
{kind=link}
GenBank数据库中黄麻EST-SSR标记的开发及其通用性评价
[张立武*  , 袁民航
, 袁民航* , 何雄威, 刘星, 方平平, 林荔辉, 陶爱芬, 徐建堂, 祁建民* ]
, 袁民航, 何雄威, 刘星, 方平平, 林荔辉, 陶爱芬, 徐建堂, 祁建民]
|
|


第一作者联系方式: 张立武, E-mail:lwzhang@fafu.edu.cn,zhang_liwu@hotmail.com, Tel: 0591-86393065; 袁民航, E-mail:947866907 @qq.com
** 同等贡献
从GenBank公共数据库中下载黄麻表达序列标签838条, 利用SSRPrimer软件对其进行SSR位点查找, 利用Primer 3.0软件设计66对SSR引物, 通过琼脂糖凝胶研究这些SSR引物的PCR扩增特点, 以检测其多态性。结果表明, 66对SSR引物在黄麻属6个不同类型材料的扩增中, 42 (63.6%)对引物至少在2个材料之间存在多态性。(AT)n重复基元和(GC-)n丰富的三核苷酸重复基元多态性较高, 可作为黄麻SSR标记引物设计的首选。黄麻EST-SSR标记开发效率较高, 不仅可以丰富黄麻分子标记的数量, 而且为剖析黄麻重要性状的遗传机制奠定基础, 这对于黄麻的遗传基础研究具有重要应用价值。
, YUAN Min-Hang, HE Xiong-Wei, LIU Xing, FANG Ping-Ping, LIN Li-Hui, TAO Ai-Fen, XU Jian-Tang, QI Jian-Min
** Contributed equally to this work
In this study, a total of 838 jute expressed sequence tags (EST) were derived from GenBank. Using software Primer 3.0, 66 pairs of primers were designed based on the simple sequence repeat (SSR) sites screened by the online program SSRPrimer. These primers were amplified in six jute accessions of different types to test their polymorphism. The agarose gel electrophoresis results showed that 42 pairs of primers (63.6%) were polymorphic between at least two accessions and the PCR products were clear and stable fragment. These polymorphic EST-SSRs contained rich motif (AT)n or (AC-)n, which could be considered as the dominant motif in jute genome. Therefore, it is effective to develop SSR markers in jute from ESTs deposited in GenBank. The EST-SSRs developed will not only facilitate genetic research in jute (
黄麻是椴树科(Tiliaceae)黄麻属( Corchorus)一年生草本植物, 是重要韧皮纤维作物。在世界上黄麻产量与种植面积仅次于棉花, 主要在亚洲(印度、孟加拉国、中国和泰国)和拉丁美洲种植。生产上栽培的有长果种( Corchorus olitorius L.)和圆果种( Corchorus capsularisL.), 染色体数均为2 n=14。黄麻纤维细软、强力大、吸湿性好、散水快, 可纺性好, 用途十分广阔[ 1]。开发重复性和稳定性较好的SSR分子标记, 对于黄麻遗传多样性研究、重要性状的遗传基础剖析等方面具有重要意义。
