


{kind=link}
{kind=link}
{kind=link}
硬粒小麦品种Lpx-B1 位点等位变异的分子鉴定及其脂肪氧化酶活性
[张福彦1, 2, * , 尚晓丽1, * , 吴培培1 , 宋双1 , 陈锋1, *  , 崔党群
, 崔党群1, * ]
, 崔党群]
|
|


硬粒小麦籽粒中脂肪氧化酶(LOX)与硬粒小麦面制品的加工品质关系密切相关, 而
Lipoxygenase (LOX) has a close relationship to the processing quality of flour products in durum wheat (
脂肪氧化酶(lipoxygenase, LOX)是一类不含血红素铁的蛋白质, 广泛存在于各种类型的植物中[ 1, 2]。它在植物的生长、发育、衰老、脂质过氧化作用、光合作用及抗逆性等方面起重要作用[ 2]。小麦籽粒中的LOX含量很低, 但其在酶促脂肪氧化过程中产生的活性氧, 不仅能通过降解色素类物质影响面粉及面条、馒头和面包等面制品颜色[ 3], 且能有效改善面制品的营养品质和加工品质[ 4, 5, 6]。因而, 小麦脂肪氧化酶活性及其在品质改良中的重要作用已成为国内外学者研究的热点之一。
小麦籽粒LOX活性是一个受多基因控制的复杂数量性状, 受基因型和环境及其互作的显著影响, 且基因型的效应大于环境及互作效应[ 7, 8]。Leenhardt等[ 9]发现普通小麦籽粒中的LOX活性高于硬粒小麦和一粒小麦, 分别是其2.5倍和7.5倍, 表明小麦类型也是影响LOX活性的因素之一。Hart和Langston[ 10]利用中国春缺体-四体系对LOX同工酶定位表明, Lpx-A1、 Lpx-B1和 Lpx-D1分别位于4AL、4BL和4DS染色体, 而 Lpx-A2、 Lpx-B2和 Lpx-D2分别位于5AL、5BL和5DL, 推测小麦LOX基因主要位于第4和第5同源群。Hsieh等[ 11]从硬粒小麦胚乳中分离出3个与LOX活性有关的同工酶, 分别命名为lipoxygeanses-1 (L-1)、lipoxygeanses-2 (L-2)和lipoxygeanses-3 (L-3)。硬粒小麦中, 脂肪氧化酶由 Lpx编码, 其主效基因分别被定位在4A[ 12]、4B[ 12, 13, 14, 15, 16, 17, 18]、4D[ 18, 19]和5D[ 19]染色体上, Lpx-B1位点的等位变异对LOX活性有显著影响[ 3]。Carrera等[ 14]分析7个硬粒小麦品种的 Lpx-B1基因及其对LOX活性的影响发现, Lpx-B1位点部分片段重叠和 Lpx-B1.1基因部分片段缺失的现象, 并将这2个基因分别命名为 Lpx-B1.1(DQ474240)和 Lpx-B1.2(DQ474241), 且后者与LOX活性降低密切相关。此外, Verlotta等[ 15]还报道了一个控制LOX活性的新基因, 将其命名为 Lpx-B1.3(HM126469), 同时认为 Lpx-B1.2和 Lpx-B1.3基因可能是一对等位基因; Lpx-B1.1位点存在3个等位变异, 分别命名为 Lpx-B1.1a、 Lpx-B1.1b和 L px-B1.1c, 并开发了相应的功能标记。
硬粒小麦是世界第二大麦类作物, 种植面积占世界小麦总面积的10%左右, 其蛋白质和面筋含量较高, 富含大量胡萝卜素, 具有较高的营养价值, 在欧洲, 拉丁美洲的许多国家很受欢迎。硬粒小麦在储藏和加工过程中脂肪氧化酶的氧化降解会导致胡萝卜素和类胡萝卜素大量损失, 降低部分营养价值[ 20, 21]。郑文寅等[ 22]分析了小麦全麦粉的色泽性状与LOX和类胡萝卜素含量之间的关系, 结果表明, LOX活性与类胡萝卜素含量呈极显著负相关, 而与全麦粉的白度、亮度和黄度无显著相关。本研究旨在明确硬粒小麦籽粒中 Lpx-B1基因的等位变异与LOX活性之间的关系, 为利用分子标记选择相应LOX活性的硬粒小麦品种提供理论依据。
