{kind=link}
{kind=link}
{kind=link}
[周清元, 崔翠, 阴涛, 陈东亮, 张正圣, 李加纳*  ]
]
]
|
|


角果是油菜产量构成要素中重要的组成部分。本文以长角果品种中双11和短角果材料10D130为亲本配制杂交组合, 采用主基因+多基因混合遗传模型分析方法对该组合6世代遗传群体(P1、P2、F1、BCP1、BCP2和F2)的果身长、角果长和果喙长进行遗传分析。结果表明, 该组合的3个角果性状均呈连续分布, 其中, 果身长最适遗传模型为E-0 (2对加性-显性-上位性主基因+加性-显性-上位性多基因模型), 2对主基因加性效应值分别是1.75和-0.06, 显性效应值分别是-0.59和-0.86, 主基因遗传率在BCP1、BCP2和F2中分别是51.10%、74.23%和66.93%, 多基因遗传率分别为29.16%、17.11%和23.96%。角果长的最适遗传模型为E-1 (2对加性-显性-上位性主基因+加性-显性多基因模型), 其中, 第1对主基因加性效应为0.34, 显性效应为-0.81, 第2对主基因加性效应为0.34, 显性效应为-0.47, 主基因遗传率在BCP1、BCP2和F2中分别是47.63%、68.51%和79.45%, 多基因遗传率分别为29.40%、20.89%和12.47%。果喙长的最适遗传模型为E-3模型 (2对加性主基因+加-显多基因遗传模型), 2对主基因加性效应值分别是0.2和-0.2, 主基因遗传率在BCP1、BCP2和F2中分别是33.71%、72.75%和52.25%, 多基因遗传率分别为40.08%、5.37%和27.60%。
Silique is one of major components for rapeseed yield. Inheritance of silique body length (SBL), valid silique length (VSL) and beak length (BL) in a cross of variety Zhongshuang 11 with long silique (P1) and line 10D130 with short silique (P2) was investigated by the mixed major gene plus polygene inheritance model. The results showed that SBL, VSL and BL in the populations of F2, BCP1, and BCP2 were controlled by the major gene and polygenes. The SBL were dominated by two major genes with additive-dominance-epistasis effects plus polygenes with additive-dominance-epistasis effects (E-0 model). The heritability values of the major genes of SBL in BCP1, BCP2, and F2 were estimated as 51.10%, 74.23%, and 66.93%, respectively, and the heritability values of the polygene were 29.16%, 17.11%, and 23.96%, respectively. The additive effects of two major genes of SBL were 1.75 and -0.06, and the dominant effects of two major genes were -0.59 and -0.86, respectively. The valid silique length was controlled by two major genes with additive-dominance-epistasis effects plus polygenes with additive-dominance effects (E-1 model). Heritability values of the major genes for SL in BCP1, BCP2, and F2generations were estimated as 47.63%, 68.51%, and 79.45%, respectively, and the heritability values of the polygene were 29.40%, 20.89%, and 12.47%, respectively. The additive effects of two major genes were equal (0.34) to that of the cross, but the dominant effects of the two major genes were -0.81 and -0.47, respectively. The beak length was dominated by two major gene with additive effects plus polygenes with additive-dominance effects (E-3 model). Heritability values of the major genes of BL in the cross were 33.71%, 72.75%, and 52.25%, respectively, and the heritability values of the polygene were 40.08%, 5.37%, and 27.60%, respectively. The additive effects of two major genes were 0.20 and -0.20, respectively.
