
{kind=link}
{kind=link}
花铃期渍水对棉纤维糖代谢的影响及其与纤维比强度的关系
[杨长琴1, 2 , 刘瑞显2 , 张国伟2 , 杨富强2 , 周治国1, *  ]
]
]
|
|


以美棉33B为材料, 2007年和2009年在江苏南京池栽条件下设置渍水试验, 研究花铃期渍水对棉纤维糖代谢的影响及其与纤维比强度的关系。结果表明, 渍水条件下花后17~38 d纤维蔗糖和β-1,3-葡聚糖含量均值分别降低24.86%~81.30%和8.59%~36.30%, 纤维素最大累积速率在受渍害较轻条件下增加而受渍害较重条件下降低, 但快速持续期缩短, 成熟期纤维素含量降低, 比强度下降3.57%~10.03%。受渍害较轻条件下, 花后17~38 d纤维蔗糖合酶(SS)和蔗糖磷酸合酶(SPS)活性均值分别增加8.45%~24.59%和12.79%~18.20%, 生成碳源用于维持生存代谢, 而受渍害较重条件下分别降低7.06%~8.16%和11.40%~11.64%; 蔗糖酶和β-1,3-葡聚糖酶活性在花后17 d后降低。因此, 渍水条件下纤维加厚发育期碳源供应不足、碳代谢消耗增加或代谢被抑制是最终比强度下降的重要原因。
The experiments were conducted using a cotton cultivar NuCOTN 33B with pool culture under different waterlogging treatments in 2007 and 2009 to study effects of waterlogging on carbohydrate metabolism and its relationship with fiber strength during flowering and bolling stage in Nanjing, China. The results showed that the mean values of sucrose and β-1,3-glucan contents decreased by 24.86% to 81.30% and 8.59% to 36.30% respectively at 17 to 38 days after anthesis under waterlogging. The maximum rate of cellulose accumulation increased in response to mild waterlogging conditions, decreased in response to serious conditions, but the duration of maximum cellulose accumulation shortened, which resulted in the decrease in cellulose content. The value of fiber strength at mature period decreased by 3.57% to 10.03% under waterlogging. The mean values of SS and SPS activities increased by 8.45% to 24.59% and 12.79% to 18.20% respectively in response to mild waterlogging conditions, but decreased by 7.06% to 8.16% and 11.40% to 11.64% respectively at 17 to 38 days after anthesis in response to the serious waterlogging conditions, which were due to enhanced metabolic demands. Waterlogging decreased the activities of sucrase and β-1,3-glucanase after 17 days after anthesis. Therefore, insufficient carbon source, together with the increased carbon consumption or the inhibited sugar metabolism were potential factors contributing to low fiber strength for waterlogging plants.
纤维比强度是重要的原棉品质指标, 除基因型外, 环境因子如温度、土壤水分等对其影响也很大[ 1, 2]。纤维比强度取决于纤维发育过程中纤维素累积特性, 而纤维素的累积实质是纤维中糖代谢循环的过程[ 3, 4]。因此, 研究纤维发育过程中糖代谢生理特征及其对环境因子的响应, 对阐明纤维比强度的生态响应机制及高比强棉花栽培具有重要意义。