然而, 相对于水稻等模式作物, 黄麻分子标记开发及遗传连锁图谱的构建与其他作物差距很大, 目前国内外报道较少[ 2, 3, 4, 5, 6]。这些研究报道多用RAPD、AFLP和ISSR, 少量用SSR等为分子标记。如Nishat等[ 5]用抗寒性有差别的2个亲本O-9897和Acc.1805, 构建了F2代作图群体, 用8个ISSR引物构建了总长度为87.3 cM的包括3个连锁群的长果种黄麻遗传连锁图谱, 3个连锁群的大小在4.8~52.9 cM之间, 相邻标记平均距离为8.73 cM, 该图谱远远不能满足基因定位、基因克隆等研究的要求。Das等[ 6]用纤维品质有差别的2个亲本JRO524和PPO4, 构建了F6代RIL作图群体, 用36个SSR标记构建长果种黄麻遗传连锁图谱, 但只有6个连锁群, 标记密度达21.8 cM。由于SSR标记具有高效、高共显性等优点, 尤其是重复性好, 不同研究间可互比较, 是物种的起源与进化、遗传连锁图谱构建等研究优先考虑的分子标记[ 7]。国内外学者采用Primer3等软件已开发了水稻[ 8]、大麦[ 9]、玉米[ 10]、油菜[ 11, 12, 13]等作物的SSR标记, 为研究该作物的遗传多样性、遗传连锁图谱构建、关联分析等打下了基础。
本研究旨在通过对GenBank公共数据库黄麻属EST中SSR信息进行分析, 发掘SSR位点并探明其规律, 探讨SSR在不同类型黄麻材料的多态性, 为黄麻遗传研究提供SSR信息。
用于研究的黄麻属2个种6个不同类型材料均来自福建农林大学麻类遗传育种研究室(表1)。
| 表1 供试黄麻种质资源的品种名称及来源 Table 1 Names and origins of tested germplasm in jute |
从GenBank公共数据库(http://misuse.ncbi.nih.gov/)中下载黄麻表达序列标签EST (expressed sequence tag) 838条(截至2013年11月8日)。利用网上的SSRPrimer工具(http://hornbill.cspp.latrobe.edu.au/ssrdiscovery.html)搜索这些序列所包含的SSR, 筛选SSR的标准为: 二核苷酸重复次数>6次, 三核苷酸重复次数>4次, 四核苷酸重复次数>3次, 五核苷酸重复次数>2次。利用Primer3软件对其进行SSR位点查找, 设计113对引物。通过与发表文献比较, 剔除已发表的SSR, 共合成66对EST-SSR引物, 其中编号CcSSR001~CcSSR045为圆果种黄麻EST-SSR, CoSSR046~CoSSR066为长果种黄麻EST-SSR。这66对SSR引物由生工生物工程(上海)有限公司合成。
采用改良的CTAB法提取黄麻基因组DNA[ 14]。
PCR体系10 μL, 含50 ng μL-1DNA 2.0 μL、10 μmol L-1引物0.5 μL、0.5 U μL-1 Taq酶0.1 μL、10 mmol L-1dNTPs 0.2 μL、10×PCR buffer 1 μL、50 mmol L-1Mg2+0.8 μL、dd H2O 5.4 μL。PCR程序为94℃预变性3 min; 94℃变性30 s, 60℃退火30 s, 72℃延伸45 s, 共10个循环, 每个循环退火温度降低0.5℃; 94℃变性30 s, 55℃退火30 s, 72℃延伸 45 s, 共35个循环; 最后72℃延伸10 min, 10℃保存10 min。 Taq酶、dNTP等试剂购自生工生物工程(上海)有限公司, 采用1.5%琼脂糖凝胶电泳。
从GenBank数据库EST中总共开发113个SSR (表2), 其中三核苷酸重复所占比例最多, 为69.9%,其次是四核苷酸重复, 占13.3%, 二核苷酸重复和五核苷酸重复比较少, 分别占9.7%和7.1%, 在二至五核苷酸重复类型中三核苷酸和五核苷酸重复序列平均长度最长, 均为18.1 bp, 最短的是四核苷酸重复, 为13.1 bp。
| 表2 黄麻EST中SSR分布频率 Table 2 Frequencies of SSR number in jute EST |
在这113个EST-SSR中共找到43种不同类型的重复基元, 其中二核苷酸重复3种、三核苷酸重复23种、四核苷酸重复11种、五核苷酸重复有6种(表3)。其中二核苷酸重复基元以(CT)n和(AT)n为主, 三核苷酸重复基元中(ACC)n、(CGA)n和(AGA)n所占比例高。
| 表3 黄麻EST-SSR中不同重复基元的分布 Table 3 Frequencies of EST-SSR repeat types in jute |