我国油菜常年种植面积和总产均居世界首位。随着人口的增长和生物柴油概念的提出, 油菜需求日益增长, 供求矛盾凸显, 因此, 提高油菜单产水平成为重要的育种目标, 也是解决该矛盾的有效方式之一。角果是油菜产量构成因素中重要的组成部分, 角果及角果相关性状对油菜产量具有直接或者间接的作用。油菜角果作为油菜的结实器官, 不仅是重要的贮藏器官, 也是重要的光合器官, 具有“库”和“源”的双重作用[ 1, 2]。在油菜生长初期, 油菜的光合作用主要通过叶片和茎杆来完成, 但是当油菜进入到盛花期和角果快速生长时期, 角果及其内部籽粒的生长所需有机物大多都需要角果光合作用来提供, 叶片的作用减少[ 3, 4]。因此, 角果的生长发育严重影响油菜的产量和品质[ 5]。角果长度是影响油菜产量的重要因素[ 6], 与每果粒数、千粒重显著相关[ 7, 8]。培养长角果可以提高油菜的产量潜力[ 7], 角果长度主要受到核基因控制, 且不同的研究材料和杂交组合在不同的研究方法下具有不同的遗传模式[ 8, 9, 10, 11, 12]。
育种目标性状, 如产量、品质、生育期、耐逆性等大部分是数量性状。数量性状的遗传体系比较复杂, 可以由个别或者少数主基因组成、若干微效基因组成、也可以是个别或者少数主基因与若干微效基因共同组成。王建康等[ 13]提出了分析主基因+多基因混合遗传模型的原则和步骤, 建立了多世代群体的联合分析方法, 近年已广泛应用到不同作物的不同性状上, 如水稻和小麦的抗病性[ 14, 15, 16, 17]、大豆的根系性状[ 18]、棉花的耐低温[ 19]、花生产量[ 20]、黄瓜种子休眠[ 21]、油菜的含油量和芥酸含量[ 22, 23, 24]、油菜株高和抗倒性[ 25, 26]、油菜角果种子粒数和千粒重[ 27, 28]、油菜花色[ 29]等性状, 并取得了很好的研究进展。本文以6个世代研究群体(P1、P2、F1、F2、BCP1和BCP2), 通过主基因+多基因混合遗传模型方法研究角果长度相关的3个性状, 以期揭示角果长度的遗传机制、评估单个主基因不同的遗传效应和各分离世代的遗传率, 为进一步研究和利用奠定基础。
亲本材料分别为中双11 (P1)和10D130 (P2), 中双11是中国农业科学院油料所选育的常规优质油菜品种, 主要遗传来源为(中双96/2F10)//26102, 角果长度在9~10 cm之间; 10D130是由西南大学油菜工程技术中心从芥菜型油菜和羽衣甘蓝种间杂种后代中选育出来的高油黄籽半矮杆短角果稳定的高世代自交系(S6)[ 30], 角果长度在5~6 cm之间。从2009年到2011年, 我们连续3年将2个亲本材料杂交, 并在次年观察其杂种F1代角果长度, 结果均表明, 2个亲本间角果长度(3个性状)差异显著( P<0.05), 且F1介于2个亲本之间, 与2012年研究结果相似。
