{kind=link}
{kind=link}
以EMS诱变创制软质小麦宁麦9号高分子量谷蛋白亚基突变体
[张纪元, 张平平, 姚金保, 杨丹, 杨学明, 马鸿翔*  ]
]
]
|
|


本研究旨在创制遗传背景一致的不同高分子量谷蛋白亚基(HMW-GS)缺失系, 为小麦品质研究和育种提供材料。将软质小麦品种宁麦9号, 用0.4% EMS溶液处理10 000粒种子, 获得3781个M1代单株, 采用“半粒法”对每个株系进行SDS-PAGE鉴定, 从中筛选出299个(7.91%) HMW-GS突变株, 包括HMW-GS缺失和分子量突变2种类型。其中, HMW-GS缺失突变176株, 突变频率为4.65%, 缺失亚基涉及Ax1、Bx7、By8、Dx2和Dy12, 突变频率为0.24%~3.28%; 分子量突变130株, 突变频率为3.44%。将突变体的具胚端种子于温室中繁殖获得M2代, 再次鉴定各株系的HMW-GS, 并经M3代验证, 最终获得Ax1、Dx2、Bx7、By8和Dy12缺失突变体, 及Ax1+By8双缺失突变体。用高效液相色谱(HPLC)分析这些突变体的谷蛋白大聚体(GMP)和谷蛋白/醇溶蛋白(GLU/GLI)比值, 发现不同缺失突变体的GMP含量都有不同程度的降低, 尤以Bx7亚基缺失突变体中GMP含量降幅最大, 高达42%。另外, 不同缺失突变体的谷蛋白总量和GLU/GLI比值也低于对照, Ax1+By8双缺失突变体的GLU/GLI比值最小。
The objective of this study was to create a series of deletion lines of high molecular weight-glutenin subunit (HMW-GS) in similar backgrounds for wheat quality research and breeding. A total of 3781 M1 plants of Ningmai 9 (a soft wheat cultivar) were obtained from 10 000 seeds induced by 0.4% ethyl methane sulfonate (EMS). The HMW-GS compositions of these plants were identified via SDS-PAGE using half seed. Two hundred and ninety-nine lines with a mutation percentage of 7.91% were found to be HMW-GS mutants, including HMW-GS deletion and molecular weight mutation. Among them, 176 lines were HMW-GS deletion mutants, with a mutation percentage of 4.65% containing subunits Ax1, Bx7, By8, Dx2, and Dy12, with the mutation percentages ranging from 0.24% to 3.28%. The mutants of molecular weight were 130 lines with a mutation percentage of 3.44%. Each of the half M1 seed with embryo was used to produce M2 generation in greenhouse. Homozygous deletion lines of HMW-GS Ax1, Dx2, Bx7, By8, Dy12, and Ax1+ By8 were detected using SDS-PAGE and confirmed in M3 generation. The content of glutenin macropolymer (GMP) and the ratio of glutenin-to-gliadin (GLU/GLI) ratio were determined using high performance liquid chromatography (HPLC). The results showed that all the deletion mutants had lower GMP contents than Ningmai 9, particularly, the Bx7 deletion line had the reduced GMP content of 42%. The GLU/GLI ratio of the deletion lines was also smaller than that of Ningmai 9, and the lowest GLU/GLI ratio was found in Ax1+ By8 deletion line.
