

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
低温胁迫对微胚乳超甜超高油玉米耐寒性生理生化特性的影响
[郝小琴 , 姚鹏鹤, 高峥荣, 吴子恺]
, 姚鹏鹤, 高峥荣, 吴子恺]
, 姚鹏鹤, 高峥荣, 吴子恺]
|
|


为了解新型种质微胚乳超甜超高油玉米(简称为微胚乳玉米)的耐寒性, 选用5个微胚乳玉米杂交组合(ME), 以高油115和正甜68为对照, 在发芽期和苗期低温胁迫下, 测定发芽率、发芽势、相对电导率、丙二醛含量、脯氨酸含量、可溶性糖含量、可溶性蛋白含量、叶绿素含量以及SOD、POD活性等生理生化指标, 鉴定其耐寒性, 并利用模糊隶属函数法综合评定其耐寒性。结果表明, ME1、ME2、ME3和高油115在发芽期的相对电导率上升幅度较小, 脯氨酸含量和SOD活性上升幅度较大; 而ME5、ME4和正甜68在发芽期的相对电导率上升幅度较大, 脯氨酸含量和SOD活性上升幅度较小。利用相对电导率、脯氨酸含量、SOD活性3个指标能较有效预测微胚乳玉米在发芽期的耐寒性。ME2、ME1和高油115在苗期的丙二醛含量上升幅度较小, 脯氨酸含量、SOD和POD活性上升幅度较大; 而ME5和正甜68在苗期的丙二醛含量上升幅度较大, 脯氨酸含量、SOD和POD活性上升幅度较小。利用丙二醛含量、脯氨酸含量、SOD和POD活性4个指标能较好地预测微胚乳玉米在苗期的耐寒性。以隶属函数值综合评价表明, 5个微胚乳玉米材料的耐寒性均强于正甜68。ME2在发芽期和苗期都表现较强的耐寒性, 其次为ME1和 ME3, 而ME4和ME5耐寒性均较弱。
In the germination and seedling stages of the maize, five micro-endosperm super sweet and super high oil maize (abbreviation as ME) hybrid combinations, two control materials Gaoyou 115 and Zhengtian 68 were used to study the changes of physiological and biochemical characteristics, such as germination rate, germination energy, relative conductivity, contents of MDA, proline, soluble sugar, soluble protein, chlorophyll, and SOD, POD activities under low temperature stress. At the same time, the identification of cold tolerance of the materials were conducted. Based on the multiple physiological and biochemical parameters, we employed the fuzzy membership function to evaluate the cold tolerance of the experiment materials comprehensively. The results showed that the relative conductivity had a little increase, but the proline content and SOD activity increased largely in ME1, ME2, ME3, and Gaoyou 115, as compared with ME5, ME4 and Zhengtian 68 in the germination stage. It was advised that relative conductivity, proline content and SOD activity could be used to predict cold tolerance of the materials more effectively in germination stage. In the seedling stage, compared with ME5 and Zhengtian 68, the MDA content increased a little and activities of SOD, POD and proline content increased largely in ME2, ME1, and Gaoyou 115. Therefore, we thought that MDA content, proline content, SOD and POD activities can be used to predict the cold tolerance of the experiment materials in seedling stage. Meanwhile, membership function value showed that the cold tolerance of the five ME materials was stronger than that of Zhengtian 68. Among the five ME materials, ME2 showed a strong ability to tolerate low temperature stress in germination and seedling stages, ME1 and ME3 showed moderate tolerance, and ME4 and ME5 showed weak tolerance.