2009年秋, 在西南大学油菜工程技术中心歇马实验基地(29.45°N, 106.22°E)种植亲本, 2010年春配制并获得F1(P1×P2)种子。2011年春对F1套袋自交得到F2, 并配制获得BCP1(F1×P1)和BCP2(F1×P2)。2011年, 收集6个世代(P1、P2、F1、F2、BCP1和BCP2)种子。
2011年9月, 将6个世代群体种子同期播种在西南大学油菜工程技术中心歇马实验基地, 11月1日移栽。行距 40 cm、株距21 cm, 每窝单株。试验地前茬为空闲地, 黏壤土, 肥力中等; 2011年10月26日基施375 kg hm-2复合肥(N∶P∶K=1∶1∶1), 27日由拖拉机旋耕, 深度30~40 cm。2011年12月3日追肥(每公顷施52 500 kg清水+75 kg尿素), 12月13日中耕培土1次。其他措施按常规栽培管理。其中, P1、P2和F1每个群体种植3~5行(每行10株), F2群体共种植38行, BCP1群体种植13行, BCP2群体种植19行。2012年5月8号按单株收获各群体。自然风干后, 在每个单株主花序中部收获发育完全的10个角果, 分别测定其果身长(包含果喙长度)、有效角果长(不包含果喙长度)和果喙长度。计算10个角果的平均长度作为单株角果长度, 用于遗传分析。
根据盖钧镒等[ 13]和章元明等[ 31]提出的植物数量性状主基因+多基因混合遗传多世代联合分析方法, 对油菜角果长性状进行联合分析, 采用极大似然法和IECM算法估计各世代、各成分分布的参数, 通过AIC值选择最佳模型[ 32], 并进行一组适合性检验, 包括均匀性 U12、 U22和 U32检验, Kolmo-Smimov检验( n W2)和Kolmo-Gorov检验( D n), 根据检验结果选择最优遗传模型。最后采用最小二乘法依据最优模型的各成分分布参数估计各基因效应值、方差等遗传参数[ 13]。植物数量性状主基因+多基因遗传体系分析软件由南京农业大学国家大豆改良中心提供(http://jpkc. njau.edu.cn/swtj/show.asp?classid=35&classtype=26)。用Mi-crosoft Excel软件估算平均数、遗传参数等, DPS7.05软件进行方差分析和多重比较。
比较亲本和F1群体3个角果长性状(表1)表明, 中双11的角果果身长、有效角果长和果喙长分别为9.87、8.32和1.55, 而10D130 3个性状分别为5.55、4.37和1.18, 2个亲本3个性状间差异均达到显著水平( P<0.05); 杂种F1的这3个性状分别为8.97、7.35和1.61, 更接近于中双11, 与10D130差异达到显著水平( P<0.05)。表2表明, 3个分离世代群体(F2、BCP1和BCP2)的极差和变异系数均大于3个不分离世代群体(P1、P2和F1), 说明分离世代群体离散程度高, 具有较高的遗传多态性, 为进一步遗传分析提供较好的研究基础。
