



{kind=link}
{kind=link}
{kind=link}
{kind=link}
赤霉素生物合成酶基因GhCPS 和GhKS 参与甲哌鎓对棉花幼苗叶片生长的控制
[王丽1, 2 , 张明才1 , 杜明伟1 , 田晓莉1 , 李召虎1, *  ]
]
]
|
|


室内盆栽欣抗4, 在棉花幼苗第3片真叶完全展平时(第4叶未展开)叶面喷施甲哌鎓(DPC), 研究DPC对棉花幼苗叶片生长的控制与赤霉素(GA)合成早期关键酶柯巴基焦磷酸合酶(CPS)和内根-贝壳杉烯合酶(KS)基因表达的关系。结果表明, DPC处理显著减小棉花幼苗第3和第4叶的叶面积, 第4叶叶面积受控制程度较第3叶大; 80 mg L-1DPC处理的棉花幼苗第3和第4叶中GA4含量分别于处理后4 d和4~6 d显著低于对照; 与对照相比, 80 mg L-1DPC处理的棉花幼苗第3叶中
Ent-copalyl diphosphate synthase (CPS) and ent-kaurene synthase (KS) are the key enzymes involved in the early steps of gibberellin (GA) biosynthesis. This paper aimed at elucidating whether the action of mepiquat chloride (DPC) on leaf growth was related to the expression levels of
甲哌鎓(mepiquat chloride, N, N-二甲基哌啶鎓氯化物, DPC)是世界棉花生产上应用最广泛的植物生长延缓剂, 可降低株高、缩短节间、减小叶面积[ 1, 2, 3], 塑造较理想的棉花群体形态。DPC还可增加棉花叶片的叶绿素含量[ 4], 增强其净光合速率[ 5], 提高光能利用效率[ 6], 有利于提高棉花的产量和品质。
DPC属于鎓类植物生长延缓剂, 矮壮素(CCC)和AMO-1618同属此类延缓剂[ 7]。研究表明CCC和AMO-1618可抑制赤霉素(GA)合成早期关键酶柯巴基焦磷酸合酶(ent-copalyl diphosphate synthase, CPS)和内根-贝壳杉烯合酶(ent-kaurene synthase, KS)的活性[ 8, 9]。Rademacher[ 7]认为DPC与CCC和AMO- 1618具有相似的化学结构, 其靶酶也是CPS和KS。CPS活性的调控主要取决于 CPS基因的表达水平, 该基因主要受植物生长发育的调控[ 10, 11]。 CPS基因若完全突变, 植物不能产生任何GA, 导致植物种子不能萌发、植株矮化和雄性不育[ 12, 13]。KS是GA合成途径的第2个酶, 编码KS的基因突变也严重影响植株的发育[ 12, 13]。CPS和KS是否在转录水平上受DPC的调控, 参与DPC对棉花幼苗叶片生长的控制, 目前尚无报道。本研究利用荧光定量PCR技术, 研究了DPC对棉花幼苗刚展平叶片和未展开的心叶 GhCPS和 GhKS表达的动态调控, 以探讨其表达模式与DPC控制棉花幼苗叶片生长的关系。