玉米原产热带, 是典型的喜温C4植物, 对低温胁迫比较敏感, 容易受低温影响而严重减产。为使玉米的授粉期能避开夏季高温、干旱的不利天气, 并便于安排后茬作物而进行耕作改制, 春季适期早播成了玉米生产中的一个重要措施。而早春低温寒害是玉米生产上主要的气象灾害之一[ 1]。大部分玉米品种萌发及幼苗期不能适应早春气温骤降的这种不良气候[ 2]。虽然玉米的耐寒性较差, 但是品种间存在着较大的差异, 同一品种的不同生长发育阶段其耐寒性也不同。因此从生理生化角度比较研究不同类型玉米品种的耐寒性, 探讨各生理生化指标与耐寒性的关系, 评价不同类型玉米品种的耐寒性, 可为玉米耐寒育种、品种推广及在早春低温条件下选择适宜的品种提供依据。
玉米的耐寒性是一个复杂的生理生化过程, 仅用一两个生理生化指标难以准确地鉴定。一般主要从细胞膜透性, 膜的过氧化产物丙二醛(MDA), 胞内可溶性糖、可溶性蛋白和脯氨酸等渗透调节物质, 超氧化物歧化酶(SOD)和过氧化物酶(POD)等保护酶活性, 叶绿素等方面开展耐寒性研究。
早在20世纪70年代, 就有研究表明, 细胞膜作为低温寒害的原初部位, 其完整性和通透性与植株的耐寒性密切相关[ 3]。低温胁迫下, 细胞膜的通透性增加, 细胞内溶物外渗, 电解质渗出率增高, 致使电导率值升高, 且耐寒性弱的植株升高幅度较大[ 4]。丙二醛作为膜脂过氧化的主要产物, 其含量的高低直接反映膜脂被过氧化而受伤害的程度。低温胁迫下, 植物体内的丙二醛含量随胁迫程度的加强不断增加, 且与其耐寒性呈显著负相关[ 5, 6]。马凤鸣等[ 7]对4个普通玉米品种三叶期幼苗进行低温处理, 4个品种的MDA含量均有不同程度的增加, 且较强低温胁迫程度下抗寒性较弱品种的MDA含量增加的幅度大。脯氨酸、胞内可溶性糖、可溶性蛋白作为植物体内的渗透调节物质, 在植物的抗寒反应中起着重要的作用。有研究表明, 低温胁迫可明显增加玉米体内的脯氨酸含量, 且抗寒品种较不抗寒品种增加的幅度大[ 8]; 但也有研究表明, 植物的耐寒性与其体内脯氨酸的增加幅度呈负相关[ 9]。杨光等[ 10]从296份玉米自交系中筛选抗冷自交系的研究发现, 低温胁迫下, 各自交系的可溶性糖含量呈显著上升趋势, 并与其抗冷性呈正相关。大量研究表明, 低温胁迫下, 可溶性蛋白含量增加, 并与植物的耐寒性正相关[ 11, 12, 13]; 然而, 也有研究表明, 可溶性蛋白含量的增加与植物的耐寒性并不呈正相关[ 14, 15]。武兰芳[ 16]曾对抗冷性不同的玉米自交系三叶期幼苗的研究发现, 低温胁迫下, 各自交系的可溶性蛋白含量均下降, 而抗寒性强的下降幅度较小。保护性酶可以保护植物体免受自由基和活性氧强氧化作用的伤害, 逆境胁迫下, 植物对氧的利用能力下降, 致使体内的自由基和活性氧含量增加, 保护性酶的含量和活性也会相应适量增加。随着低温胁迫程度的加强, 植物体内的保护性酶活性逐渐升高, 且抗寒性强的升高幅度比较大[ 17, 18, 19, 20]。叶绿素在植物体光合作用的光吸收中起着核心作用, 植物光合作用的强弱直接受其含量多少的影响。低温胁迫下, 植物体内的叶绿素不仅降解加快, 而且其合成也受到抑制, 致使叶绿素含量降低。随着低温胁迫程度的加强, 植物体内的叶绿素含量逐渐下降[ 21, 22, 23, 24]。
