{kind=link}
{kind=link}
[刘臣* , 陆建农* , 殷学贵*  , 毕川, 文淡悠, 郑军, 刘帅, 石卓兴, 成粤湘]
, 毕川, 文淡悠, 郑军, 刘帅, 石卓兴, 成粤湘]
, 毕川, 文淡悠, 郑军, 刘帅, 石卓兴, 成粤湘]
|
|


用YC2 (高杆)×YF1 (矮杆)和YC1 (高杆)×YF1 (矮杆)组合衍生的2个F2代群体, 对蓖麻株高性状进行相关、回归和QTL定位分析。结果表明, 株高与主穗位高、主茎节长和主茎茎粗之间显著正相关, 但与主茎节数不相关; 主穗位高与主茎节数、主茎节长和主茎茎粗之间呈显著正相关; 主茎节数与主茎节长之间呈显著负相关。利用QTLNetwork 2.0软件在YC2×YF1的F2群体中检测到株高、主穗位高、主茎节数、主茎节长和主茎茎粗的5、4、6、3和2个QTL, 分别解释了45.9%、45.3%、66.1%、55.4%和12.6%的总变异。在YC1×YF1的F2群体中检测到3、4、5、1和2个上述5性状的QTL, 分别解释了26.0%、25.5%、35.4%、37.4%和7.6%的总变异。证明QTL间的“一因多效”和连锁是株高、主穗位高和主茎节长之间高度相关的遗传基础, 加性效应是株高、主穗位高和主茎节长的主要遗传组分, 互作效应是主茎节数和主茎茎粗的主要遗传组分。建议育种上将主穗位高和主茎节长作为株高早期选择和预测的间接指标, 并将多节数和短节间作为高产育种的主攻方向。
, BI Chuan, WEN Dan-You, ZHENG Jun, LIU Shuai, SHI Zhuo-Xing, CHENG Yue-Xiang
The traits related to plant height were analyzed with correlation, regression and QTL mapping in two F2 populations derived from crosses of YC2×YF1 and YC1×YF1, respectively. The results indicated that plant height (PH) was significantly and positively correlated with bearing height of primary raceme (PRH), length of main stem internode (MSIL) and main stem diameter (MSD) but not with node number of main stem (MSNN), PRH was significantly and positively correlated with MSNN, MSIL and MSD, MSNN and MSIL was significantly and negatively correlated with each other. In F2 population from a cross of YC2×YF1, five, four, six, three, and two QTLs for PH, PRH, MSNN, MSIL, and MSD were detected by QTLnetwork 2.0, which explained for 45.9%, 45.3%, 66.1%, 55.4%, and 12.6% of the phenotypic variation, respectively. In F2 population from a cross of YC1×YF1, three, four, five, one, and two QTLs for the five traits mentioned above were detected, which explained for 45.9%, 45.3%, 66.1%, 55.4%, and 12.6% of the phenotypic variation, respectively. We concluded that the pleiotropy or linkage between QTLs results in the positive correlation between PH, PRH and MSIL, the additive effect was the main genetic component of PH, PRH, MSIL, and the epistatic effect is the main genetic component of MSNN and MSD. It suggested that PRH and MSIL should be used to select and predict PH indirectly in the early stage, more MSNN and shorter MSIL should be regarded as the target traits for high-yield breeding.