陆地棉欣抗4由河北省河间市国欣农村技术服务总会提供; DPC (纯度97%)购自河北国欣诺农生物技术有限公司。
采用盆栽试验, 选取饱满且完好的种子浸种后点播在装有营养土∶蛭石(w/w)=1∶1的花盆(13 cm直径×13 cm高)中, 每盆点4粒种子, 待棉花幼苗第1片真叶出现时, 每盆仅留1株。培养在中国农业大学转基因基地的人工气候室中, 光照强度为400 μmol m-2 s-1, 光/暗周期为14 h/10 h, 昼/夜温度为28℃/20℃。
试验1设置5个处理, 即清水(对照)和DPC (40、80、160和320 mg L-1), 每处理3个重复, 每个重复15盆幼苗。待棉花幼苗的第3片真叶刚展平且第4片真叶未展开时于全株叶面均匀喷施每株8 mL。处 理后10 d测定各处理组第3和第4叶的叶面积。
试验2设置2个处理, 即清水(对照)和80 mg L-1DPC, 每处理3个重复, 每个重复20盆幼苗。处理方法同试验1。于处理后0、1、2、4、6、8、10 d取第3和第4叶片, 用液氮速冻并存于-80℃, 用于RNA提取; 于处理后0、2、4、6和10 d取第3和第4叶片, 用液氮速冻并存于-80℃, 用于GA含量的测定; 于处理后0、2、4、6、8和10 d量取第3和第4叶片的主叶脉长度; 处理后10 d测量各处理第3和第4叶的叶面积。
1.3.1 叶面积的测定 测定处理后10 d叶面积采用LI-3100叶面积仪(LI-COR Inc., Lincoln, NE)。根据处理后10 d主叶脉长度和叶面积建立方程 A = 0.07 L2+8.1328 L-16.902 ( R2= 0.9494) ( A为叶面积; L为主叶脉长度), 据此计算处理后0、2、4、6和8 d第3和第4叶的叶面积[ 14]。
1.3.2 GA4含量的测定 采用间接酶联免疫吸附法(ELISA)。所用单克隆抗体由中国农业大学化控室提供, 参照何钟佩[ 15]方法配制试剂与测定。
1.3.3 RNA的提取及实时荧光定量RT-PCR 采用植物RNA提取试剂盒(艾德莱, 北京)提取棉花幼苗叶片总RNA, 经DNase I (艾德莱, 北京)消化后, 用逆转录酶(M-MLV Reverse Transcriptase, Promega)合成cDNA。采用SYBR green II荧光染料(TaKaRa, Japan)法进行荧光定量RT-PCR分析。以cDNA为模板, 用棉花内参基因 GhUBQ7和基因特异性引物(表1)和SYBR Premier Ex Taq mix (TaKaRa, Japan)进行PCR, 扩增条件为95℃ 30 s; 95℃ 5 s, 60℃ 35 s, 40个循环。采用2-ΔΔCt分析基因相对表达量[ 16]。
| 表1 引物序列 Table 1 Primer sequences used in this study |
独立重复各试验3次, 各次结果趋势一致, 选用其中具代表性的一次。采用SPSS17.0软件进行数据统计分析, ANOVA法进行差异显著性分析( P=0.05), Duncan’s法进行多重比较。
不同浓度DPC处理棉花幼苗后10 d, 第3和第4叶的叶面积均显著减小(图1)。与对照相比, 第3叶减小13%~26%, 第4叶减小28%~60%。与80 mg L-1DPC处理组相比, 160 mg L-1和320 mg L-1DPC处理的第3叶无显著变化, 而第4叶存在显著差异。