小麦是主要粮食作物之一, 品质改良是小麦育种的重要目标。麦谷蛋白是贮藏蛋白的主要成分, 对小麦的加工品质影响很大。根据十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE), 麦谷蛋白可分为高分子量谷蛋白亚基(HMW-GS)和低分子量谷蛋白亚基(LMW-GS)。HMW-GS虽然只占小麦贮藏蛋白的5%~10%, 但对小麦品质有重要影响, 并在很大程度上决定加工品质。HMW-GS与面团流变学特性和加工品质的关系已有大量报道, 研究结果在小麦育种中得到应用[ 1]。由于遗传背景对HMW-GS基因的表达有一定影响, 不同试验材料导致研究结果存在一定差异。因此创制背景一致的近等基因系或单一亚基缺失材料非常重要和必要, 目前亟待加强。
甲基磺酸乙酯(ethyl methane sulfonate, EMS)是一种可直接作用于DNA鸟嘌呤引起点突变及染色体损伤的化学诱变剂, 其诱变效果远远高于电离辐射和自发诱变, 目前应用最广泛、效果最好。利用EMS技术创制的突变体材料表现形态、生理、生化和抗病性变异, 是研究基因及其功能的良好材料。在小麦上, Feiz等[ 2]利用该技术创建了630株普通软质小麦突变体库; 李卫华等[ 3]研究了不同浓度EMS对新春11主要农艺性状和抗病性的诱变效应; Kuraparthy等[ 4]利用二倍体小麦EMS突变体库, 对控制分蘖的 tin3基因进行了染色体定位; 崔秋华等[ 5]用0.05%~0.80% EMS处理春小麦长春1号和吉麦2号幼穗, 发现染色体畸变率和微核率明显提高; 赵天祥等[ 6]用0.7%和1.2% EMS诱变偃展4110种子, 在M2代中发现了株高为10~15 cm的特矮变异; 徐艳花等[ 7]用0.8% EMS处理豫农201, 从M2代中鉴定出HMW-GS缺失1亚基、5亚基、7亚基和10亚基的突变体。
宁麦9号由江苏省农业科学院以扬麦6号为母本、日本品种西风为父本进行有性杂交, 采用改良集团法选育而成, 1997年通过江苏省品种审定, 是江苏省农业科学院育成的优质高产抗病弱筋小麦品种, 具有丰产性好、弱筋品质稳定、抗赤霉病和小麦黄花叶病等特点, 而且其穗粒数、弱筋品质指标和赤霉病抗性的一般配合力较高, 作为亲本已经育成12个小麦新品种。为了进一步利用该品种, 本研究以EMS处理进行诱变, 对其后代进行HMW-GS突变体的筛选及谷蛋白质量分析, 预测不同HMW-GS缺失对谷蛋白大聚体(glutenin macropoly-mer, GMP)的影响, 为HMW-GS对弱筋品质形成效应的研究以及弱筋小麦育种提供新的重要种质。
取1000粒完好的宁麦9号种子, 用清水在室温条件下浸种18 h, 然后换EMS-磷酸缓冲液缓慢振荡12 h, 再用流水反复冲洗种子12 h除去残余的EMS。EMS溶液(w/v)梯度为0.2%、0.4%和0.6%; 磷酸缓冲液(pH 7.0)含0.1 mol L-1 Na2HPO412H2O和0.1 mol L-1 NaH2PO42H2O。将种子晾干后置铺有滤纸的培养皿中, 保持滤纸湿润, 放入20℃光照培养箱中, 3 d后统计发芽率, 2次重复。以未处理宁麦9号种子为对照。
用最佳浓度EMS处理10 000粒种子, 经冲洗晾干后于2010年10月27日播于江苏省农业科学院试验地, 株距0.05 m, 行距0.23 m, 常规管理, 成熟后收获M1单株种子。采用“半粒法”将含胚的单粒M1种子播于温室中, 获得M2代, 继续采用单株种植获得M3代种子。
EMS浓度对宁麦9号的种子发芽率有显著影响, 随EMS浓度提高, 发芽率和相对致死率升高, 浓度0.4%接近半致死剂量(表1)。用0.4% EMS溶液处理10 000粒种子, 获得M1代3781单株, 由此建立了宁麦9号突变体库。田间调查表明, 200个对照行的平均田间出苗率为80%, 按成苗率计算, 0.4% EMS处理的致死率为52.7%。因此0.4%为最佳处理浓度。
| 表1 不同浓度EMS诱变处理的种子发芽率 Table 1 The percentages of seed germination affected by different concentrations of EMS |
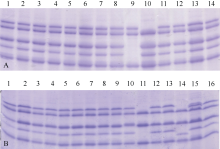