167份硬粒小麦(意大利博洛尼亚大学农业环境科技学院的Roberto Tuberosa教授提供)来自不同国家和地区, 为历史品种和当前主栽品种, 其中意大利47份、国际玉米小麦改良中心(CIMMYT) 12份、叙利亚80份、西班牙17份、摩洛哥11份。2010—2011年生长季种植于河南农业大学科教示范园区, 2次重复, 常规田间管理。所有材料在当地均能正常成熟, 且未发现有倒伏和穗发芽现象。
取完熟后籽粒, 用分光光度计法[ 16]测定籽粒中LOX活性, 每份材料3次重复, 求平均值。操作步骤主要有LOX酶的提取、亚油酸底物的配制(现用现配)、分光光度计检测。所用试剂包括40 mmol L-1 PBS缓冲液(pH 6.8)、亚油酸(Sigma, L1376)、Tween 20、1 mol L-1 NaOH、0.1 mol L-1 NaOH, 以及ddH2O。使用UV-2600型紫外可见分光光度计(尤尼柯上海仪器有限公司)在234 nm处用1 cm光程石英比色皿测定共轭过氧化物的吸光度∆A, 以每毫克总蛋白每分钟吸光值增加0.001为一个酶活性单位(AU min-1 g-1), 用经过高温煮沸LOX粗酶液底物做对照。
从每份试验材料取3粒有代表性的种子, 用锤子砸碎后放入2.0 mL离心管, 参照单籽粒种子DNA快速提取方法[ 23]提取DNA, 但用SLS (十二烷基肌氨酸钠)代替SDS (十二烷基硫酸钠)。利用Verlotta等[ 15]在硬粒小麦开发的共显性标记 Lpx-B1.1a/b以及显性标记 Lpx-B1.1c、 Lpx-B1.2和 Lpx-B1.3 (表1)检测参试品种4B染色体上 Lpx-B1位点等位变异。所有引物均由生工生物工程(上海)有限公司合成。
在ABI 9700或MJ Research PTC-200中进行PCR扩增, 扩增体系为25 μL, 包含2 μL基因组DNA (50 ng μL-1)、 0.5 μL dNTP (10 mmol L-1)、各0.4 μL上、下游引物 (10 pmol)、2.5 μL 10× buffer (20 mmol L-1 Tris-HCl pH 8.4, 25 mmol L-1 KCl, 1.5 mmol L-1 MgCl2)、0.4 μL Taq DNA聚合酶(2.5 U μL-1)。以上试剂均购自TIANGEN公司。PCR程序为95℃预变性5 min; 95℃变性30 s, 58~64℃退火30 s (不同引物退火温度见表1), 72℃延伸1 min, 35个循环; 72℃延伸10 min, 4℃保存。
| 表1 硬粒小麦中检测 Lpx-B1基因等位变异的引物 Table 1 Specific primers for identification of Lpx-B1 alleles in durum wheat |
在1×TBE的缓冲液中, 用含有溴化乙锭(EB)的1.0% (w/v)琼脂糖凝胶电泳分离PCR扩增产物, 150 V电压下电泳15 min, 最后在凝胶成像系统中扫描观察, 并对电泳结果统计分析。 Lpx-B1.1a和 Lpx-B1.1b等位变异片段差别较小, 只相差74 bp, 故采用2.0% (w/v)琼脂糖凝胶电泳分离检测, 150 V电压下电泳30 min后扫描观察。
利用SPSS 19.0软件分析LOX活性平均值和标准差等参数, 用LSR方法比较不同基因型组合的LOX活性差异显著性。
首先利用 Lpx-B1.1a/b标记检测, 扩增产物为1312 bp和1238 bp两种带型, 分别对应 Lpx-B1.1a和 Lpx-B1.1b等位变异(图1); 部分品种中没有检测到扩增产物, 可能为 Lpx-B1.1c等位变异, 进一步用 Lpx-B1.1c的特异引物检测, 结果Cannizzo、Claudio、Lesina和Norba等7个材料中扩增出1558 bp的条带(图2), 确认为 Lpx-B1.1c变异类型。