| 表1 中双11、10D130及其F1角果长(2012) Table 1 Statistics of traits related to silique among P1, P2, and F1 generations in 2012 (mean± SD, cm) |
| 表2 中双11×10D130各世代角果长极差和变异系数 Table 2 Variation coefficients of rapeseed traits related to silique in six generations of ZS11×10D130 |
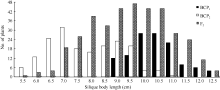
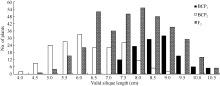
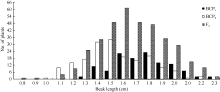
将3个世代群体(BCP1、BCP2和F2)角果的果身长、有效角果长和果喙长分组, 统计其分布频次(图1、图2和图3)表明, 该组合角果长度3个性状具有广泛的遗传变异, 且均呈连续性分布, 不易分组, 具有典型的数量性状遗传特征。另外, 部分分离世代群体具有明显多峰现象, 说明该组合3个角果长度性状属于受到主基因遗传效应影响的数量性状。
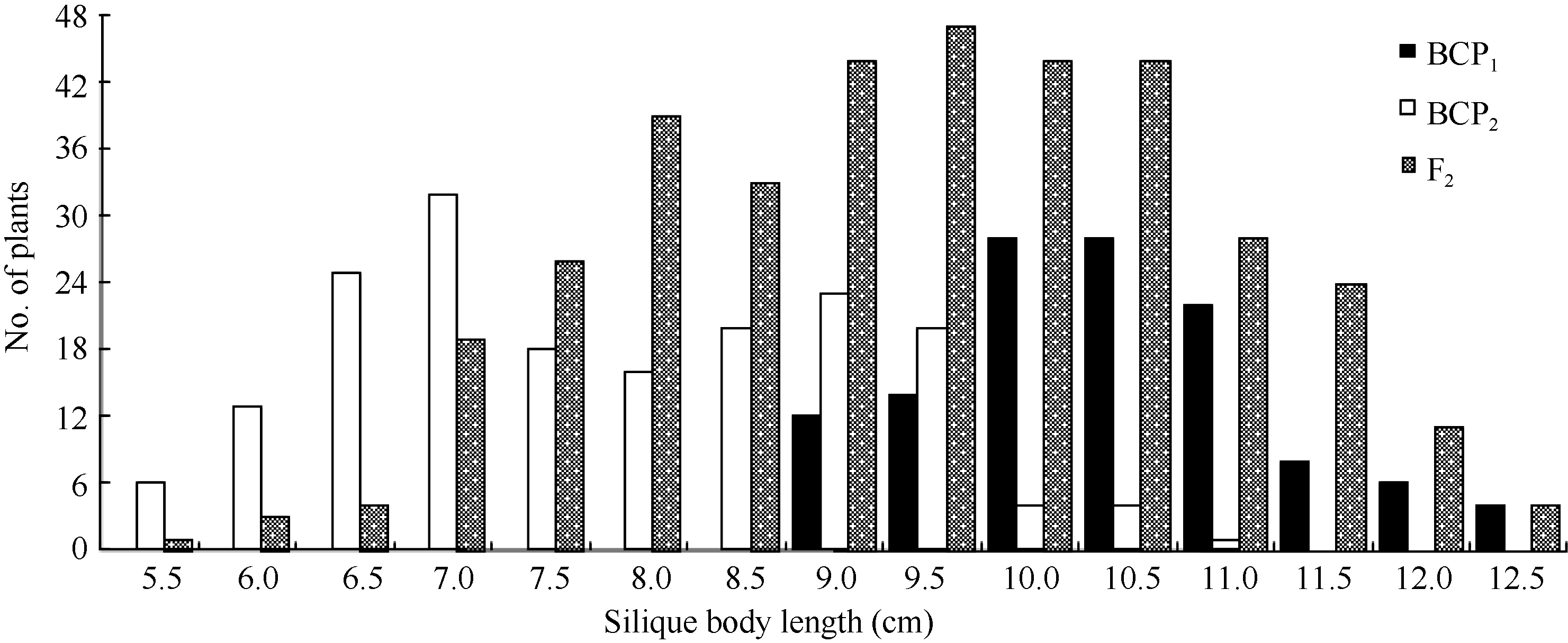 | 图1 中双11×10D130杂交组合3个分离世代群体果身长的次数分布Fig. 1 Frequency distribution of silique body length in BCP1, BCP2, and F2 populations from the cross of ZS11×10D130 |
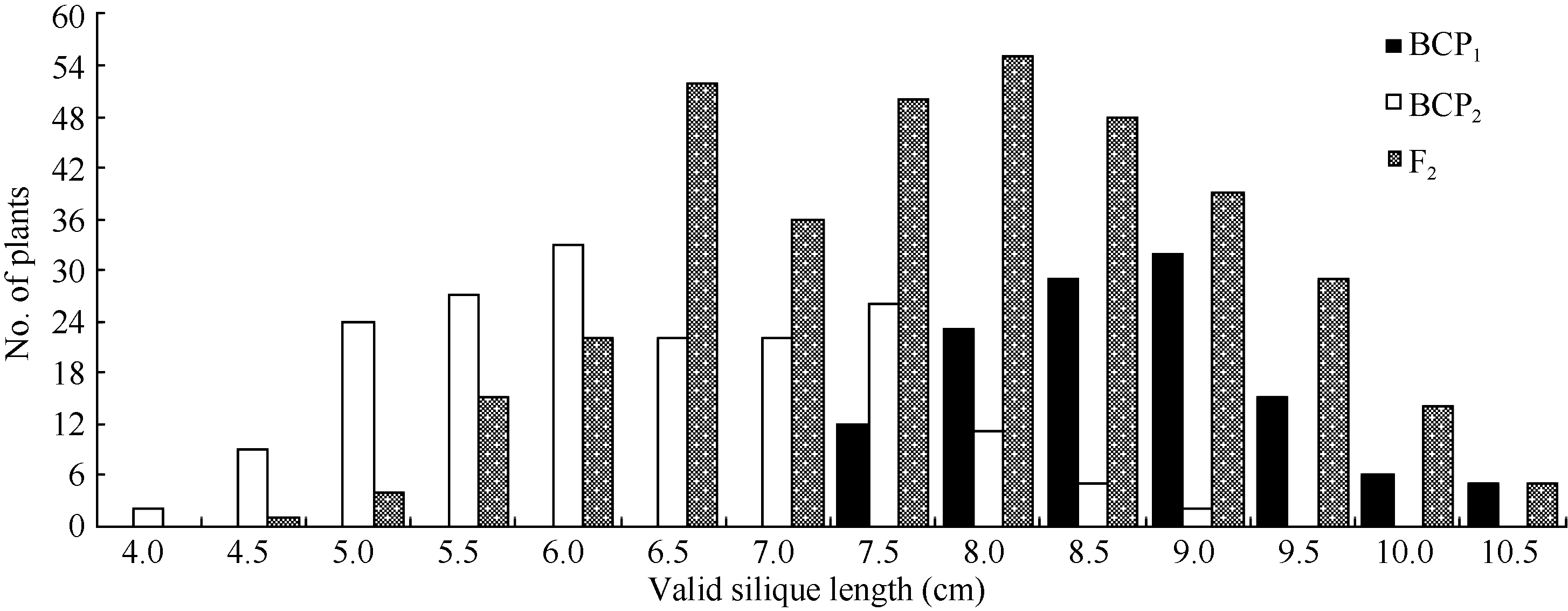 | 图2 中双11×10D130杂交组合3个分离世代群体角果长的次数分布Fig. 2 Frequency distribution of valid silique length in BCP1, BCP2, and F2 populations from the cross of ZS11×10D130 |
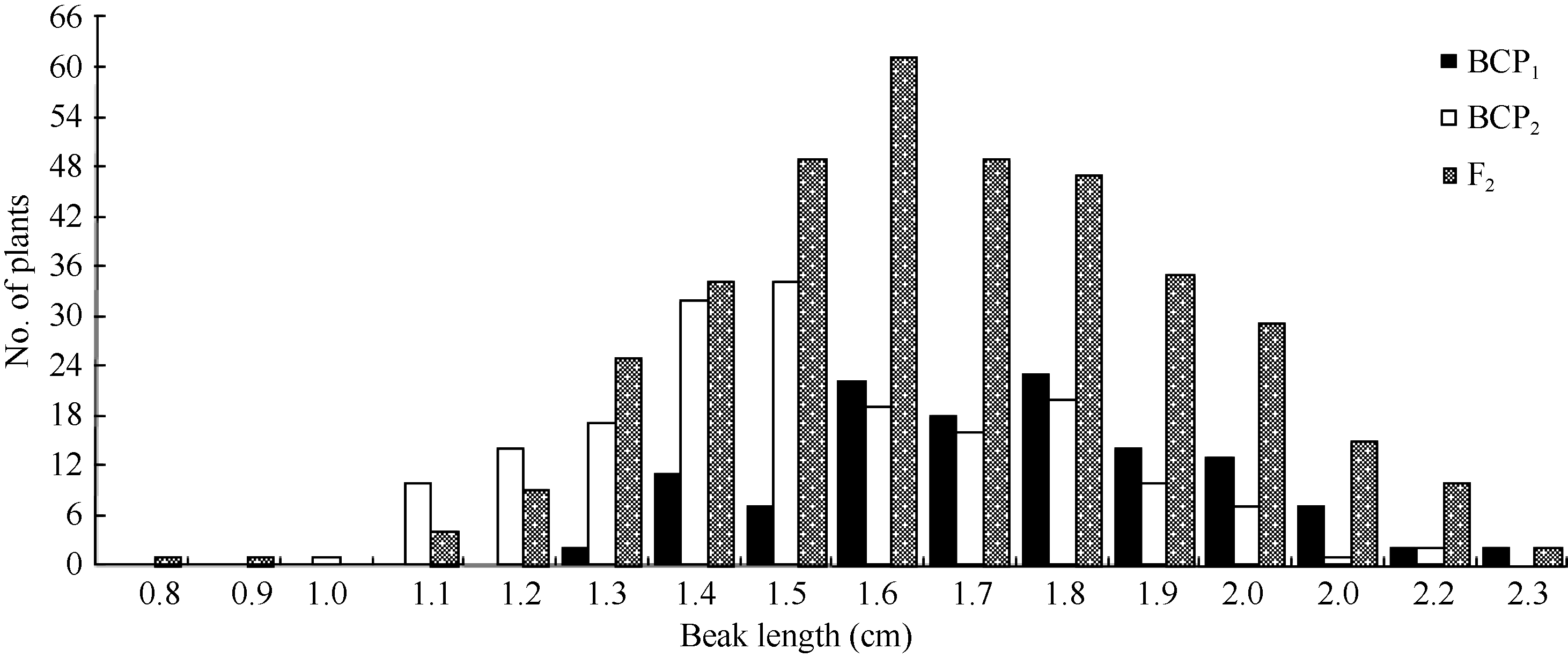 | 图3 中双11×10D130杂交组合3个分离世代群体果喙长的次数分布Fig. 3 Frequency distribution of beak length in BCP1, BCP2, and F2 populations from the cross of ZS11×10D130 |