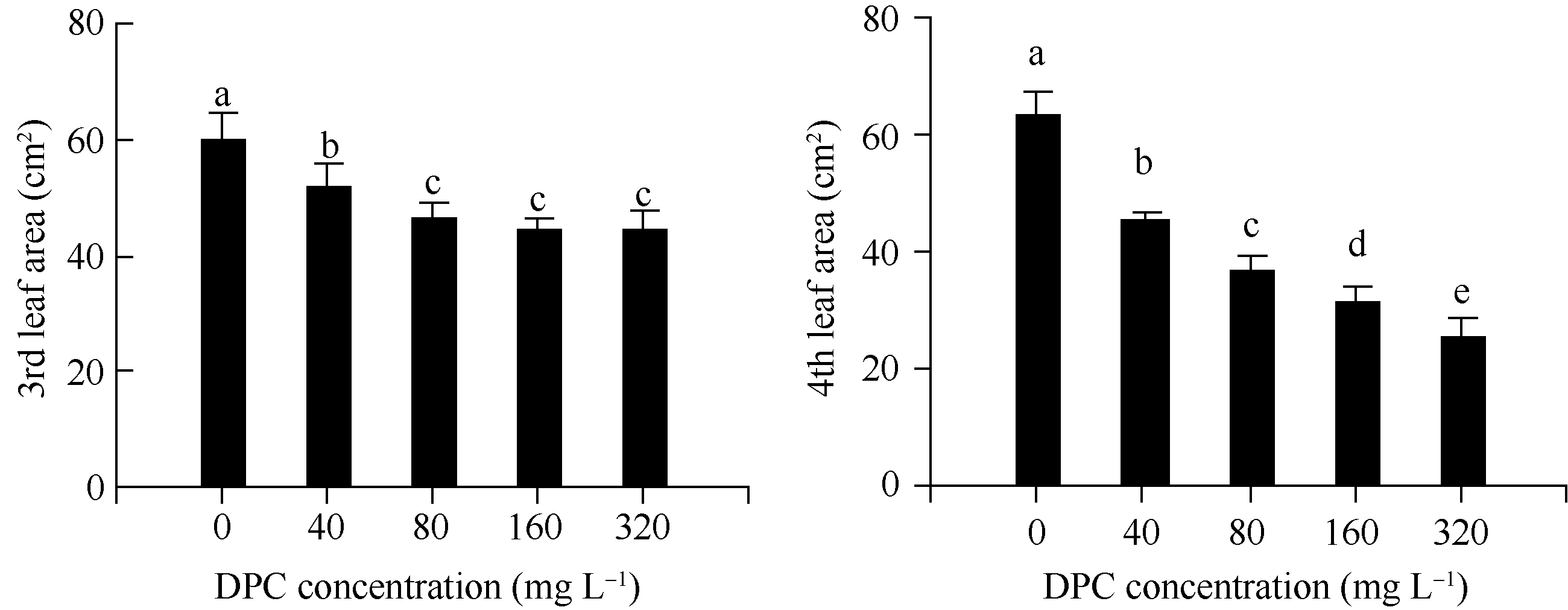 | 图1 不同浓度DPC对棉花幼苗第3和第4叶叶面积的影响DPC: 甲哌鎓; 数据为平均值± SE; 标以不同字母的柱值在0.05水平上差异显著。Fig. 1 Effect of different concentrations of mepiquat chloride on the 3rd and 4th leaf area in cotton seedlingsDPC: mepiquat chloride; data are means± SE; Bars superscripted by different letters are significantly different ( P<0.05). |
如图2所示, 与对照相比, 80 mg L-1DPC处理后4 d, 棉花幼苗第3和第4叶的叶面积均开始显著减小, 处理后4、6、8和10 d第3叶的叶面积分别减小19%、21%、26%和23%, 第4叶的叶面积分别减小51%、53%、51%和44%。与不同浓度DPC处理结果一致, 第4叶的叶面积受DPC抑制程度较第3叶大。
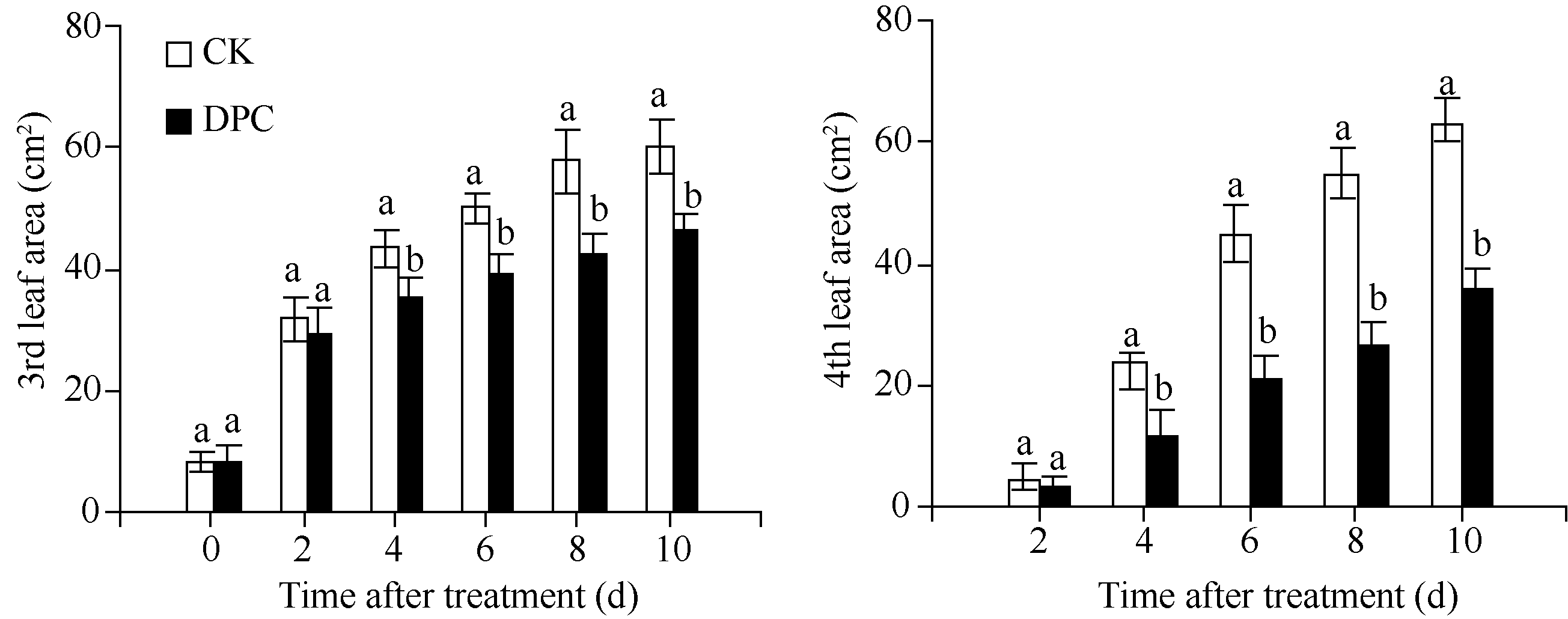 | 图2 DPC对棉花幼苗第3和第4叶叶面积扩展动态的调控DPC: 80 mg L-1甲哌鎓; 数据为平均值± SE; 标以不同字母的柱值在0.05水平上差异显著。Fig. 2 Regulation of mepiquat chloride on the expansion of 3rd and 4th leaf area in cotton seedlingsDPC: 80 mg L-1 mepiquat chloride; data are means± SE; Bars superscripted by different letters are significantly different ( P<0.05). |
随着第3叶片的生长, 对照组中GA4含量先升后降, 峰值位于处理后4 d; DPC处理组中GA4含量的变化与对照组相似, 但峰值降低。80 mg L-1 DPC处理后4 d第3叶中GA4含量显著低于对照, 降低了29% (图3-左)。随着第4叶片的生长, 对照组中GA4的含量先升后降, 峰值位于处理后6 d; DPC处理组中GA4含量在处理后2 d增加, 之后趋于稳定, 无峰值出现。与对照相比, 80 mg L-1DPC处理后4 d和6 d棉花幼苗第4叶中GA4的含量分别降低了31%和39% (图3-右)。
 | 图3 DPC对棉花幼苗第3和第4叶GA4含量的动态调控DPC: 80 mgL-1甲哌鎓; 数据为平均值±SE; 标以不同字母的柱值在0.05水平上差异显著。Fig. 3 Regulation of mepiquat chloride on GA4 content in the 3rd and 4th leaves of cotton seedlingsDPC: 80 mg L-1 mepiquat chloride; data are means± SE; Bars superscripted by different letters are significantly different ( P<0.05). |