微胚乳玉米是本课题组选育的一种新型的超甜超高油兼用型玉米种质, 与普通玉米有较大差异, 它具有胚乳少而胚相对较大的特点[ 25]。微胚乳玉米乳熟期可以作为甜玉米或水果玉米鲜食, 完熟期籽粒可以直接用于榨油, 目前尚未对其耐寒性进行研究。其耐寒性生理生化特性是否与普通玉米相同, 从微胚乳玉米中能否选育出综合耐寒性较好的材料, 尚未进行研究。
本研究选用5个微胚乳超甜超高油玉米(简称为微胚乳玉米)杂交组合为材料, 在发芽期和苗期(三叶期)低温胁迫, 通过测定发芽率、发芽势、相对电导率、丙二醛含量、脯氨酸含量、可溶性糖含量、可溶性蛋白含量、叶绿素含量、SOD活性、POD活性等一系列生理生化指标, 对微胚乳玉米发芽期及苗期的耐寒性进行比较研究, 探讨其与耐寒性的关系, 评价各微胚乳玉米材料耐寒性, 为微胚乳玉米的耐寒栽培和育种提供理论参考。
本课题组自主选育的5个微胚乳超甜超高油玉米杂交组合的编号为ME1、ME2、ME3、ME4、ME5; 以普通高油玉米高油115和超甜玉米正甜68为对照。
1.2.1 室内发芽试验 在室内常温下(日平均温度约30℃, 日最低温度约28℃, 日最高温度约32℃)进行发芽试验。设每个材料3个重复, 从每个重复
随机取30粒种子, 经0.1%升汞浸泡消毒10 min, 用蒸馏水冲洗干净, 蒸馏水浸种3 h后, 播于铺有两层湿润滤纸的发芽盒内。每天统计发芽数, 发芽结束后计算发芽率及发芽势, 并进行生理生化指标的测定。
发芽率(%)=(第7天发芽数/播种粒数)×100%
发芽势(%)=(第3天发芽数/播种粒数)×100%
于光照培养箱内进行低温发芽试验。分别设10℃、8℃、6℃ 3个温度梯度, 每天统计发芽数, 20 d后统计发芽势, 38 d后统计发芽率, 并进行生理生化指标的测定。
发芽率(%)=(第38天发芽数/播种粒数)×100%
发芽势(%)=(第20天发芽数/播种粒数)×100%
相对发芽率(势)=低温发芽率(势)/常温发芽率(势)
1.2.2 苗期低温处理 在室内常温下育苗, 长至三叶期放入培养箱内进行3℃低温胁迫。分别胁迫1 d、3 d、5 d进行生理生化指标的测定。
1.2.3 生理生化指标的测定 发芽期取发芽种子或未发芽而仍有活力的种子进行生理生化指标的测定, 苗期取去掉残余种子后的整个幼苗进行生理生化指标的测定。参照张宪政主编的《作物生理研究方法》[ 26] 测定相对电导率、丙二醛、脯氨酸、可溶性糖、可溶性蛋白、叶绿素含量及SOD、POD活性等生理生化指标。
1.2.4 数据统计与处理 利用Microsoft Excel 2007和DPS7.05软件统计分析数据, 对各项生理生化指标低温与常温对照下测定值的比值进行差异显著性检验, 用Duncan’s新复极差法进行多重比较, 采用模糊隶属函数法综合评价微胚乳玉米的耐寒性。参照许桂芳等[ 27]的方法计算模糊隶属函数。
标准差系数
权重系数