2.2.1 M1代HMW-GS突变筛选 SDS-PAGE检测表明, 宁麦9号的HMW-GS组成为(1, 7+8, 2+12)。在3781个M1单株中共检测到7种HMW-GS突变类型, 包括单亚基缺失的Ax、Bx、By、Dx和Dy缺失型及双亚基缺失的Bx+By和Ax+By缺失型(图1)。
 | 图1 以SDS-PAGE检测M1代HMW-GS缺失突变体A图为第3089至第3100号单株, 其中第9泳道为Bx+By双缺失, 第10泳道为Ax缺失; B图为第37至第51号单株, 其中第11泳道为Ax+By双缺失, 第14泳道为By分子量突变。Fig. 1 HMW-GS mutant detection of M1 plants using SDS-PAGEPanel A shows the SDS-PAGE result of lines from 3089 to 3100 plants, including Bx+By deletion (lane 9) and Ax deletion (lane 10). Panel B shows the SDS-PAGE result of lines from 37 to 51 plants, including Ax+By deletion (lane 11) and molecular weight mutant of HMW-GS By (lane 14). |
突变体库中HMW-GS缺失突变单株共299个, 突变频率为7.91%。其中HMW-GS分子量改变的材料(包括3份同时含Ax缺失的突变体)130份, 突变频率为3.44%。在172个(突变频率4.55%) HMW-GS缺失突变体中, Ax1亚基缺失频率最高, 共124份(包括3份Ax缺失+分子量突变及3份Ax+By双缺失), 突变频率为3.28%; 其他HMW-GS缺失突变型都低于15份, 突变频率低于0.4%, 尤以双缺失突变型最少(表2)。
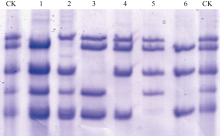
2.2.2 M2代HMW-GS分析 由M1代HMW-GS缺失突变体种子加代获得的M2代种子, 经SDS- PAGE检测, 仅发现25个株系在B、D位点控制的HMW-GS有缺失突变, 部分M1突变株系在M2代恢复正常, 表明部分M1代的突变体株系不能稳定遗传。M2代鉴定的Ax1、Bx7、By8、Dx2、Dy12单亚基缺失及Ax1+By8双亚基缺失突变株系能够稳定遗传至M3代, 且M3代基因型纯合(图2)。
| 表2 高分子量谷蛋白亚基突变体数目及频率 Table 2 The number and percentage of mutants on HMW-GS |
 | 图2 以SDS-PAGE检测M2代HMW-GS缺失突变体CK: 宁麦9号; 1: Ax1缺失突变体; 2: Dx2缺失突变体; 3: Bx7缺失突变体; 4: By8缺失突变体; 5: Dy12缺失突变体; 7: Ax1+By8双缺失突变体。Fig. 2 HMW-GS mutant of M2 lines detected using SDS-PAGECK: Ningmai 9; 1: Ax1 deletion mutant; 2: Dx2 deletion mutant; 3: Bx7 deletion mutant; 4: By8 deletion mutant; 5: Dy12 deletion mutant; 7: Ax1+By8 deletion mutant. |
不同HMW-GS缺失突变体间及其与对照间的GMP含量及GLU/GLI比值差异显著, 各突变体Ax、Dx、Bx、By、Dy和By+Dy缺失突变体的GMP含量分别比对照宁麦9号低9.18%、14.69%、17.79%、9.76%、14.86%和13.8%, 以Bx缺失突变体的降低量(14.69%)和降低百分率(42%)最大, Dy和Dx缺失突变体其次, Ax和By缺失突变体降幅最小。由于编码HMW-GS亚基的缺失, 导致谷蛋白总量的减少, 最终不同缺失突变体的GLU/GLI明显降低, Bx缺失类型的GLU/GLI比仅为1.07 (表3)。值得一提的是, Ax+By双缺失系的GMP降幅并非最大, 可能在这些缺失突变体中, 除HMW-GS缺失外, 还有其他影响谷蛋白和醇溶蛋白合成、翻译的相关基因突变。
| 表3 HMW-GS缺失突变体谷蛋白组成差异 Table 3 Variations of glutenin in HMW-GS deletion mutants |