矮化育种实现了水稻和小麦产量的突破性增长[ 1, 2]。目前, 小麦中已有25个主效 Rht (reduced height)基因被命名[ 3, 4], 13个被定位[ 5], Rht-B1b[ 3] 、Rht-D1b[ 5] 、Rht-8和 Rht-9[ 6]等数量性状基因已被克隆。水稻中至少有70个矮杆和半矮杆基因被发现和鉴定, 主要分为与 SD-1相关的基因和与 SD-1互不等位的基因[ 7], 其中50多个已被定位[ 8], 部分基因已被克隆, 它们大多参与GA等植物激素的合成和细胞信号转导过程[ 9]。Gramene数据库中已有1011个水稻株高相关的QTL。关于蓖麻株高的遗传研究报道较少, Lavanya等[ 10]和李金琴等[ 11]认为蓖麻株高受1对单显基因控制。蓖麻株高相关基因细胞色素 P450基因片段已被克隆, 该基因被干扰后可明显降低转基因植株的株高[ 12]。
我国是蓖麻油产品主要出口国之一, 其中癸二酸出口量占世界第一, 但由于种植面积下降, 同时也成为蓖麻籽第一进口大国。我国蓖麻单产高于印度, 但较高的生产成本使我国蓖麻籽失去了价格竞争优势。品种产量低导致了种植效益低下, 影响了种植者的积极性。矮杆品种抗倒伏性强、经济系数高, 便于合理密植和机械化操作, 能够大幅度提高单产和种植效益。然而, 目前生产上所用的品种大多是高杆品种, 加之纯度低、抗逆性差, 产量低而不稳。
蓖麻具无限生长习性, 其株高(plant height, PH)与生育期紧密相关, 是后期表达的复杂数量性状。一年生情况下, 一般在二级分枝穗停止伸长时测量。主穗位高(bearing height of primary raceme, PRH)、主茎节数(node number of main stem, MSNN)、主茎节长(length of main stem internode, MSIL)是株高的重要组成部分, 受环境影响相对较小, 为早期表达的性状, 在主穗停止伸长时即可测量。本文利用2个高×矮组合的F2群体对蓖麻的株高、主穗位高、主茎节数、主茎节长和主茎茎粗(简称茎粗, 下同)(main stem diameter, MSD)进行相关、回归分析和QTL定位, 以期为揭示蓖麻株高遗传和矮化育种提供参考。
以矮杆亲本YF1为父本, 高杆亲本YC1和YC2为母本, 创建了2个F2群体, 分别为161株和129株。3个亲本均为本课题组经单株连续多代自交培育的稳定自交系。2011年11月2日于广东海洋大学东海岛蓖麻试验基地种植亲本、F1和F2群体, 行株距1 m×1 m。试验地肥力均匀、地势平坦。亲本及后代表现见表1。
| 表1 亲本及后代的表型分析 Table 1 Phenotype variation of parents, F1, and F2 populations |
