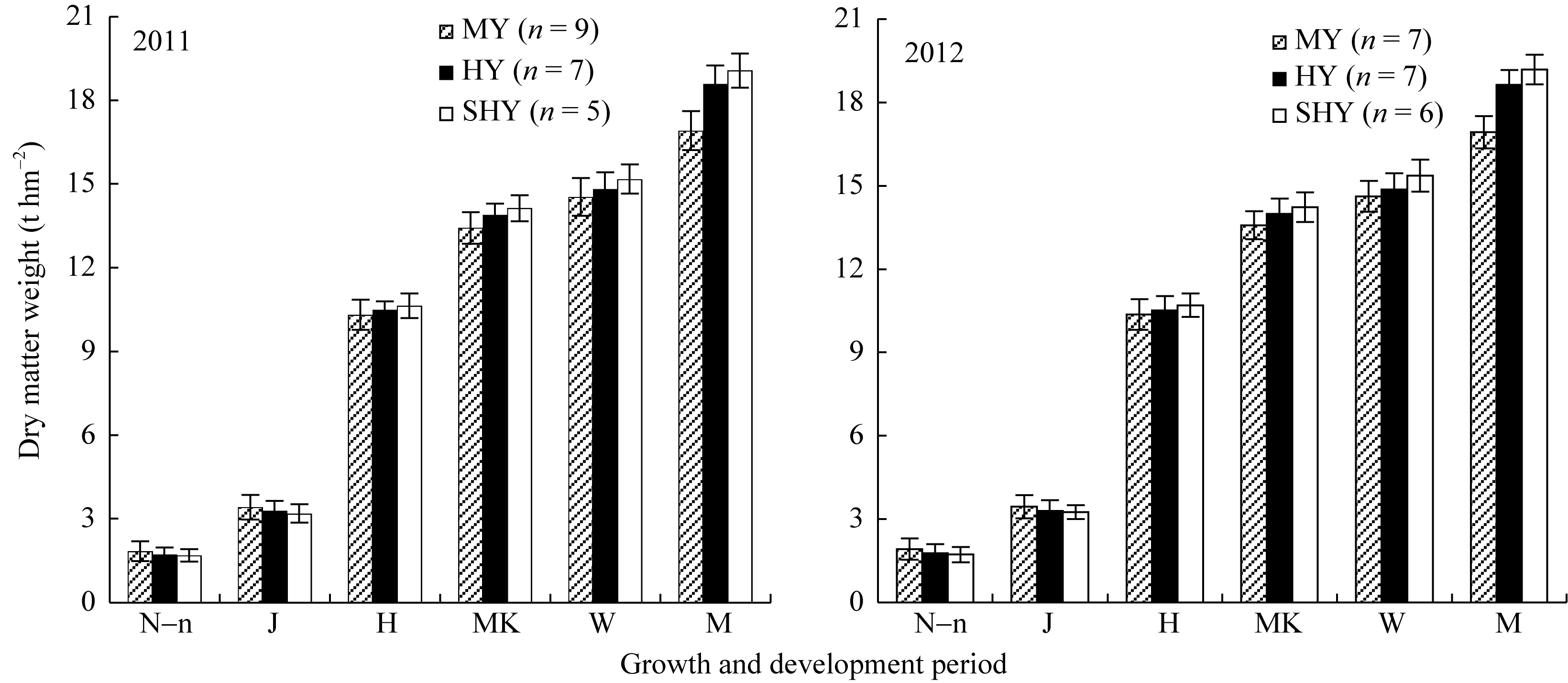 | 图4 不同产量水平杂交晚粳稻不同生育时期干物质积累量MY: 中产; HY: 高产; SHY: 超高产; N-n: 有效分蘖临界叶龄期; J: 拔节期; H: 抽穗期; MK: 乳熟期; W: 蜡熟期; M: 成熟期。Fig. 4 Dry matter accumulation of late japonica hybrid rice at each growth periodMY: medium yield; HY: high yield; SHY: super-high yield; N-n: critical leaf-age for productive tillers; J: jointing; H: heading; MK: milky stage; W: waxy stage; M: maturity. |
| 表2 不同产量水平杂交晚粳稻群体抽穗期干物质积累量、叶面积组成与粒叶比 Table 2 Dry matter accumulation, components of leaf area, and grain-leaf ratio in heading under different yield levels |
MY: 中产; HY: 高产; SHY: 超高产。同一年内同列不同字母分别表示在0.05和0.01水平上差异显著性。
MY: medium yield; HY: high yield; SHY: super-high yield. Values followed by different letters of the same year are significantly different at the 0.05 (lowercases) and 0.01 (capitals) probability levels.