估算获得1对主基因(A类)、2对主基因(B类)、多基因(C类)、1对主基因+多基因(D类)、2对主基因+多基因(E类)共5类24种遗传模型的极大似然函数值和AIC值[ 13], 每个性状获得23种遗传模型的AIC值(表3)。根据遗传模型的选取原则, 即AIC最小准则[ 32], 选取AIC值最小及与最小AIC值比较接近的一组遗传模型作为备选模型。结果3个角果性状的AIC值主要以E类模型为较小, 其中果身长和角果长备选模型为E-0和E-1, 果喙长备选模型为E-1和E-3。
| 表3 中双11×10D130组合后代各遗传模型的AIC值 Table 3 AIC values of the genetic models from the cross of ZS11×10D130 |
| 表4 中双11×10D130组合后代备选遗传模型的适合性检验 Table 4 Test of goodness-of-fit in the selected genetic models from the cross of 10D130×ZS11 |
| 表5 10D130×中双11杂交组合角果长的一阶遗传参数估计值 Table 5 First order genetic parameters estimated for the fitting models from the cross of 10D130×ZS11 |
| 表6 10D130×中双11杂交组合角果长的二阶遗传参数估计值 Table 6 Second order genetic parameters estimated for the fitting models from the cross of ZS11×10D130 |
对备选模型进行一组( U12、 U22、 U32、 n W
根据选择的最优遗传模型估计3个性状最优遗传模型一阶、二阶遗传参数[ 13](表5和表6)。控制角果果身长的2对主基因加性效应值差别较大, 分别为1.75和-0.06, 第1对基因加性效应( d a)作用大于第2对基因( d b), 长角果亲本中双11加性效应大于短角果亲本10D130; 2对基因的显性效应值( h a和 h b)分别为-0.59和-0.86, 均为负值, 表明10D130显性效应作用大于中双11。2对基因的加性效应间互作( i)为-0.16, 显性效应间互作( l)为1.76, 2对基因加性和显性效应之间的互作( j ab和 j ba)分别为-0.81和0.20, 表明2对基因之间存在着明显互作, 对该组合果身长影响较大。2对基因的显性度( h a/ d a和 h b/ d b )分别为-0.34和13.77, 说明第1对主基因部分显性, 第2对基因超显性。BCP1、BCP2和F2 分离世代群体遗传率分别为80.26%、91.34%和90.89%, 其中主基因遗传率分别为51.10%、74.23%和66.93%, 多基因遗传率分别为29.16%、17.11%和23.96%, 主基因遗传率占总遗传率的比值分别为0.64, 0.81和0.74, 结果表明果身长主要受到2对主基因影响。3个分离世代群体环境因素引起的变异分别占19.74%、8.66%和9.11%。