主茎节长是由主穗位高除以主茎节数获得, 茎粗为主茎上、中、下3节茎粗的平均值, 按照《蓖麻种质资源描述规范和数据标准》[ 13]调查其余3个性状。将两群体数据整合后, 采用SPSS18.0进行性状间的相关和回归分析。
本试验所用引物有三类, 一是自行开发的SSR引物, 编号为RCM+开发序号, 共计1100对(未发表)。二是SRAP引物, 参照已发表的序列。三是SSR与SRAP上下联之间的组合扩增引物, 编号为SSR的开发序号+F(或R)+Em(或Me)序号, 例如97F/Em2为RCM97的上联与SRAP引物中的Em2组合扩增。
株高与主穗位高、主茎节长和茎粗之间均呈极显著正相关, 与主茎节数不相关; 主穗位高与主茎节数、主茎节长和茎粗均呈极显著正相关; 主茎节数和主茎节长呈极显著负相关(表2)。
| 表2 性状间的相关系数 Table 2 Correlation coefficients between traits |
回归分析显示株高与主穗位高、主茎节长之间存在极显著( F=106.3> F0.01=4.75)的线形关系, y=68.59+0.70 x1+ 7.09 x2( t1=5.68> t0.01=2.60, t2=7.24> t0.01=2.60)。
以上分析表明, 利用主穗位高和主茎节长对蓖麻的株高进行早期间接选择和预测是可行的, 尤以主茎节长作用更加明显。主成分分析也表明(数据未显示)主茎节长对株高的贡献更大。
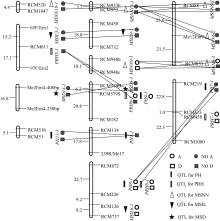
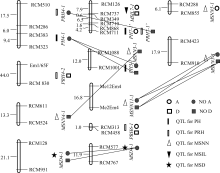
YC2×YF1组合F2群体(群体1, 后同)的定位结果见表3和表5, YC1×YF1组合F2群体(群体2, 后同)的定位结果见表4和表6。
2.2.1 株高 群体1中共检测到5个株高QTL, 解释了45.9%的总变异, 其中加、显和上位性3种效应的贡献率分别占总贡献率的64.79%、5.66%和29.48%。 PH8-1的单位点贡献率为32.23%, 其中加性效应贡献率为23.70%; 在群体2中检测到3个株高QTL, 解释了26%的总变异, 3种效应的贡献率分别占总贡献率的69.77%、0和30.03%。 PH7-1’仅有加性效应, 贡献率达18.14%。
2.2.2 主穗位高 群体1中共检测到4个主穗位高QTL, 解释了45.3%的总变异, 3种效应的贡献率分别占总贡献率的54.08%、22.96%和22.96%。 PRH7-1和 PRH8-1
| 表3 (YC2×YF1)F2群体的QTL定位结果 Table 3 QTLs detected in (YC2×YF1)F2 population |
| 表4 (YC1×YF1)F2群体的QTL定位结果 Table 4 QTLs detected in (YC1×YF1)F2 population |