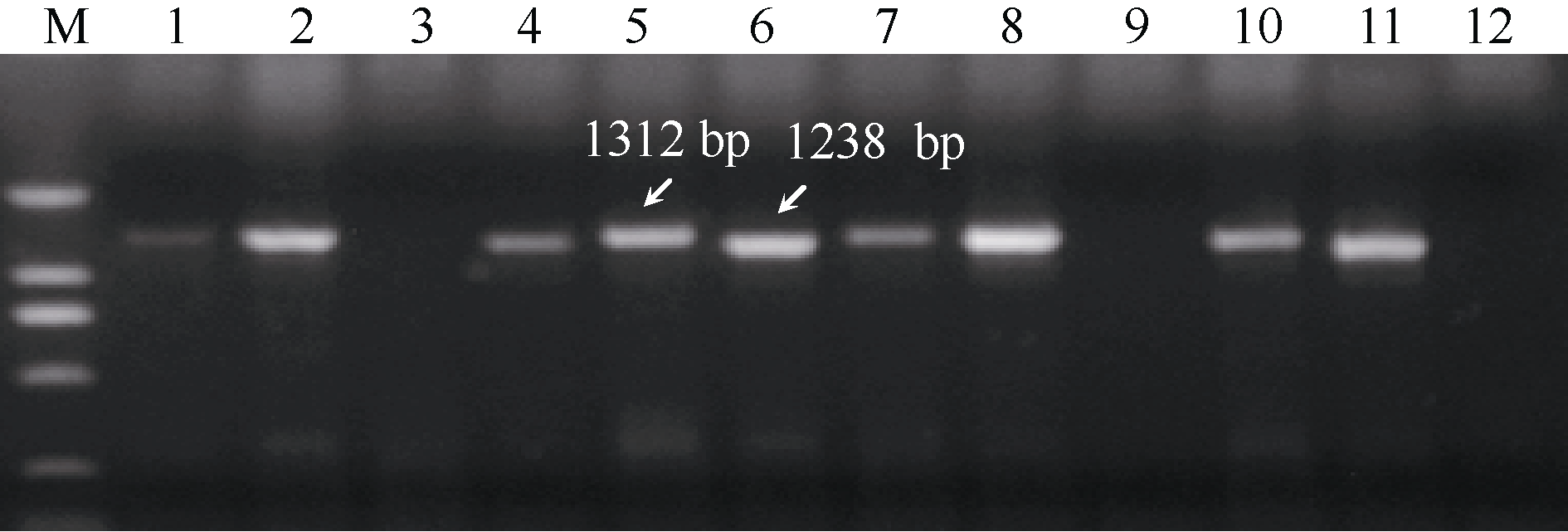 | 图1 Lpx-B1.1a/b引物在部分硬粒小麦品种中的扩增结果Fig. 1 Amplification profile of Lpx-B1.1a/b primers in partial varieties of durum wheatM: DL2000; 1: Meridiano; 2: Appio; 3: Cannizzo; 4: Trinakria; 5: Bolenga; 6: Appulo; 7: Cappelli; 8: Torrebianca; 9: Mongibello; 10: Roqueno; 11: Quadrato; 12: Colosseo. |
 | 图2 Lpx-B1.1c引物在部分硬粒小麦品种中的扩增结果Fig. 2 Amplification profile of Lpx-B1.1c primers in partial varieties of durum wheatM: DL2000; 1: Cannizzo; 2: Claudio; 3: Lesina; 4: West Bred Turbo; 5: Roqueno; 6: Torrebianca; 7: Bravadur; 8: Meridiano; 9: Norba; 10: Mongibello; 11: Colosseo; 12: Younes-1. |
在 Lpx-B1位点共发现5种变异类型, 分别为 Lpx-B1.1a、 Lpx-B1.1b、 Lpx-B1.1c、 Lpx-B1.2和 Lpx- B1.3。 L px-B1.1位点有3种等位变异, 其中 Lpx-B1.1a类型品种有92份, 占55.1%, Lpx-B1.1b和 Lpx-B1.1c类型的品种分别为13份和62份, 占7.8%和37.1%。此外, 没有发现同一品种兼有 Lpx-B1.2和 Lpx-B1.32个等位变异, 即二者总是互补出现(表2), 146份品种为 Lpx-B1.2变异类型(占87.4%), 而另外21份品种则为 Lpx-B1.3类型(占12.6%)。
Lpx-B1位点基因型因品种来源不同而异。来自叙利亚和意大利的品种, Lpx-B1位点变异类型较为丰富, 拥有所有的5种变异类型; 而在来自CIMMYT、摩洛哥和西班牙的品种中, 没有发现 Lpx-B1.1b和 Lpx-B1.3两种类型。80份叙利亚材料中, Lpx-B1.1位点 Lpx-B1.1a、 Lpx-B1.1b和 Lpx-B1.1c类型分别有46、8和26份; 而在另一位点, 有65份为 Lpx-B1.2类型, 15份为 Lpx-B1.3类型。47份意大利材料中, Lpx-B1.1位点有19份为 Lpx-B1.1a类型, 5份为 Lpx-B1.1b类型, 23份为 Lpx-B1.1c类型; 另一位点 Lpx-B1.2类型有41份, Lpx-B1.3类型有6份(表3)。结果表明, Lpx-B1.1a、 Lpx-B1.1c和 Lpx-B1.2三种基因型在硬粒小麦中占主导地位。
方差分析结果表明, 硬粒小麦籽粒中 Lpx-B1位点不同变异类型间LOX活性有显著差异(图3)。 Lpx-B1.1b和 Lpx-B1.3类型品种的LOX活性最高, 分别为5.74 AU min-1 g-1和5.04 AU min-1 g-1; Lpx-B1.1c变异类型的LOX活性最小, 仅为0.92 AU min-1 g-1; 其他2类品种的LOX活性分别为2.50 AU min-1 g-1( Lpx-B1 .1a)和1.75 AU min-1 g-1 ( Lpx-B1.2)。
| 表2 部分硬粒小麦品种的LOX活性和 Lpx-B1基因型鉴定 Table 2 Lpx-B1 genotypes and LOX activities in part of durum wheat varieties |