通过与已发表文献[ 3- 4, 6]比较, 剔除已发表的引物, 共合成66对EST-SSR引物。利用这66对SSR引物对黄麻属2个种6个不同类型材料进行扩增(表4和图1), 64 (97.0%)对引物得到强且清晰的PCR产物, 42 (63.6%)对引物至少在2个材料间存在差异, 表现出多态性。其中, 有些引物在这6个材料中表现出种间和种内多态性, 如CcSSR008、CcSSR013、CcSSR016、CcSSR024、CcSSR029、CcSSR040、CoSSR049、CoSSR050、CoSSR052和CoSSR053等; 有些引物仅表现出种间多态性, 如CcSSR007、CcSSR015、CcSSR026等。分析这些有多态性引物的重复基元构成(表4), 我们可以看出, (AT)n重复基元和(AC-)n丰富的三核苷酸重复基元多态性较高, 可作为黄麻SSR标记引物设计的首选。
| 表4 黄麻EST-SSR的多态性和扩增效率 Table 4 Amplified and polymorphic primers of EST-SSR in jute |
| 续表4 黄麻EST-SSR的多态性和扩增效率 Continued 4 Amplified and polymorphic primers of EST-SSR in jute |
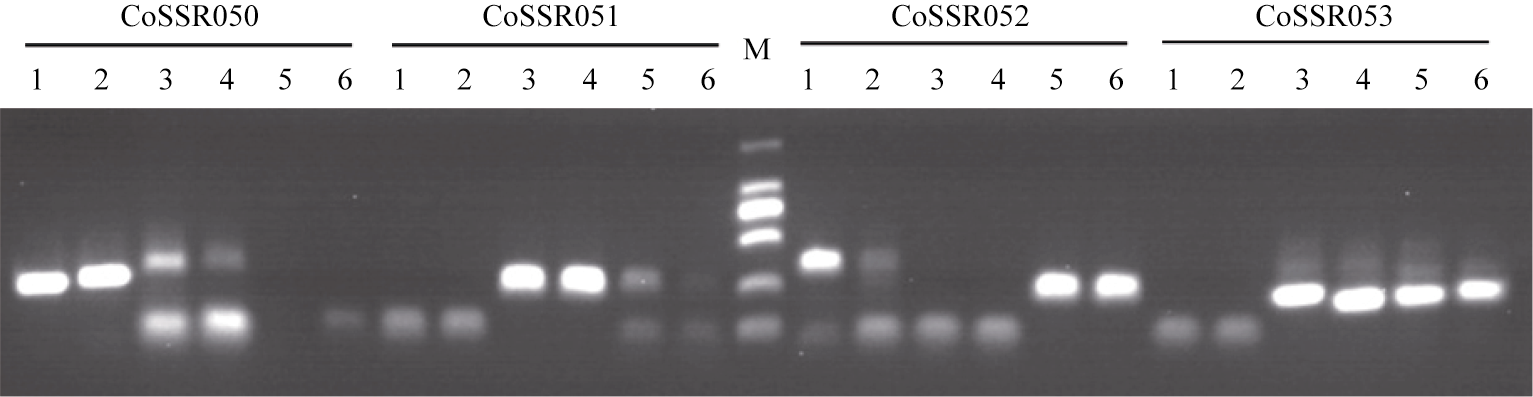 | 图1 SSR标记(CoSSR50-53)对黄麻属6个不同类型材料的扩增结果Fig. 1 Amplification results of SSR primers (CoSSR50-53) in six jute accessions of different types |
本研究从GenBank数据库中下载黄麻EST序列838条, 共包含有113个SSR。结果表明, 大部分重复基元为二至五核苷酸, 其中三核苷酸重复类型数量较多, 所占比例为69.9%。而这与在大多数植物基因组中三核苷酸重复的分布频率较高的结果是一致的[ 11, 12, 13]。Hong等[ 15]分析白菜中SSR重复基元发现, 三核苷酸重复出现的频率是37%。Goff等[ 16]在水稻中, 也发现基因组SSR中三核苷酸重复占59%。本研究中, 黄麻EST-SSR的分析结果显示, 二、三、四核苷酸中所有的重复基元类型都有, 但不同重复基元的比例存在显著差异。其中二核苷酸重复中(AT)n和(CT)n类型最丰富, 三核苷酸重复中(AC-)n最多, 这种分布趋势与Das等[ 6]对来源于长果种黄麻BAC末端序列SSR的分析结果一致。四核苷酸重复中具有较高比例的(AAGA)n和(AAAT)n基元, 这与Mir等[ 3]对黄麻SSR的分析结果类似。
Temnykh等[ 8]研究发现, SSR长度大于或等于 20 bp的SSR具有较高的多态性。为保证开发的SSR标记具有较高的多态性, 我们也采用这样方法开发了113个EST-SSR标记。其中二核苷酸的(AT)n基元和三核苷酸的(AC-)n基元在基因组中出现的频率较高, 而且在黄麻中扩增的多态性也较高, 是开发黄麻SSR引物的优势重复基元。因此, 通过黄麻EST序列开发SSR标记是可行的, 表现较高多态性和具有比对信息的SSR标记, 不仅可以丰富分子标记的数量, 有利于高密度遗传图谱构建, 还将为在分子水平上研究黄麻重要性状的遗传基础及黄麻的起源与进化奠定基础。
目前黄麻属约有40多个种, 具有栽培价值的2个种是圆果种黄麻和长果种黄麻[ 1]。利用黄麻属2个种间6个不同类型材料进行多态性检测, 可以检测出2个种间以及种内的多态性。张亚东等[ 17]分析不同杨树种间的遗传差异发现, EST-SSR比基因组的SSR在不同杨树种间具有更好的通用性。这是因为EST-SSR标记来源于相对保守的转录区域, EST- SSR在近缘物种甚至远缘物种间具有更好的通用性。
基于此, 本研究开发的EST-SSR也可以在圆果种、长果种和假黄麻( Corchorus acutangulus Lam)等黄麻属不同种间得到应用。
此外, 由于基因组包含内含子, 与外显子相比, 内含子在植物进化过程中受人类驯化的选择相对较少, 保留了大量的序列多态性, 所以, 通常情况下基因组多态性高于EST多态性[ 8]。但本研究表明, GenBank数据库中EST-SSR有65.6%呈现出清晰的多态性条带, 说明EST-SSR也具有较高多态性。但不同研究者, 由于使用的材料不同, 可能结果也不一致。Basu等[ 4]用SSR及AFLP标记分析时, 却认为SSR标记在黄麻种质资源中的多态性较低。这可能跟该研究只使用长果栽培种和圆果栽培种作为试验材料有关。在本研究中, EST-SSR检测出的多态性包含黄麻属不同种间的多态性, 所以表现出较高的多态性。
利用GenBank公共EST数据库成功开发出66对SSR引物, 这些SSR标记具有较好的稳定性和多态性。(AT)n重复基元和(AC-)n丰富的三核苷酸重复基元多态性高, 可作为黄麻SSR标记引物设计的首选。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|