| 表5 (YC2×YF1)F2群体的QTL遗传效应 Table 5 QTL effects in (YC2×YF1)F2 population |
的单位点贡献率分别为15.4%和14.6%, 其中加性效应贡献率分别为8.0%和11.6%, 显性效应贡献率分别为7.4%和3.0%, 无上位性效应。群体2中检测到4个主穗位高QTL, 解释了25.5%的总变异, 3种效应的贡献率分别占总贡献率的24.82%、43.88%和30.04%。 PRH6-1贡献率为13.77%, 虽位于第6连锁群, 但与群体1第7连锁群的 PRH7-1同与标记 RCM313连锁, 效应组成和大小相近, 应为同一QTL。
2.2.3 主茎节数 在群体1中共检测到6个主茎节数QTL, 解释了66.1%的总变异, 3种效应的贡献率分别占总贡献率的28.14%、1.20%和70.66%。其中只有2个QTL存在单位点效应, 其余4个只有上位性效应。在群体2中共检测到5个主茎节数QTL, 解释了35.4%的总变异, 3种效应的贡献率分别占总贡献率的24.77%、0和75.23%。其中只有1个QTL存在单位点效应, 其余5个只有上位性效应。
| 表6 (YC1×YF1)F2群体的QTL遗传效应 Table 6 QTL effects in (YC1×YF1)F2 population |
2.2.4 主茎节长 群体1共检测到3个主茎节长QTL, 解释了55.36%的总变异, 3种效应的贡献率分别占总贡献率的65.53%、18.77和15.87%。 MSIL8-1贡献率最大, 其加性效应贡献率为36.28%, 显性效应贡献率为10.4%, 无上位性效应。群体2只检测到1个主茎节长QTL MSIL7-1, 解释了44.99%的总变异, 加性和显性效应的贡献率分别占总贡献率的80.68%和19.31%。
2.2.5 茎粗 2个F2群体中各检测到2个茎粗QTL, 都只有上位性效应, 累计贡献率分别为12.60%和7.64%, 说明茎粗的遗传率很低, 狭义遗传率更低。
虽然2个群体中每个性状检测到的QTL数目以及位置有较大差异, 但实际上, QTL尤其是主效QTL的数目、效应和作用方式有较高的一致性。两群体中各有1个株高主效QTL、1个主穗位高主效QTL (群体1共有2个主效QTL)和1个主茎节长主效QTL, 它们的位置、效应、作用方式都很接近(表7)。由于遗传图谱不够饱和(全长 2818.2 cM, 231个标记, 平均间距12.2 cM, 连锁群数目大于染色体基数), 加之群体间的遗传背景差异, 同一性状主效QTL在两群体中所在的连锁群不同, 但从QTL与标记的连锁关系判断, 它们仍在染色体同一区域(表7), 很可能为同一QTL。株高、主穗位高和主茎节长的遗传以加性效应为主, 主茎节数和茎粗的遗传以上位性效应为主, 这在2个群体中是一致的(表5和表6)。