14.24, 有效叶面积率和高效叶面积率及粒叶比均显著高于高产群体(平均分别为91.70%、71.65%; 0.601、0.521、13.87), 极显著高于中产群体(平均分别为84.99%、62.87%; 0.552、0.478、1277)。可见, 超高产群体具有更大、更厚的冠层结构(超高产群体抽穗期干物质量较高产群体高1.65%, 高产群体较中产群体高1.42%); 同时, 超高产水平优良的群体结构, 即较高的有效叶面积率、高效叶面积率、群体粒叶比, 可促进超高产群体安全生长。
(3) 抽穗后期光合物质生产与转运特征 分析2年各群体抽穗期至成熟期光合物质生产表明(表3), 超高产群体净积累量平均为8.47 t hm-2, 占产量平均为77.59%, 显著高于高产群体(积累量为8.12 t hm-2, 占产量平均为76.66%), 极显著高于中产群体(积累量为6.58 t hm-2, 占产量平均为70.49%), 说明超高产群体籽粒产量主要来源于抽穗后较高的群体光合物质生产与积累量。超高产群体抽穗后群体生长率(g m-2 d-1)、光合势(×104 m2 d hm-2)和净同化率(g m-2 d-1) 2年分别平均为11.43、304.94、2.614, 均极显著高于高产和中产群体, 说明超高产群体后期(抽穗期至成熟期)具有高效的光合能力。超高产群体单茎茎鞘最大输出率与最大输出物质转运率2年平均分别为33.84%和14.50%, 极显著高于高产和中产水平群体(表3), 成熟期茎鞘表观输出率(2年平均为15.66%)显著小于后两者。可见, 超高产水平群体抽穗前积累的茎鞘物质的运转能力和抽穗后群体光合能力强; 同时, 更高效的光合能力使超高产群体具有较强的二次物质生产与积累能力, 最终表观输出率最低, 茎鞘充实更好, 有利于增强超高产群体抗倒伏能力。
2.2.4 群体根系特征 根干重是根系生长状况的集中体现, 反映了根系发达程度。2年不同产量等级晚粳稻单茎根干重动态, 随生育进程呈先增加后降低趋势, 根冠比呈降低趋势(表4)。超高产水平群体的根重和根冠比均显著或极显著高于高产和中产水平群体。2年超高产田晚粳稻单茎根干重(g)、根冠比平均值, 拔节期分别为0.262和0.290, 抽穗期分别为0.374和0.133, 成熟期分别为0.195和0.095。各群体晚粳稻颖花根流量[mg 颖花-1 h-1、mg 实粒-1 h-1和mg粒重(g)-1 h-1]均是超高产水平显著高于高产水平, 高产水平极显著高于中产水平, 2年超高产水平群体颖花根流量[mg颖花-1 h-1、mg实粒-1 h-1和mg粒重(g)-1 h-1]平均值分别为2.318、2.470和13.888, 表明灌浆充实期超高产水平群体根系活力旺盛, 利于群体安全成熟。
2.3.1 主要生育时期植株含氮率和吸氮量 由表5可见, 不同产量等级晚粳稻植株含氮率随生育期推进逐渐降低, 拔节期最高, 成熟期最低; 不同产量等级高低关系在不同生育期表现不一致, 拔节期超高产水平显著低于高产, 高产水平极显著低于中产, 抽穗期、成熟期超高产水平显著高于高产水平, 高产水平极显著高于中产水平; 超高产水平植株含氮率2年平均分别为, 拔节期1.73%、抽穗期1.61%、成熟期1.33%。不同产量等级植株吸氮量随生育进程呈增加趋势, 成熟期达到最大值。不同产量等级间, 拔节期超高产水平植株吸氮量低于高产, 差异显著; 高产水平低于中产水平, 差异不显著; 抽穗期和成熟期超高产水平显著或极显著高于高产水平, 高产水平显著或极显著高于中产水平。超高产群体主要生育时期植株吸氮量2年平均分别为, 拔节期0.056 t hm-2、抽穗期0.171 t hm-2、成熟期0.220 t hm-2。
2.3.2 氮肥利用效率 由表6可见, 不同产量水平双季晚粳稻百千克籽粒需氮量均是超高产水平最高, 10.5 t hm-2以上产量等级的百千克籽粒需氮量为2.0 kg左右; 超高产水平的氮肥当季利用率、生理利用率、氮肥农学利用率及氮肥表观生产力均最高, 2年平均分别为41.45%、44.01%、18.24和40.67, 显著或极显著高于高产水平(平均分别为39.02%、42.71%、17.06和39.19)和中产水平(平均分别为38.01%、37.67%、14.32和34.57)。
| 表3 不同产量水平杂交晚粳稻群体抽穗后光合物质生产特征 Table 3 Production of dry matter of late japonica hybrid rice after heading under different yield levels |