随着第3叶片的生长, 对照组中 GhCPS和 GhKS的表达先增后降, 峰值位于处理后2~4 d; DPC处理组中 GhCPS和 GhKS的表达在处理后1~2 d降低, 4 d开始增加, 峰值分别位于处理后6 d和8 d (图4)。与对照相比, 80 mg L-1 DPC处理后1 d, 棉花幼苗第3叶中 GhCPS和 GhKS表达显著下调, 在处理后1~4 d, GhC PS的表达下调18%~59%, GhKS的表达下调39%~63%。随着第4叶片的生长, 对照组中 GhCPS和 GhKS的表达先增后降, 6 d时出现峰值; DPC处理组中 GhCPS和 GhKS的表达在处理后1~6 d明显降低, 8 d时开始增加并出现峰值(图4)。与对照相比, 80 mg L-1 DPC处理后1 d, 棉花幼苗第4叶中 GhCPS和 GhKS的表达显著下调, 在处理后1~6 d, GhCPS的表达下调39%~71%, GhKS的表达下调23%~67%。
 | 图4 DPC对棉花幼苗第3和第4叶 GhCPS和 GhKS表达的动态调控DPC: 80 mg L-1甲哌鎓; 数据为平均值± SE; 标以不同字母的柱值在0.05水平上差异显著。Fig. 4 Regulation of mepiquat chloride on expressions of GhCPS and GhKS in the 3rd and 4th leaves in cotton seedlingsDPC: 80 mg L-1 mepiquat chloride; data are means± SE; Bars superscripted by different letters are significantly different ( P<0.05). |
DPC是一种水溶性的有机分子, 主要通过叶面喷施, 由叶片吸收, 在植株中转运和重新分配, 控制植物的生长。研究表明DPC可减小棉花的叶面积, 且该作用与DPC的浓度和棉花的发育时期有关[ 3, 17, 18], 棉花叶片的生长速率随DPC浓度的增加呈线性减小[ 4]。我们研究发现不同浓度DPC处理均显著减小
棉花幼苗第3和第4叶的叶面积, 这与前人研究结果是一致的。陈吟等[ 19]用DPC处理七叶期棉花幼苗的第5叶(自茎基部向上), 发现DPC可在棉花幼苗内转运, 且处理叶上部的茎和顶部叶片(第7叶)中DPC的积累速率和积累量明显高于处理叶下部的茎和叶片, 认为DPC优先向顶部生长旺盛的组织运输。DPC处理时未展开叶片即第4叶的生长比已展平叶片第3叶受DPC控制程度大, 可能与其较快地吸收DPC或积累较多的DPC有关。