控制角果长的2对主基因的加性效应( d a和 d b)大小相当, 都为0.34, 2对主基因的显性效应( h a和 h b)分别为-0.81和-0.476。2对主基因的加性互作效应( i)为-1.5, 显性互作效应( l)为1.75, 2对主基因加性和显性效应之间的互作( j ab和 j ba)分别为0.86和-0.17, 说明2对基因之间存在着明显互作。2对基因的显性度 h a /da、 h b /d b分别为-2.38和-1.77, 均表现为超显性。多基因的加性累计效应[ d]为1.64, 显性效应[ h]为-0.90。BCP1、BCP2和F2分离世代群体总遗传率分别为77.03%、89.40%和91.92%, 其中主基因遗传率分别为47.63%、68.51%和79.45%, 多基因遗传率分别为29.40%、20.89%和12.47%, 主基因遗传率和多基因遗传率的比值分别为1.63、3.28和6.25, 表明角果长主要受2对主基因的影响。3个分离世代中环境因素引起的变异分别占22.97%、10.60%和8.08%。
控制果喙长的2对主基因加性效应( d a和 d b)分别是0.20和-0.20, 微效多基因加性效应[ d]为0.18, 显性效应[ h]为0.29。BCP1、BCP2和F2分离世代群体遗传率分别为73.79%、78.12%和79.85%, 其中主基因遗传率分别为33.71%、72.75%和52.25%, 多基因遗传率分别为40.08%、5.37%和27.60%。3个分离世代中环境因素引起的变异分别占26.21%、22.88%和20.15%。
角果在产量形成中具有重要的地位和作用, 是油菜产量构成的重要要素。本研究表明角果长度性状分为果身长(包含果喙长度)、有效角果长(不包含角果果喙的长度)和果喙长3个性状在2个亲本中差异均达到显著水平, F1代均接近于中双11, 和10D130差异达到显著水平, 3个性状中长角果较短角果为显性性状, 但是从3个世代分离群体的频次分布来看, 3个性状均表现为连续分布, 不应该简单分组, 因此应采用植物数量性状主基因+多基因混合遗传模型进行遗传分析。
在现有的甘蓝型油菜品种资源中, 果身一般长度为6~8 cm, 近年来由于部分长角材料的发现, 角果长度的遗传相继有报道。Chay等[ 7]利用经典遗传分析认为长角性状受2对主效基因的控制, 长角对短角为显性。危文亮[ 10]用6个世代的遗传分析后认为角果长性状在双亲间存在2对基因的差异, 遗传符合加性显性二基因互作模型, 存在显著的上位性效应, 且存在超显性。