| 表4 不同产量水平杂交晚粳稻根系干重及根冠比特征 Table 4 Dry weight of root and root-shoot ratio of late japonica hybrid rice at each growth period under different yield levels |
| 表5 不同产量水平杂交晚粳稻主要生育时期植株含氮率和氮积累量特征 Table 5 N content and uptaking in plant of late japonica hybrid rice at main growth period under different yield levels |
| 表6 不同产量水平杂交晚粳稻氮肥利用效率 Table 6 N use efficiency of late japonicahybrid rice under different yield levels |
2011—2012年, 在江西上高县进行不同双季杂交晚粳稻品种超高产连片攻关试验(表7), 每一品种种植面积0.067 hm2以上, 结果表明超高产产量构成与甬优8号有较大相似性; 双季杂交晚粳稻产量10.5 t hm-2以上的群体颖花量均为50 000×104 hm-2以上, 结实率86.0%左右, 千粒重26 g左右。
对不同品种杂交晚粳稻群体特征及养分吸收利用特性分析发现(表8), 不同品种杂交晚粳稻具有与甬优8号类似特性。超高产群体茎蘖成穗率78%以上, 其中2011年甬优7号成穗率高达80% (表8); 超高产群体成熟期总生物量均在19.0 t hm-2以上, 收获指数0.490以上, 颖花/叶(cm2)为0.700左右, 氮素当季利用率40%以上。另外, 不同品种杂交晚粳稻的根重及活力(数据未列)等其他特征与甬优8号有一定相似性, 在此不赘述。
| 表7 不同品种杂交晚粳稻超高产攻关方产量及构成 Table 7 Grain yield and its components of different cultivars of late japonicarice in demonstration fields |
| 表8 不同品种杂交晚粳稻超高产攻关方群体特征及氮素利用率 Table 8 Population characteristics and N utilization of different cultivars late japonica rice in demonstration fields |
水稻产量由库容(由有效穗数和每穗粒数组成)和充实度(结实率和千粒重)两部分构成。水稻超高产实现关键在于如何稳定实现产量构成因素在较高水平上的协同发展[ 23]。前人较一致认为“以足量大穗形成高群体颖花量”是水稻超高产产量形成的一条重要途径[ 14, 16, 24, 25]。本试验结果表明, 在南方双季稻区不同产量水平杂交晚粳稻高产实现的途径有所不同, 产量水平由中产到高产, 主要依靠提高有效穗数并适当增大穗型; 产量由高产到超高产水平, 需在适量增加穗数的同时主攻大穗来提高群体颖花量。表面上看与单季粳稻和双季晚籼稻超高产产量形成规律一致, 实质上有较大区别。本研究中甬优8号结实率和千粒重较在单季稻区种植均有所降低, 原因是引种至双季稻区作为晚稻种植后全生育期大大缩短, 籽粒的灌浆充实不及单季稻区。若进一步挖掘其高产潜力, 应从扩大库容着手, 同时与稳定的结实率、千粒重协同发展。南方双季晚稻较同期单季稻区气候有“高温、高湿、温差小”的特点, 单位面积所容纳的穗数有限(粳稻群体茎蘖数多, 易感染纹枯病和稻曲病, 同时群体过大易贪青晚熟), 仅依靠增穗扩库容不符合客观实际, 因此, 实现双季杂交晚粳稻较大库容关键在于稳定或适当增加单位面积穗数, 同时主攻大穗, 提高群体“安全库容”[ 16]。双季杂交晚粳稻超高产群体保持稳定的结实率和千粒重难度较大, 种植双季晚粳稻的各个环节必须更加精确, 实现超高产群体颖花量和较高的结实率、千粒重协同发展。本文仅针对大穗型杂交粳稻品种甬优8号, 其它类型粳稻品种产量形成特征还有待进一步研究。