隶属函数值 D= 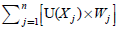 , j= 1, 2, 3,
, j= 1, 2, 3,
若指标值与耐寒性呈正相关, 则用 U( X j) = 计算, 若指标值与耐寒性呈负相关(相对电导率、丙二醛含量), 则用U( X j) = 1 -
计算。式中, X j表示第 j个指标的测定值, Xmax表示第 j个指标的最大值, Xmin表示第 j个指标的最小值。隶属函数 D值即为各供试材料在低温胁迫下耐寒性的综合评价值。
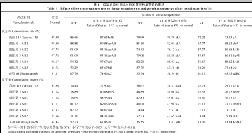
如表1所示, 随着胁迫程度的加强, 各供试材料的发芽率和发芽势均表现出下降的趋势, 但各材料的下降幅度不同。在10℃低温胁迫下, 除ME5外, 其余微胚乳玉米材料的发芽率和发芽势均高于对照高油115, 更高于对照正甜68。在8℃低温胁迫下, ME1、ME4的发芽率及ME1、ME2、ME3等微胚乳玉米材料的发芽势高于对照高油115和正甜68。6℃低温胁迫下, 高油115的发芽率和发芽势最高, 而5个微胚乳玉米材料表现不一, 其中ME1的发芽率和发芽势较高, ME2其次, 而ME5的发芽率最低, ME4的发芽势最低。
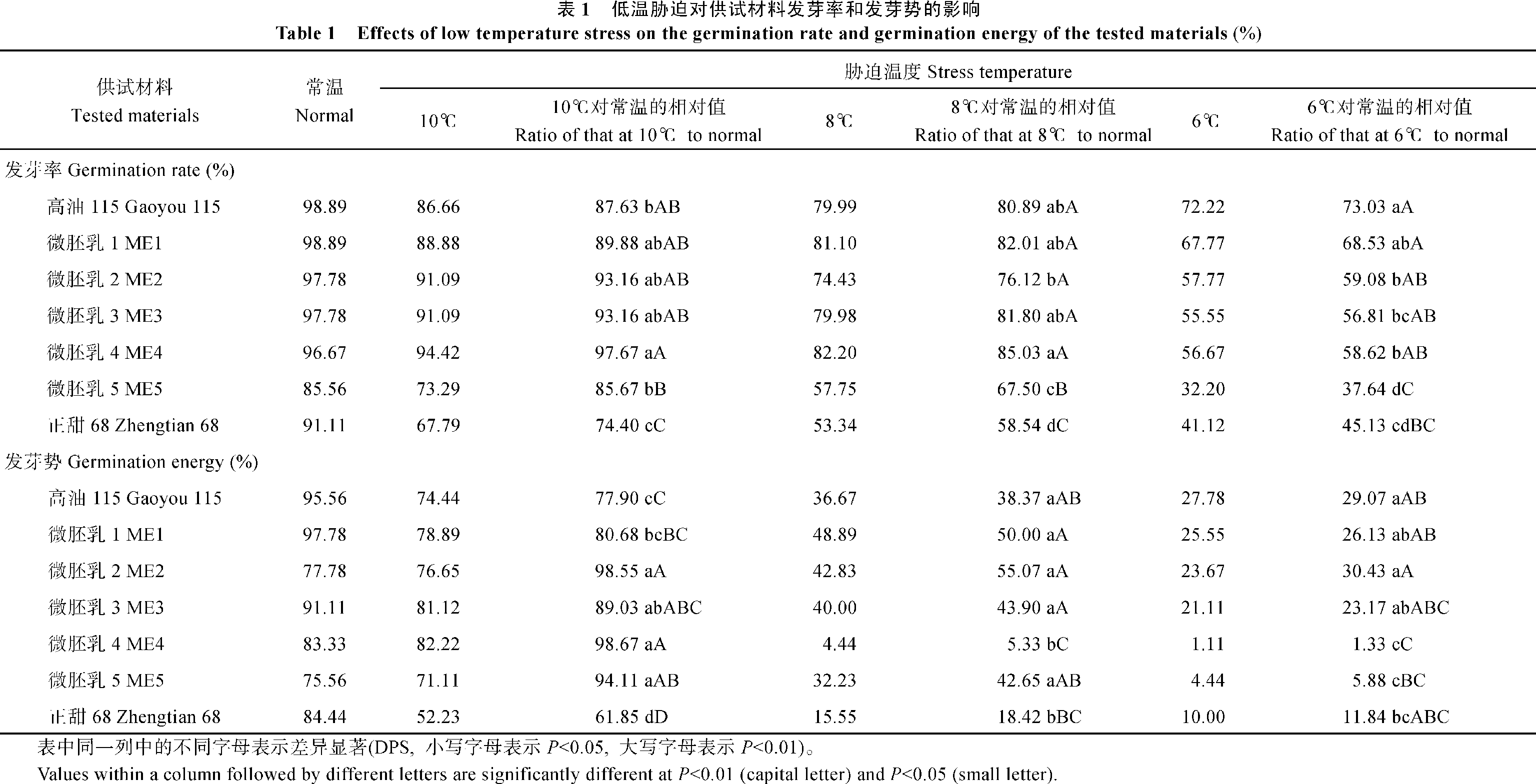 | 表1 Table 1 |
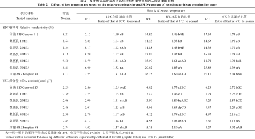
2.2.1 对相对电导率和丙二醛含量的影响 如表2所示, 随着胁迫程度的加强, 各供试材料的相对电导率和丙二醛含量均表现出逐渐上升的趋势。ME1、ME2和ME3的相对电导率上升幅度较小, 而ME5上升幅度最大。ME1、ME3、ME4的丙二醛含量上升幅度较小, 而ME5上升幅度最大。
2.2.2 对脯氨酸含量、可溶性糖含量、可溶性蛋白含量的影响 如表3所示, 随着胁迫程度的加强, 各供试材料的脯氨酸含量均表现出逐渐上升的趋势, 但上升幅度不同。对照高油115上升幅度最大, ME1次之, 再次是ME2和ME3, 而ME5上升幅度最小。除ME4外, 各供试材料的可溶性糖含量均随着胁迫的加强呈现先升后降趋势; 10℃低温下达峰值, 8℃低温下呈下降趋势, 其中ME1的降幅最大, 6℃低温下, ME4略有回升, 其余6个材料呈继续下降趋势。各供试材料的可溶性蛋白含量均随着胁迫的加强表现出先上升后下降的趋势, 但其变化幅度不同。各材料在10℃低温下均达峰值; 8℃低温下均表现出下降趋势; 6℃低温下继续下降, ME2的降幅最小, 其次是ME3, 但二者仍高于常温对照, 正甜68的降幅最大, 其可溶性蛋白含量仅为常温对照的1/2。
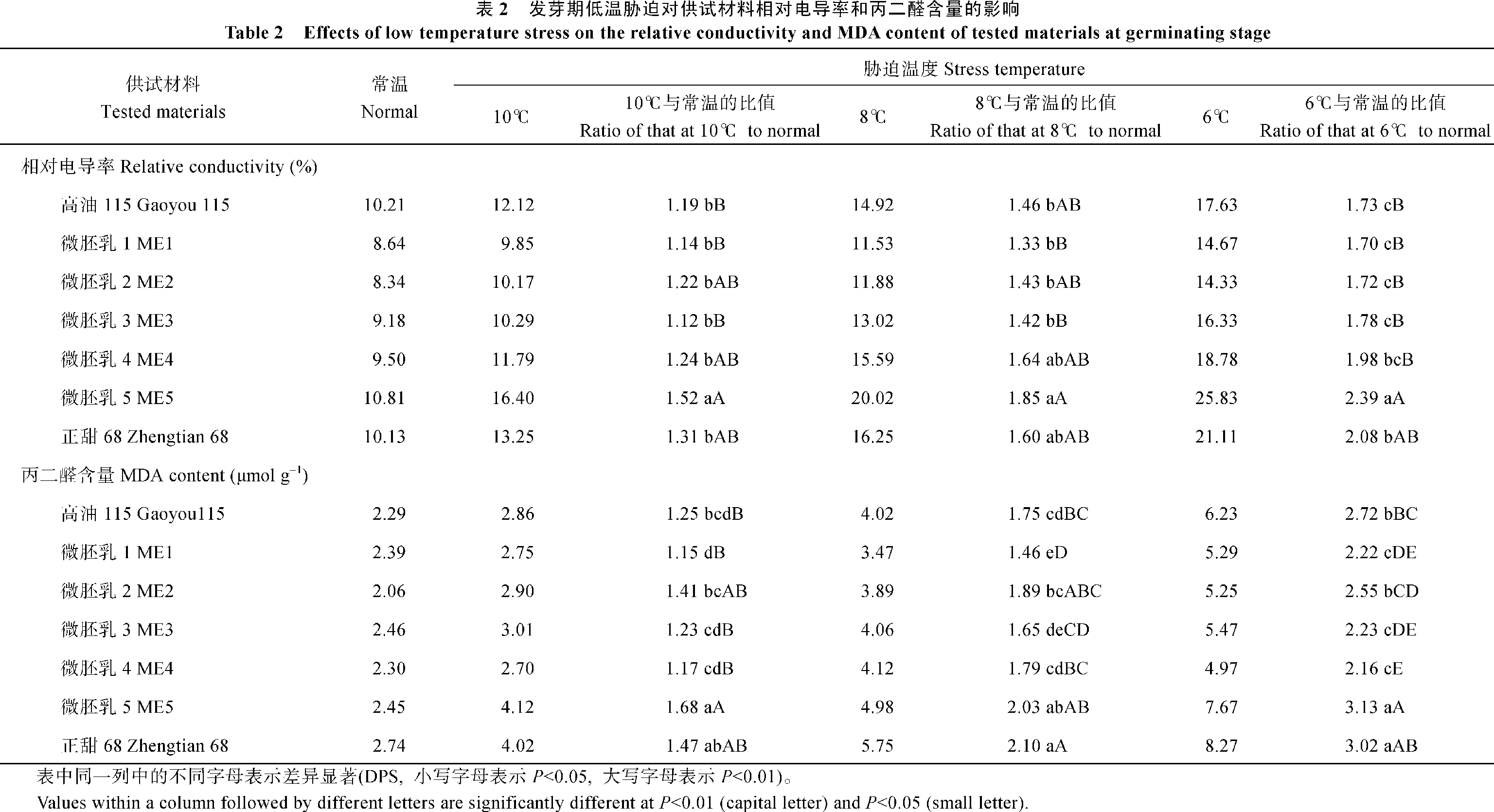 | 表2 Table 2 |
 | 表3 Table 3 |
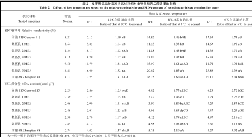