GA参与植物生长发育的许多过程, 如促进种子萌发、节间伸长、叶片生长、花发生及果实与种子发育等[ 20, 21]。GA在调控叶片细胞分裂、细胞伸长和器官形成中起重要作用[ 22]。研究发现一些植物矮化突变体的叶片生长较野生型缓慢, 主要与其对GA敏感性降低或内源活性GA含量降低有关[ 23]。Kang等[ 24]通过叶面喷施调环酸钙, 发现大白菜的叶面积显著减小, 叶片中GA1和GA4含量显著降低。DPC处理显著降低棉花幼苗第3和第4叶中GA4含量, 表明DPC对棉花幼苗叶片生长控制与内源活性GA4含量降低有关, 其中第4叶生长受DPC控制程度较大与GA4含量降低及持续时间长有关。Otani等[ 25]通过过表达GA2氧化酶基因获得百合科油点草属矮化植株, 其叶片的扩展程度与叶片内活性GA4含量密切相关。
GA合成的前体为牻牛儿基牻牛儿基焦磷酸(geranylgeranyl diphosphate, GGDP), CPS和KS可催化GGDP形成内根-贝壳杉烯。CPS是正式进入GA合成途径的第一个限速酶, 其基因的表达主要受生长发育的调控, 在快速生长的组织或组织快速生长阶段表达水平较高[ 10, 11]。 KS的表达是组成型的, 并在生长的组织中表达量较高, 如在叶尖和发育中的子叶[ 26]。在豌豆萌发与南瓜和拟南芥幼苗生长过程中, 子叶的 CPS表达峰值出现在其快速伸展阶段[ 10, 27, 28]。本研究对照组中棉花幼苗在第3和第4叶生长过程中, GhCPS和 GhKS表达均有峰值出现, 但DPC处理推迟了叶片中 GhCPS和 GhKS表达峰值的出现, 这说明DPC处理延缓了叶片的快速生长。
DPC显著抑制第3和第4叶中 GhCPS和 GhKS的表达, 表明DPC处理后棉花幼苗叶片中GA4的含量降低与 GhCPS和 GhKS的表达下调有关。植物体中编码CPS的基因若完全突变, 植物不能产生GA, 表现为GA缺陷表型, 即种子不能萌发、植株矮化、叶片变小等[ 12, 13, 29]。在植物 KS突变体也出现类似的表型[ 12, 13]。我们发现DPC处理植株叶片中 GhCPS和 GhKS的表达并未完全被抑制, 叶片表现出缓慢生长、叶片变小。不同叶龄叶片生长受到抑制程度存在显著差异, 如与第3叶相比, 第4叶中 GhCPS和 GhKS表达受DPC作用下调时间较长, GA4含量维持低水平时间较长, 叶片生长受DPC控制程度较大。
生产上棉花应用DPC后, 除叶片变小外, 植株矮化、节间缩短、叶色变深、叶片变厚、叶绿素含量增加和叶片光合速率提高[ 1, 2, 3, 4, 6], 与GA缺陷突变体的表型相似[ 12, 13]。本试验结果表明DPC可抑制棉花幼苗叶片 GhCPS和 GhKS的表达, 引起GA含量的降低, 从而抑制叶片生长。因此生产上棉花应用DPC后所表现的生长抑制可能与 GhCPS和 GhKS的表达下调引起GA含量的降低有关。但DPC对棉花幼苗叶片生长的抑制是由于叶肉细胞变小还是数目减少或两者兼有, 仍需进一步研究。此外, 与植物生长密切相关的其他激素如细胞分裂素、生长素和油菜素内酯等是否也参与DPC对棉花幼苗生长的抑制, 仍需进一步探讨。
DPC控制棉花生长与 GhCPS和 GhKS的表达下调引起的GA含量降低有关; DPC处理后棉花幼苗较幼嫩叶片 GhCPS和 GhKS的下调表达幅度较大且时间较长, 表明幼嫩部位是DPC作用的主要部位。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|