| 表7 两群体中主效QTL的比较 Table 7 Comparison of major QTLs between two populations |
群体1中(表3、表5和图1), PH8-1与 PRH8-1等位, 单位点解释26.3%、14.6%的株高和主穗位高变异, 其中加性效应分别占90.11%和79.45%, 主要作用方式相同, 应属同一QTL。 PH9-1与 PRH9-1等位, 单位点解释6.04%和4.90%的株高和主穗位高变异, 加性效应分别占100.00%和32.03%, 均无显性效应, 其余效应均为负值, 应属同一QTL。群体2中(表4、表6和图2), PH7-1’与 PRH7-1’紧密连锁, 遗传距离仅2 cM。可见, “一因多效”和QTL间的连锁是株高与主穗位高之间显著相关的重要原因。在群体1中, 3个主茎节长QTL中有2个与株高、主穗位高QTL连锁(表3和图1)。其中 MSIL8-1与 PH8-1、 PRH8-1连锁, 遗传距离为19.9 cM, MSIL5-1与 PRH5-1连锁, 遗传距离为14.2 cM。在群体2中, MSIL7-1与 PH7-1’ 、PRH7-1’连锁, 遗传距离分别为12.6 cM和10.6 cM。连锁位点的贡献率及主要作用方式接近。可见, QTL间的连锁也是主茎节长与株高和主穗位高之间显著相关的重要原因。
在2个群体中检出的主茎节数QTL数目较多, 都以上位性为主。大多数与株高QTL之间不存在一因多效和连锁, 但这不能作为株高与主茎节数之间不相关的原因。因为同样不存在一因多效和连锁, 主穗位高与主茎节数之间又显著相关。主茎节数对株高应该是有贡献的, 二者之所以不相关, 很可能是相对于株高后期的大生长量和对环境的敏感性, 主茎节数的直接贡献已经很微弱了。主茎节数与主茎节长之间的显著负相关, 表明二者之间存在某种相互抑制机制, 值得进一步研究。
株高在F2群体中接近正态分布, 为典型数量性状, 这与以前报道[ 10, 11]不同, 与水稻[ 18]、小麦[ 5, 19]、玉米[ 20]和黄瓜[ 21]等作物研究结果一致。张维峰等[ 22]报道, 蓖麻株高主要受基因的加性效应控制, Narkhede等[ 23]报道, 蓖麻株高的遗传中累加基因占优势, 主茎节数的遗传中上位基因占优势, 与本试验结果有一致性。
株高、主穗位高、主茎节长在F2代均为正态分布, 均有主效QTL被检出, 而主茎节数呈明显偏态分布, 却无主效QTL被检出, 说明不能仅从变异分布类型判断是否存在主基因(或主要基因)。