2.2.3 对SOD和POD活性的影响 如表4所示, 随着胁迫程度的加强, 各供试材料的SOD活性均表现出先上升后下降的趋势, 但变化幅度不同。各供试材料在10℃低温下均达峰值; 8℃低温下均呈下降趋势; 6℃低温下继续下降, 其中ME2的下降幅度最小, 其次是ME1, 正甜68的下降幅度最大。随着胁迫程度的加强, 各供试材料POD活性的变化趋势各不相同, 但总体上表现为先下降后上升再下降的趋势, 10℃低温下均下降; 8℃低温下, 除ME3和正甜68外均有小幅上升; 6℃低温下, ME3继续升高, 其余6个供试材料又有小幅下降。
| 表4 发芽期低温胁迫对供试材料SOD和POD活性的影响 Table 4 Effects of low temperature stress on SOD and POD activities of tested materials at germinating stage |
2.3.1 对相对电导率和丙二醛含量的影响 随着胁迫的加强, 各供试材料的相对电导率和丙二醛含量均表现出逐渐上升的趋势, 但上升幅度不同。随低温胁迫时间的延长, 相对电导率上升幅度以ME2、ME3和ME1较小, ME4、ME5较大, 正甜68最大。ME2的丙二醛含量上升幅度最小, 其次是对
照高油115和ME1, 但三者差异不显著; 对照正甜68的丙二醛含量上升幅度最大(表5)。
| 表5 苗期低温胁迫对供试材料相对电导率和丙二醛含量的影响 Table 5 Effects of low temperature stress on the relative conductivity and MDA content of tested materials at seedling stage |
2.3.2 对脯氨酸含量、可溶性糖含量、可溶性蛋白含量的影响 如表6所示, 随着胁迫的加强, 各供试材料的脯氨酸含量均表现逐渐上升趋势。ME2、对照高油115、ME1升幅较大, 对照正甜68升幅最小。
随着胁迫程度的加强, 可溶性糖含量大体呈先下降后上升的趋势, 但各材料的变化幅度不同。低温胁迫1 d时均呈下降趋势; 低温胁迫3 d时均呈缓慢上升趋势; 低温胁迫5 d时, ME5和正甜68呈持续下降趋势, 其他供试材料均呈继续上升趋势, 高油115、ME2上升幅度较大。
随着胁迫程度的加强, 可溶性蛋白含量均呈先上升后下降的趋势。低温胁迫1 d时均达峰值; 低温胁迫3 d时均呈下降趋势; 低温胁迫5 d时, 除ME3外, 其余供试材料均持续下降, 但仍高于常温下的可溶性蛋白含量, 其中ME1的下降幅度最小, 为常温对照的3.61倍, ME5的降幅最大, 为常温对照的2.55倍。
2.3.3 对SOD、POD活性和叶绿素含量的影响
如表7所示, 随着胁迫程度的加强, ME3、ME4、对照正甜68的SOD活性呈先升后降趋势, 其余供试材料均呈先升后降再升的趋势, ME2的上升幅度最大。各供试材料的POD活性均呈先升后降再升的趋势, 但变化幅度不同。低温胁迫1 d, 各供试材料的POD活性均呈上升趋势, 除ME4外, 其余供试材料的POD活性均达峰值; 低温胁迫3 d, 各供试材料POD活性均呈下降趋势; 低温胁迫5 d, 各供试材料POD活性均又呈上升趋势, 其中ME2的升幅最大。