双季晚粳稻超高产群体起点较高, 茎蘖增长速度慢, 在有效分蘖临界叶龄期及时够苗, 且高峰苗数量小, 后期(抽穗至成熟期)群体茎蘖数消亡较平缓, 抽穗期达预期穗数, 成穗率较高(78%以上)。虽然双季晚粳稻和单季粳稻超高产群体均是同一生育时期够苗, 但双季晚粳稻群体茎蘖数更需要依靠主茎数(群体起点高), 单季稻主要依靠分蘖数(群体起点低), 因双季晚粳稻全生育期较单季粳稻明显缩短, 其中缩短较多的是营养生长期[ 26], 分蘖的发生主要在该生育时期, 因此双季晚粳稻超高产田基本苗高于单季粳稻。另外, 双季晚粳稻超高产田茎蘖增长速度小于单季粳稻, 分析其原因可能是双季晚粳稻秧苗移栽大田后, 田间温度较高, 茎蘖发生受阻; 生育中期和后期的茎蘖动态与单季稻区超高产田变化趋势相似[ 16]。
与双季晚粳稻中产和高产群体相比, 超高产群体叶面积指数、光合势和干物质重方面表现为, 拔节期前生长量小, 中期生长量明显提高, 抽穗至成熟期物质生产能力较强(叶面积、光合势和干物质量均高)。源、库、流特征表现为粒叶比高、物质运转率高、收获指数高; 根系特征表现为根量大(抽穗期至成熟期单茎根重及根冠比均高)、根系活性强(根系伤流量大); 氮素吸收利用特征表现为吸收能力强, 积累量大(抽穗期至成熟期, 群体植株含氮率和含氮量均显著或极显著高于中产和高产水平群体), 氮素当季利用率高(40%以上)。与单季粳稻和双季籼稻超高产群体质量特征[ 14, 15, 17, 27, 28]相比有一定相似性, 但双季晚粳稻超高产具有自身特征, 例如, 群体物质生产方面, 双季晚粳稻生育前期物质积累量较少, 产量更多来自于生育中后期的物质生产, 这正是双季晚粳稻高产优势所在, 即双季稻区晚稻季充足的温光资源使晚粳稻生育后期具有更强的物质生产能力[ 26]。因此, 应依据双季晚粳稻群体特征适时调整栽培策略。
本课题组已连续多年在江西不同纬度地区开展“籼改粳”研究, 晚粳稻较当地晚籼稻表现出较大产量优势, 2011—2012年上高县6.77 hm2连片示范方平均实际产量均超过了9.75 t hm-2(江西省农业厅组织专家进行田间实际产量测定), 较当地晚籼稻高15%以上(江西省农业厅组织对当地晚籼稻田间实际产量测定, 最高产为8.25 t hm-2左右), 达到超高产水平[ 26]; 同时, 最高产田块实际产量甚至超过了10.5 t hm-2(2011、2012年甬优8号最高产量分别为10.8 t hm-2和10.9 t hm-2)。可见, 杂交晚粳稻在当地种植的产量潜力较大, 9.75 t hm-2是推广晚粳稻的产量起点。为此, 依托本研究, 单独分析10.5 t hm-2以上超高产田块, 提出双季杂交晚粳稻10.5 t hm-2以上超高产群体指标(表9), 供双季晚粳稻超高产栽培和育种参考。
| 表9 双季杂交晚粳稻10.5 t hm-2以上超高产群体指标 Table 9 Population indexes of super high-yielding (>10.5 t hm-2) late japonicahybrid rice in double-cropping rice area |
进行超高产栽培时, 要达到上述群体指标, 应以超高产群体形成特征为依据, 采取综合栽培措施, 限于文章篇幅, 在此列出几项关键栽培技术供探讨。
(1)选择生育期适宜、穗型较大的粳稻稻品种。双季晚粳稻营养生长期较单季稻区大大缩短, 粳稻品种有效分蘖叶位减少, 应选择生育期适宜且穗型大、灌浆速率快的粳稻品种。
(2)延长育秧时间, 培育壮秧。通过延长秧苗生长期, 向前延伸双季晚粳稻营养生长期, 培育叶蘖同伸壮秧(移栽时分蘖2个以上), 充分挖掘晚粳稻超高产潜力。
(3)建立较大群体起点。双季晚粳稻营养生长期短, 靠分蘖争取足穗较为困难。一方面可培育带蘖壮秧, 另一方面可适当增加基本苗, 依据基本苗公式精确群体起点。
(4)精确肥水管理。根据晚粳稻产量目标采用斯坦福方程精确计算施氮量。同时, 双季晚粳稻氮肥运筹与单季粳稻有一定差别, 依靠前期重施基蘖肥, 促使晚粳稻群体茎蘖早发快发, 及时够苗, 最终达到足穗; 近年专题试验研究认为氮肥运筹应以7︰3较为适宜(相关研究将另文阐述)。双季晚粳稻水分管理除参照单季粳稻超高产方案外, 还应区别于当地晚籼稻品种类型, 粳稻后期需水量大, 需根据后期群体状况进行水分精确管理。
与中产和高产水平群体相比, 双季杂交晚粳稻超高产群体产量构成特征为穗数足、穗型大、结实率和千粒重正常; 群体茎蘖动态特征为群体起点高、及时够苗和够苗后及时转入平缓增长, 高峰苗数量小、后期群体下降平缓、至抽穗期具有适宜穗数; 群体质量特征为光合势、粒叶比、物质运转率、收获指数高, 根系发达、活力强, 氮素吸收多、利用率高。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|