王艳惠等[ 11, 12]用世代均数法进行基因效应分析后认为, 角果长、果身长均符合加性-显性-上位性模型, 且以加性效应为主; 用主基因+多基因模型分析后认为, 2个组合的果身长、角果长均以C-0模型为最适遗传模型, 即受加性-显性-上位性多基因系统控制。我们用六世代群体联合遗传分析表明, 果身长、角果长和果喙长度均受2对基因控制, 其中角果长和有效角果长受2对加性-显性-上位性基因控制, 果喙长受2对加性效应为主的主基因控制, 同时3对性状也受微效多基因影响, 研究结果与以往研究也存在着差异[ 7, 8, 9, 10, 11, 12], 这可能与研究材料的遗传背景和研究方法不同有关。控制果身长的2对主基因加性效应分别为1.75和-0.06, 二者差异极大, 因此在果身长的选育过程中尤其要注重第1对基因的选择; 角果长的2对基因的加性效应值相等, 果喙长的2对主基因加性效应值作用方向相反, 大小相等, 这也为选育提供了理论依据。虽然F1群体3个角果性状均接近于长角果材料, 但是从果身长和角果长的显性效应值均为负值, 与预期相反, 这可能与2对基因间复杂互作和多基因作用有关。因此, 适当的育种方法, 如回交育种和杂交育种, 可将长角果有利基因转育到相应的育种材料中。由于角果长度相关基因存在着明显的显性效应和基因互作, 因此, 适宜的杂交组合可以获得强杂种优势。从遗传率来看, 该组合角果长、果身长和果喙长的遗传率均大于70%, 且其中主基因遗传率大于多基因遗传率, 这表明角果长性状主要受遗传因素影响, 环境作用较小, 因此, 早期选择角果长性状是有效的, 且在选择中应优先考虑主基因的作用。
在植物主基因+多基因混合遗传模型联合分析方法中, 确定一个基因位点属于主基因或多基因是由其表型效应大小来决定的, 一般情况下, 主基因具有较高的遗传效应值, 在分离群体表现出多峰。本文研究结果表明, 有效角果长和果喙长分别由2对主基因控制, 虽然角果果身长包含了有效角果长和果喙长, 但是仍然由2对主基因作用, 这说明同样的基因对不同的性状作用效应值不同, 也表明主基因和多基因之间没有明显的划分标准, 对决定相关性状的主基因数目不能简单地相加或者相减; 从另外角度看, 角果果身长、果喙长和角果长之间也可能受同一主基因影响。已经有的研究结果表明, 植物数量性状的遗传模型分析分离方法与QTL检测主基因的数量相对一致[ 33], 由于该方法所推论的基因仅仅是一个概念上的基因, 很难作个别比较, 为了更好地解释控制角果长3个性状的基因数目多少、效应值大小和是否为同一基因, 有必要利用分子标记进行QTL定位研究。目前, 我们正利用SSR等分子标记对F2群体开展角果长QTL分析, 初步研究结果已经证实“一因多效”的存在, 这为分子水平上进一步阐明这些基因的遗传效应、发掘与主基因紧密连锁的分子标记、实现油菜角果长分子标记辅助育种和基因克隆提供了前期研究基础。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|