| 表6 苗期低温胁迫对供试材料脯氨酸含量、可溶性糖含量、可溶性蛋白含量的影响 Table 6 Effects of low temperature stress on proline content, soluble sugar content and soluble protein content of tested materials at seedling stage |
随着胁迫程度的加强, 各供试材料的叶绿素含量大体呈逐渐下降的趋势, 但降幅不同。ME2降幅最小, 高油115其次; 正甜68降幅最大, 与其他供试材料差异均显著或极显著。
| 表7 苗期低温胁迫对供试材料SOD、POD酶活性和叶绿素含量的影响 Table 7 Effects of low temperature stress on SOD, POD activities and chlorophyll content of tested materials at seedling stage |
表8表明, 相对电导率、丙二醛含量与其他指标多呈负相关, 含量越高, 耐寒性越差。脯氨酸、可溶性糖、可溶性蛋白含量、SOD、POD活性、叶绿素含量对耐寒性起正向作用, 含量越高, 耐寒性越好。在发芽期, 相对电导率与发芽率、发芽势、脯氨酸含量、SOD活性呈显著或极显著负相关; 脯氨酸含量与发芽率、发芽势、SOD活性呈显著或极显著正相关; SOD活性与发芽势呈显著正相关。而丙二醛、可溶性糖、可溶性蛋白含量、POD活性与其他指标间的相关性多数未达显著水平。苗期除可溶性蛋白含量外, 其他指标之间的相关性多数呈显著或极显著水平, 且丙二醛、脯氨酸含量、POD活性间的相关相对比较密切。
可见, 相对电导率、脯氨酸含量、SOD活性可作为微胚乳玉米发芽期的耐寒性鉴定指标。苗期若采用可溶性蛋白含量进行耐寒性鉴定, 必须结合其他指标; 丙二醛、脯氨酸含量和POD活性可更好地作为微胚乳玉米苗期的耐寒性鉴定指标。
为避免单一指标的局限性与片面性, 在进行植物耐寒性鉴定时, 应结合多个指标综合评定, 才能得出相对准确的结论, 正确反映植物的耐寒性实质。
采用模糊隶属函数法, 利用发芽率、发芽势、相对电导率、丙二醛、脯氨酸、可溶性糖、可溶性蛋白含量及SOD、POD活性共9个指标为依据, 对微胚乳玉米发芽期的耐寒性进行综合评价。综合评价值越大, 表明其耐寒能力越强。由表9可以看出, 7个供试材料间的综合评价值差异较大, 介于0.165~0.801之间。发芽期的综合评价值表现为ME2>ME3>高油115>ME1>ME4>ME5>正甜68。ME2的综合评价值高达0.801, 耐寒性最强, ME3次之; 对照高油115和ME1的综合评价值大于0.6, 耐寒性也较强, 而ME4、ME5的耐寒性较弱, 对照正甜68的综合评价值仅为0.165, 耐寒性最差。
根据发芽期各指标的隶属函数值与综合评价值的排序, 利用相对电导率、脯氨酸含量、SOD活性这3个指标能较有效预测微胚乳玉米在发芽期的耐寒性; 而若利用丙二醛含量、可溶性糖含量、可溶性蛋白含量和POD活性这几个指标预测微胚乳玉米在发芽期的耐寒性, 预测效果比较差。