| 表3 不同国家硬粒小麦种质的 Lpx-B1位点基因型分布 Table 3 Distribution of different Lpx-B1alleles in durum wheat varieties from various countries |
167个品种在 Lpx-B1位点共有6种基因型组合, 分别为 Lpx-B1.1a/Lpx-B1.2、 Lpx-B1.1b/Lpx-B1.2、 Lpx-B1.1c/Lpx-B1.2、 Lpx-B1.1a/Lpx-B1.3、 Lpx-B1.1b/ Lpx-B1.3和 Lpx-B1.1c/Lpx-B1.3。其中, Lpx-B1.1b/ Lpx-B1.3基因型组合为高LOX活性类型, 有11个品种(6.6%), 其LOX活性平均值高达6.03 AU min-1 g-1; Lpx-B1.1a/Lpx-B1.3和 Lpx-B1.1b/Lpx-B1.2基因型组合为偏高类型, 分别有9个(5.4%)和2个(1.2%)品种, 其LOX活性分别为4.27 AU min-1 g-1和4.17 AU min-1 g-1; Lpx-B1.1a/Lpx-B1.2基因型组合为偏低类型, 是参试品种中的最主要的类型, 包括83个品种(49.7%), 其LOX活性为2.30 AU min-1 g-1; Lpx-B1.1c/Lpx-B1.2基因型组合属于低LOX活性类型, 共有61个品种(36.5%), 其平均LOX活性最低, 仅为0.92 AU min-1 g-1; 另外, 仅发现1份 Lpx-B1.1c/Lpx-B1.3组合类型, LOX活性为1.02AU min-1 g-1, 也属于低LOX活性类型(表4)。
 | 图3 硬粒小麦中 Lpx-B1位点不同变异类型的LOX活性LOX活性为同类品种的平均值及标准差; 柱形上面的不同字母表示不同变异类型间有显著性差异( P<0.05)。Fig. 3 LOX activities in different Lpx-B1 genotypes of durum wheatLOX activity is the mean of each variety group, and the error bar shows the standard error; Different letters above columns indicate significant difference among different types of variation at P < 0.05. |
| 表4 硬粒小麦中 Lpx-B1位点不同基因型组合LOX活性比较 Table 4 Comparison of LOX activities of durum wheat varieties with different genotype combinations |
面粉或面制品的表观色泽是评价小麦品质的重要指标之一。LOX对面粉色泽有很大影响, 且对小麦品质性状具有双重效应。籽粒中LOX会偶联氧化小麦中的类胡萝卜素, 取代化学漂白剂使小麦粉变白, 提高面粉或面制品的白度; 但是过高的LOX活性则会破坏小麦籽粒中的黄色素, 使小麦粉过白而丧失许多营养成分, 降低麦类食品的营养价值[ 3, 6, 22]。研究表明, LOX与小麦籽粒、面粉及其面制品的储藏特性的关系密切, 降低LOX活性有利于延长硬粒小麦籽粒、通心粉等食品的保存期, 从而提高产品的附加值[ 24], 降低LOX活性也是长期保存种子的重要方法之一。可见, 低LOX活性可以有效地减轻脂质的氧化反应, 减少籽粒的氧化变质, 延长其储藏期, 从而减少粮食的损失[ 6, 25]。本研究在分析硬粒小麦品种籽粒LOX活性及其基因型分布特点的同时, 也试图筛选出低LOX活性遗传材料, 为今后综合性状评价和硬粒小麦品质育种提供基础信息和种质资源储备。