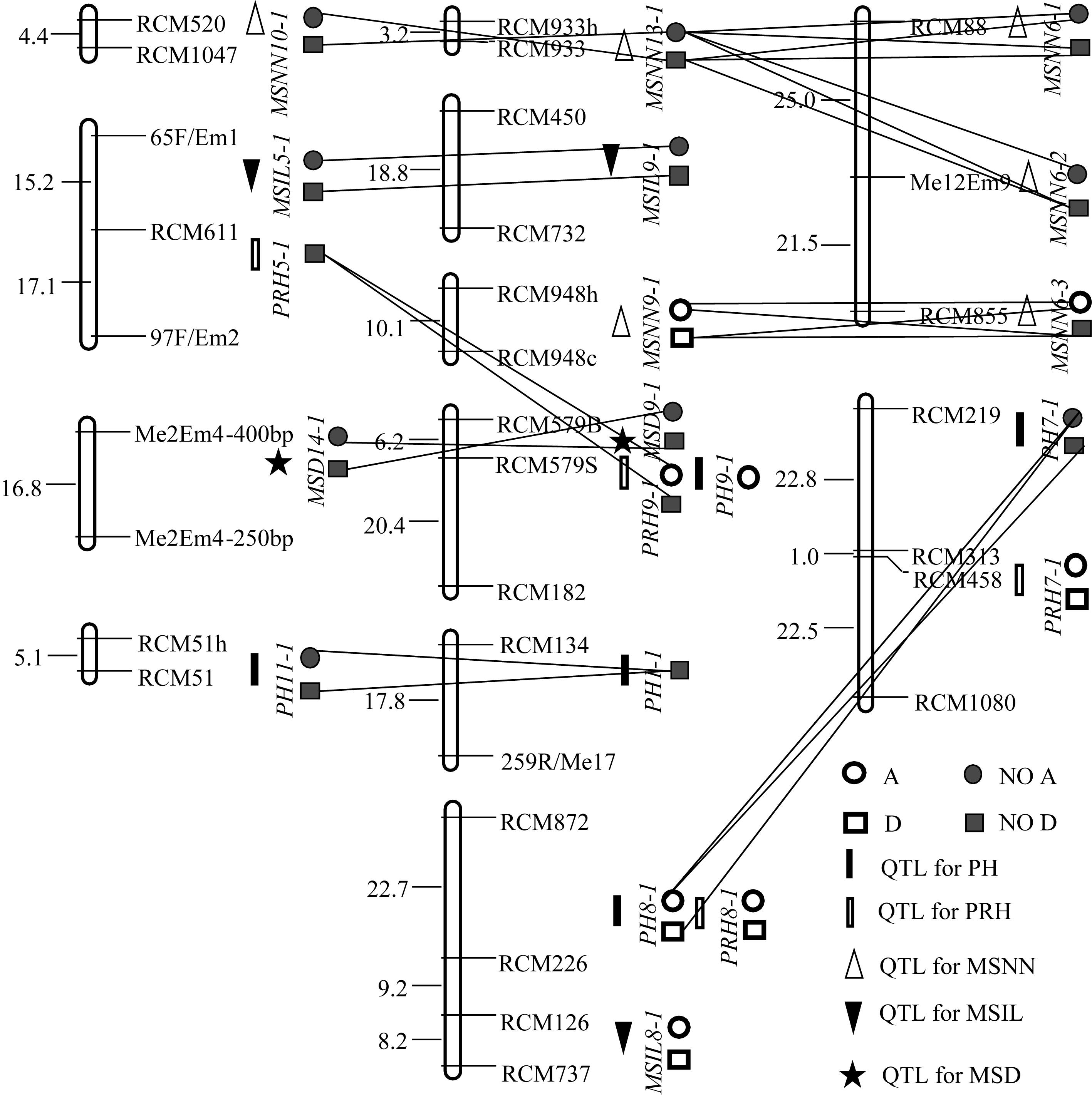 | 图1 (YC2×YF1)F2群体QTL的分布及互作A、D、NO A和NO D分别表示加性效应、显性效应、无加性效应和无显性效应; 直线相连表示2个QTL的上位性效应。Fig. 1 Distribution and interaction of QTLs in (YC2×YF1)F2 populationA, D, NO A, and NO D refer to additive effect, dominant effect, no additive effect and no dominant effect, respectively. Line joining two QTLs refers to epistatic effect. |
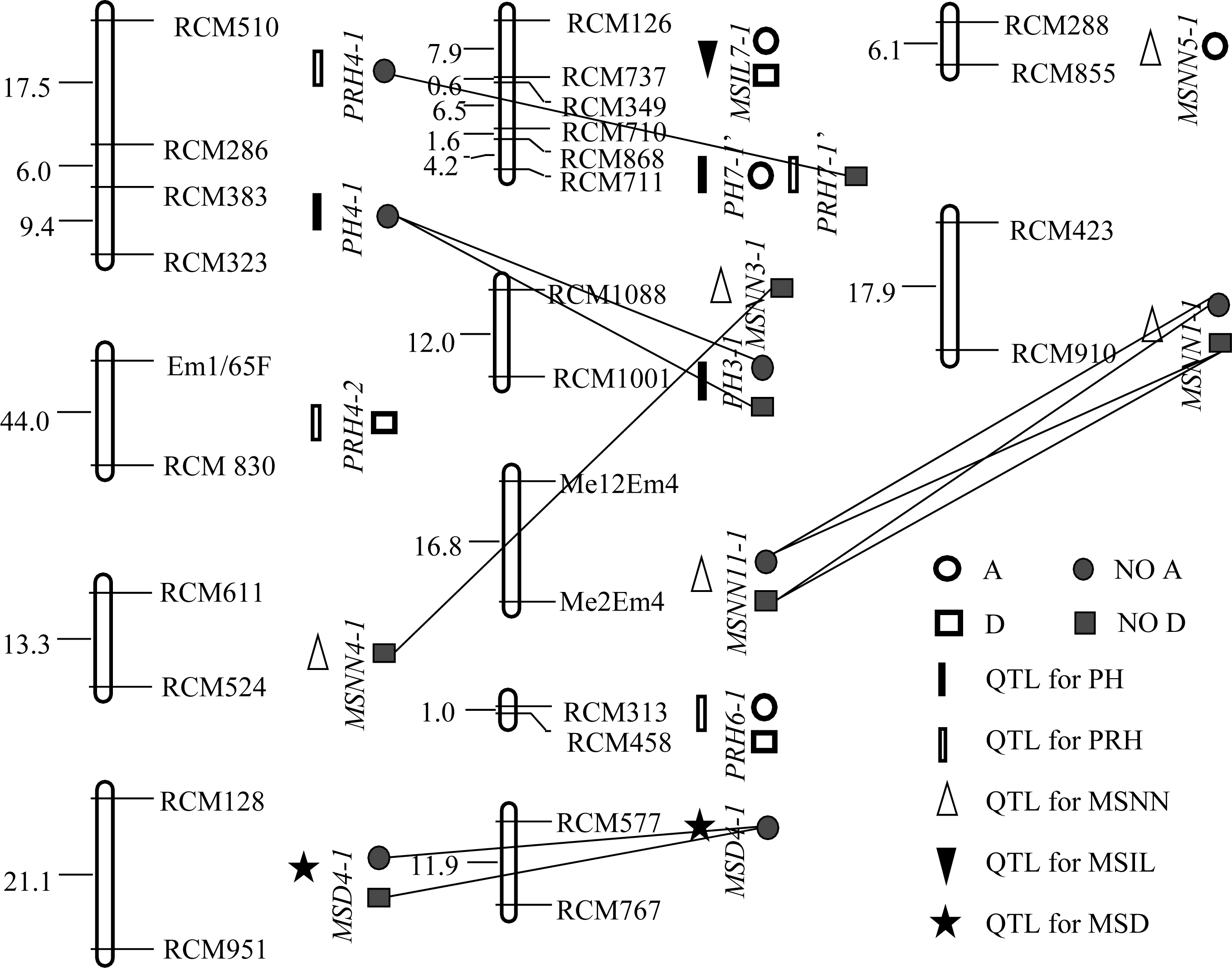 | 图2 (YC1×YF1)F2群体QTL的分布及互作Fig. 2 Distribution and interaction of QTLs in (YC1×YF1)F2 population |
产量、籽粒、株高等复杂数量性状的构成因子间存在“一因多效”、QTL连锁或重叠在小麦[ 24, 25, 26]、玉米[ 20, 27]、大豆[ 28]、黄瓜[ 21]等作物中已有报道。本试验中, 株高、主穗位高和主茎节长之间有多个主效QTL连锁乃至等位, 可见, 复杂数量性状构成因子的“一因多效”或QTL连锁在不同作物中具有普遍性。
株高QTL累计解释了2个群体45.87%、26.00%的总变异, 还有32.13%、51.00%遗传变异未被解释(广义遗传率分别为78%、77%), 有待从图谱饱和度、定位群体及定位方法本身加以改进, 但相关、回归分析结果和QTL定位结果的一致性仍然支持本文结论。
提高选择效率的关键在于抓住主基因(或主要基因)。株高、主穗位高、主茎节长都有1~2个主效QTL, 而且大多以加性效应为主, 这有利于提高选择效果。虽然主穗位高的遗传组分中, 显性效应和上位性效应也占较大比重, 增加了遗传的复杂性, 对选择效果有一定影响, 但主茎节长遗传组分中主效QTL的加性效应占绝对优势。如群体1中的 MSIL8-1和群体2中的 MSIL7-1都在RCM126~RCM737标记区间内, 区间长度分别为8.2 cM和7.9 cM, 加性效应贡献率分别为36.28%和36.30%, 是株高间接选择的关键QTL。用早期表达的、较为稳定的主茎节长对晚期表达的、更加易变的株高做出选择, 有利于提高育种效率。
主茎节数与株高不相关, 与主茎节长负相关。这一结论有助于破解两大育种难题。一是矮杆与单株产量的矛盾。试验结果显示, 后代群体的遗传倾向是少节数和长节间, 反映出自然选择的深刻烙印。较多的主茎节数往往与大穗、强生长势、叶片多、叶片厚、不早衰、宿根性等特点相联系, 但它也往往与高杆、晚熟甚至光周期敏感性相伴, 解决这一矛盾的途径就是缩短主茎节间。二是育种的方向和有效手段问题。多主茎节数和短主茎节间应该是主攻方向, 缩短主茎节间是矮化育种的突破口。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|