采用模糊隶属函数法, 以相对电导率、丙二醛、脯氨酸、可溶性糖、可溶性蛋白、叶绿素含量及SOD、POD活性共8个指标为依据, 对微胚乳玉米三叶幼苗期的耐寒性进行综合评价, 如表9所示。7个供试材料的综合评价值介于0.003~0.931之间。苗期的综合评价值表现为ME2>高油115>ME1>ME3>ME4>ME5>正甜68。ME2的综合评价值高达0.931, 耐寒性最强, 对照高油115次之; ME1、ME3的综合评价值大于0.6, 耐寒性也较强, ME4、ME5的耐寒性较差; 对照正甜68的综合评价值仅为0.003, 耐寒性最差。
根据苗期各生理生化指标的隶属函数值与综合评价值的排序, 利用丙二醛、脯氨酸含量、SOD和POD活性这4个指标能较有效预测微胚乳玉米苗期耐寒性; 而若利用相对电导率、可溶性糖、可溶性蛋白和叶绿素含量这几个指标预测微胚乳玉米苗期耐寒性, 则效果略差。
 | 表8 Table 8 |
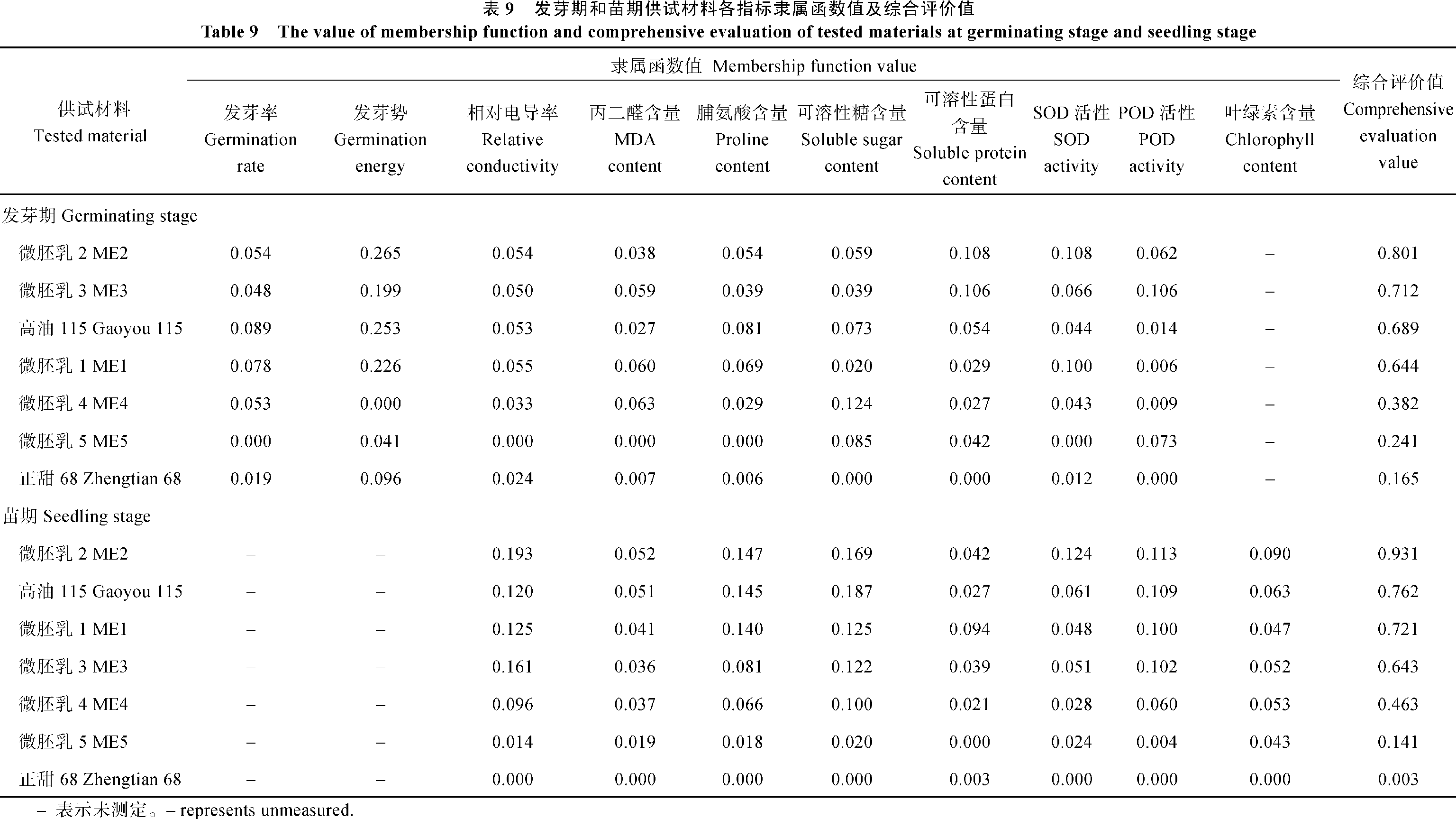 | 表9 Table 9 |
植物的耐寒性是一系列复杂的生理生化过程综合作用的表现。本试验测定微胚乳玉米发芽期和苗期低温胁迫下多个生理生化指标, 直接分析各指标与耐寒性的关系, 并结合各指标间的简单相关分析和隶属函数值综合评价。结果表明, 利用相对电导率、脯氨酸含量、SOD活性3个生理生化指标能较有效预测微胚乳玉米在发芽期的耐寒性; 利用丙二醛含量、脯氨酸含量、SOD活性和POD活性4个指标能较好预测微胚乳玉米在苗期的耐寒性。
低温胁迫下, 相对电导率值的高低直接表明植物细胞膜受伤害的程度。丙二醛是细胞膜脂过氧化的主要产物, 其含量直接反映膜脂被过氧化而受损害的程度。因此, 相对电导率和丙二醛是衡量低温伤害比较直接的鉴定指标。耐寒能力强的植物经低温胁迫后细胞膜受伤害的程度较小, 相对电导率值和丙二醛含量上升的幅度小。本试验结果表明, 各供试材料在发芽期低温胁迫, 相对电导率比较敏感, 而在苗期低温胁迫, 丙二醛含量相对较敏感。在发芽期和苗期, 各供试材料的相对电导率和丙二醛含量均随着胁迫程度的加强而不断升高, 且ME1和ME2升高的幅度较小, 耐寒性较强, ME5升高的幅度较大, 耐寒性较弱。这与马凤鸣等[ 7]以普通玉米为材料研究的结果一致。
脯氨酸作为植物体内的主要渗透调节物质, 也是鉴定植物耐寒性的一个重要指标。植物遭遇低温寒害时, 由于脯氨酸保护了酶的空间结构, 为植物体内的生化反应提供了足够的生理生化活性物质及自由水, 对细胞起了一定的保护作用[ 11, 28]。脯氨酸含量的增加是植物对寒冷适应的重要生理反应, 耐寒性强的增加幅度较大。本试验表明, 随着低温胁迫的加强, 各供试材料的脯氨酸含量逐渐增加。且耐寒性强的ME1和ME2上升幅度较大, 耐寒性弱的ME5上升幅度较小。这与高素华[ 8]报道的以普通玉米为材料的结果一致。