硬粒小麦中, LOX为受多基因控制的数量性状, Lpx-B1位点变异类型是导致LOX活性变化的重要原因。我们在167份硬粒小麦材料中发现LOX活性变异范围为0.20~7.98 AU min-1 g-1, 品种间差异较大。Verlotta等[ 15]把 Lpx-B1不同基因型组合分成3个单体型, 即单体型I ( Lpx-B1.1b/Lpx-B1.3)、单体型II ( Lpx- B1.1a/Lpx-B1.2)和单体型III ( Lpx-B1.1c/Lpx-B1.2), 其LOX活性平均值分别为4.60、2.59和0.12 AU min-1 g-1。本研究结果与此类似, LOX活性最高和最低的也分别是 Lpx-B1.1b/Lpx-B1.3和 Lpx-B1.1c/Lpx-B1.2组合类型, 平均LOX活性依次是6.03 AU min-1 g-1和0.92 AU min-1 g-1, 同时本研究结果也支持 Lpx-B1位点等位变异能够引起LOX活性的显著变化的观点。在硬粒小麦主要消费国家, 低LOX活性的硬粒小麦更受欢迎。研究表明, LOX活性与蛋白质、淀粉、湿面筋等大多数品质参数呈负相关, 且能有效增加面筋强度, 导致面团延展性降低[ 26, 27]。与普通小麦相比, 硬粒小麦中LOX活性更低, 同时由于硬粒小麦中的蛋白质和抗氧化剂化合物也显著高于普通小麦, 因此认为硬粒小麦比普通小麦具有更高的营养价值[ 28]。
在对小麦籽粒LOX活性相关基因的定位研究中, 发现一些与目标性状紧密连锁的SSR标记, 如 Xbcd1262、 Xksud2a、 Xwmc312和 Xgwm251[ 13, 29], 但这些标记不在基因内部, 不能与目的基因共分离, 因而预测基因型的准确性较低, 在实际应用中存在局限性。功能标记是根据基因内部引起表型性状变异的多态性序列而开发出的一种分子标记, 利用其进行辅助选择育种具有高效、准确、适用性广等特点[ 30, 31]。目前, 功能性标记已广泛用于小麦品质和农艺性状以及抗病性鉴定等领域, 是小麦育种的重要辅助手段之一[ 31]。本试验用功能标记对来自不同国家和地区的167份硬粒小麦品种进行 Lpx-B1位点的分子鉴定, 发现主要为 Lpx-B1.1a、 Lpx-B1.1c和 Lpx-B1.2基因型, 与Verlotta等[ 15]利用85份硬粒小麦品种的检测结果基本一致。结合LOX活性测定, 我们发现 Lpx-B1位点上的2个基因是决定硬粒小麦LOX活性的最为主要因素之一。Verlotta等[ 15]筛选出低LOX活性、综合性状优异的硬粒小麦品种, 可直接用作硬粒小麦育种亲本。与LOX活性相关的功能性标记具有很好的应用前景, 尤其适合快速、准确、简便地判定小麦种质资源中LOX基因型, 并能有效预测表型, 在一定程度上加快硬粒小麦品质改良进程。
来自不同国家和地区的167份硬粒小麦品种中 Lpx-B1基因位点有 Lpx-B1.1a、 Lpx-B1.1b、 Lpx-B1.1c、 Lpx-B1.2和 Lpx-B1.3共5种变异类型, 且在同一个品种中 Lpx-B1.1a、 Lpx-B1.1b和 Lpx-B1.1c基因交替出现, 不存在2个或2个以上基因共存现象, 同样 Lpx- B1.2和 Lpx-B1.3基因也表现互补, 似为一对等位基因。根据LOX活性, 将不同等位变异分成高或低LOX活性的变异类型, 前者包括 Lpx-B1.1b和 Lpx-B1.3基因型, 后者包括 Lpx-B1.1c和 Lpx-B1.2基因型。硬粒小麦品种在2个位点上聚合高LOX活性等位变异, 则表现为高LOX活性特征, 反之则表现为低LOX活性特征; 而当2个位点分别有高和低LOX活性等位变异时, 其表现型多属于中间类型。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|