麦谷蛋白是小麦籽粒贮藏蛋白的主要成分, 对加工品质起重要作用。高分子量麦谷蛋白是小麦种子贮藏蛋白的重要组成部分, 其数量和质量对小麦面筋含量及面粉烘烤品质有直接影响。因此, 小麦高分子量麦谷蛋白基因的表达和沉默备受关注[ 11]。利用同一品种诱变产生高分子量谷蛋白亚基缺失系, 可以克服不同品种遗传背景对实验结果的影响, 同时可以为小麦品质育种提供新的种质资源。EMS因其突变效率高、频率高和范围广等优点广泛用于农作物化学诱变[ 12], 一般使用半致死剂量作为最佳处理浓度[ 11]。在已报道的EMS诱变小麦构建突变体库试验中, 小麦品种各不相同, EMS处理浓度差异也很大, 有的用0.3%[ 13], 有的用0.8%[ 7], 还有的用1.2%[ 6]。本研究设计了3个EMS浓度梯度, 通过发芽率试验和田间出苗率验证, 认为0.4%是最佳处理浓度, 其种子发芽的致死率为49.9%, 成苗的致死率为52.7%, 接近半致死剂量。用该浓度EMS诱变宁麦9号10 000粒种子, 得到包含3781个株系的突变库。由此可见, 不同小麦品种对EMS敏感程度各异, 实际操作中有必要针对不同品种摸索最佳处理浓度。
利用EMS诱变构建突变体库, 其大小与目的基因大小有关。Feiz等[ 14]发现小麦每隔12 kb就会有一个突变体产生; Slade等[ 15]报道普通小麦平均每24 kb就有一个点突变出现, 而硬粒小麦每40 kb有一个点突变。本研究构建的突变体库包含3781个突
变株系, 编码7、8、2、12各亚基的基因分别为2388、2163、2508和1977 bp, 根据Feiz等[ 14]和Slade等[ 15]推算的突变频率, 该突变体库的大小不仅可满足对HMW-GS研究的需要, 而且可用于其他基因研究。徐艳花等[ 7]用0.8% EMS室温下诱变豫农201种子12 h, 从930个株系中鉴定出HMW-GS缺失突变株系21个, 突变频率2.26%。本研究从3781个突变株系中检测到299个HMW-GS突变株系, 突变频率为7.91%, 其中含有缺失突变的株系为172个, 突变频率为4.55%。突变频率的差异可能与品种等因素有关。
麦谷蛋白在面筋中以聚合体形式存在, 谷蛋白聚合体在非解离状态下由一系列大小不同的聚合体组成, 在SDS缓冲液中可分为可溶性和不溶性谷蛋白聚合体两类, 它们分别决定面团的延伸性和弹性, 两种聚合体的分子量和粒度大小显著不同[ 14], 其中不溶性谷蛋白分子量较大, 即GMP, 其含量与面团形成时间、面团最大抗延阻力、和面仪参数等显著相关[ 17, 18]。不同粒度的GMP均由HMW-GS和低分子量谷蛋白亚基(LMW-GS)组成, 但GMP中HMW/LMW比值较高[ 19, 20, 21]。因此, HMW-GS作为谷蛋白聚合体中的骨架, 其缺失会直接导致谷蛋白聚合体分子量的降低, 最终降低GMP含量。研究表明, GMP总的x/y-HMW比例较低, 谷蛋白聚合体中HMW-GS和y-HMW的相对含量提高时, GMP含量增加, 面筋强度加大[ 22, 23, 24]。我们构建的突变体库涉及5个HMW-GS缺失, 所有缺失突变体都导致GMP含量显著降低。由于HMW-GS的5个亚基中X型亚基数量显著大于Y型亚基, 其中Bx含量最高, 因此Bx缺失突变体导致GMP含量和GLU/GLI比值降低最多, 这可能有利于提高弱筋小麦加工品的质量。然而, 本研究仅考虑了HMW-GS 5个位点的缺失, 未分析LMW-GS以及影响谷蛋白表达的其他基因及加工品质, 这5种缺失类型与GMP和GLU/GLI比值的关系, 及对面粉及加工品质的影响仍需进一步探讨。此外, 本研究获得的含有不同谷蛋白亚基缺失突变的突变材料可作为育种材料直接利用, 也可通过彼此之间获得不同亚基组合的突变富集, 再应用于育种。
0.4% EMS处理种子12 h是诱变弱筋小麦品种宁麦9号的最适浓度。用此方法处理10 000粒种子, 获得包含3781株的突变体库, 对M1、M2和M3代进行SDS-PAGE检测, 获得遗传稳定、基因型纯合的Ax1、Bx7、Bx8、Dx2、Dx12单缺失及Ax1+By8双缺失的HMW-GS突变体, 不同缺失突变体的GMP含量和GLU/GLI比值均比宁麦9号有不同程度的降低。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|