保护性酶可以保护植物体免受自由基和活性氧强氧化作用的伤害。大量研究表明, 随着低温胁迫的加强, 植物体内的保护性酶活性逐渐升高, 且抗寒性强的升高幅度比较大[ 17, 18, 19, 20]。本试验表明, 随着胁迫的加强, 各供试材料的SOD活性均表现出先升后降的趋势, 且耐寒性强的ME1、ME2和ME3上升幅度较大, 耐寒性弱的ME4和ME5上升幅度较小。但POD活性的升降趋势不尽一致。可能是在一定的低温胁迫下, 各供试材料植物体内保护酶活性会适量升高, 以维持自由基和活性氧产生与清除之间的平衡, 但随着低温胁迫的持续加强, 体内活性氧和自由基的产生速度加快, 打破了产生与清除的平衡, 使保护酶活性又出现一定程度的下降。
本试验中5个微胚乳玉米杂交组合在发芽期和苗期的耐寒性都存在较大差异, 根据模糊隶属函数法的综合评价表明, ME2是耐寒性最好的组合, 说明从微胚乳玉米中选出发芽期和苗期耐寒性均较好的杂交组合是完全可能的。由于微胚乳玉米是一种超甜和超高油兼用的新型玉米种质, 具较好推广前景, 所以本试验选用在我国较大面积推广的普通高油玉米品种高油115和在甜玉米主产区较大面积推广的正甜68超甜玉米品种作为对照。微胚乳玉米几乎不含胚乳, 而正甜68含部分胚乳, 高油115含胚乳较多, 属于含胚乳少、中、多的3种不同类型玉米种质。虽然微胚乳玉米的胚乳含量极少, 但在遭遇低温胁迫时, 几个重要耐寒性指标均优于对照正甜68, 有些甚至优于对照高油115, 其机理值得进一步研究。
本研究中, 5个微胚乳玉米杂交组合在发芽期和苗期的耐寒性均优于超甜玉米正甜68, 说明微胚乳玉米可作为超甜玉米或水果玉米收获青苞在生产上应用, 春季可以适期早播, 在南方可一年两到三熟, 提高经济效益。ME2在发芽期和苗期的耐寒性均优于高油115, ME1是已审定的品种, 其耐寒性也比较好。在2010年黑龙江遭遇严重春寒情况下, 本课题组曾将ME1比当地普通玉米和大豆都提早播种, 播后遇到表层土壤结冰的天气, 但种子仍未遭冻害, 能正常出苗。说明这些耐寒性较好的微胚乳玉米杂交组合可作为油用型玉米在生产上推广, 在南北方都可适期早播。
微胚乳玉米发芽期和苗期的耐寒性鉴定需结合多个生理生化指标综合评价才更合理。ME1、ME2、ME3和高油115在发芽期的相对电导率上升幅度较小, 脯氨酸含量和SOD活性上升幅度较大; 而ME5、ME4和正甜68在发芽期的相对电导率上升幅度较大, 脯氨酸含量和SOD活性上升幅度较小。利用相对电导率、脯氨酸含量、SOD活性3个指标能较有效预测微胚乳玉米在发芽期的耐寒性。ME2、ME1和高油115在苗期的丙二醛含量上升幅度较小, 脯氨酸含量、SOD和POD活性上升幅度较大; 而ME5和正甜68在苗期的丙二醛含量上升幅度较大, 脯氨酸含量、SOD和POD活性上升幅度较小。利用丙二醛含量、脯氨酸含量、SOD和POD活性4个指标能较好预测微胚乳玉米在苗期的耐寒性。隶属函数值综合评价表明, 5个微胚乳玉米材料的耐寒性相对于对照高油115有强有弱, 但其耐寒性均强于对照正甜68。ME2在发芽期和苗期都表现出了较强的耐寒性, 其次为ME1和ME3, 而ME4和ME5耐寒性均较弱。表明从微胚乳玉米中可选育出发芽期和苗期综合耐寒性较好的材料。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|