蔗糖是纤维发育糖代谢的初始底物, 决定着纤维素合成的直接底物——尿苷二磷酸葡萄糖(UDPG)的生成速率[ 5, 6]; 非纤维素物质β-1,3-葡聚糖(胼胝质)合成和降解也调节着纤维素的累积[ 7]。纤维糖代谢过程中蔗糖合酶分解蔗糖成UDPG和果糖, 为纤维素的合成提供直接底物[ 8], 蔗糖磷酸合酶利用胞质中游离态UDPG为供体催化合成蔗糖[ 8, 9], 间接地调节纤维素的合成。蔗糖酶水解蔗糖为葡萄糖和果糖, 为纤维素合成提供碳源和能量[ 8, 10]; 此外, β-1,3-葡聚糖酶水解胼胝质, 降解的产物在增加细胞壁可塑性同时为纤维素累积提供直接底物[ 11]。因此, 纤维发育过程中相关物质以及酶是影响糖代谢进程的重要因子。已有的研究表明温度[ 12]、氮肥[ 13, 14]影响纤维发育的物质和酶活动态, 改变纤维素的累积特性, 致使纤维比强度下降。但未见有关花铃期渍水影响纤维比强度生理机制的报道。本文通过研究花铃期渍水对纤维发育糖代谢影响及其与纤维比强度的关系, 揭示花铃期渍水影响纤维比强度的糖代谢生理机制, 可为生产上制定栽培调节措施以及高强纤维品种选育提供理论依据。
试验于2007年和2009年在江苏南京江苏省农业科学院防雨棚水泥池中进行, 供试土壤为黄棕壤土。两年0~20 cm土壤含有机质15.3 g kg-1和l4.3 g kg-1; 全氮0.8 g kg-1和0.9 g kg-1; 碱解氮63.4 mg kg-1和52.6 mg kg-1; 速效磷29.9 mg kg-1和23.8 mg kg-1; 速效钾107.5 mg kg-1和93.2 mg kg-1。供试品种为美棉33B。2007年4月17日播种、5月17日移栽, 2009年4月25日播种、5月22日移栽; 2年种植密度分别为每公顷3.60万株和3.90万株。
2007年设对照和渍水14 d 2个处理, 2009年设对照、渍水7 d和渍水14 d 3个处理。对照土壤相对含水量(土壤相对含水量=土壤含水量/田间持水量)保持在(75±5)%; 渍水处理保持地面2~3 cm水层, 渍水结束后土壤相对含水量自然减少至(75±5)%。于中部6~8台果枝开花时渍水处理, 开始处理时标记第1、第2果节当日花和花后10 d棉铃, 自花后10 d起每7 d取发育一致的棉铃6~8个(上午9:00至10:00取样), 将纤维与种子分离后, 液氮速冻并在-30℃下保存以供测定酶活性。标记棉铃吐絮时, 收取大小一致的棉铃约20个, 风干轧花后测定3.2 mm隔距纤维比强度。
用间苯二酚显色法[ 15]测定蔗糖含量; 用荧光分光光度计法[ 16]测定β-1,3-葡聚糖含量; 参照Updegraff方法[ 17]并略有改进, 测定纤维素含量。用蔗糖和UDP比色法测定蔗糖合酶(SS)活性(降解方向)[ 18], 果糖和UDPG比色法[ 15]测定蔗糖磷酸合酶(SPS)活性; 用DNS显色法[ 15]测定蔗糖酶活性; 用昆布多糖比色法测定β-1,3-葡聚糖酶活性[ 15]。用国产Y162A型束纤维强力机测定3.2 mm隔距比强度, 6次重复, 并用中国纤维检验局的标准棉样品修正。
用Microsoft Excel和SPSS 16.0软件分析数据。
表2表明, 花铃期渍水处理显著降低成熟期纤维比强度( P<0.05)。与对照相比, 渍水7 d处理起始于花后0 d和10 d, 成熟期纤维比强度分别下降4.99%和3.57% (2009); 渍水14 d处理起始于花后0 d, 成熟期纤维比强度下降6.87% (2007)和6.01% (2009), 起始于花后10 d时下降10.03% (2007)和9.27% (2009)。表明渍水7 d和渍水14 d处理分别对渍水起始于花后0 d和花后10 d棉铃纤维比强度影响较大。渍水处理间相比, 成熟期纤维比强度渍水7 d处理比渍水14 d处理高1.1%~6.3%。
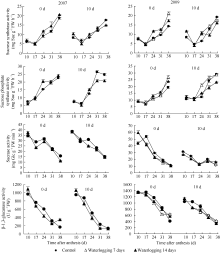
2.2.1 蔗糖含量 由图1可见, 对照纤维蔗糖含量呈单峰曲线变化, 峰值在花后17 d。渍水处理降低纤维加厚发育期蔗糖含量。渍水起始于花后0 d时, 渍水7 d和渍水14 d处理蔗糖含量分别于花后17~31 d和17~24 d显著降低( P<0.05); 起始于花后10 d时于花后24~31 d显著降低( P<0.05)。渍水处理间相比, 渍水起始于花后0 d时, 花后17~31 d蔗糖含量以渍水14 d处理较高; 起始于花后10 d时, 花后24~38 d蔗糖含量以渍水7 d处理较高。
2.2.2 β-1,3-葡聚糖含量 纤维β-1,3-葡聚糖含量呈单峰曲线变化, 峰值在花后17 d或24 d (图1)。渍水处理降低了峰值期β-1,3-葡聚糖含量, 如渍水起始于花后0 d时, 渍水7 d和14 d处理β-1,3-葡聚糖含量分别于花后17~31 d和17~24 d降低( P<0.05)。渍水处理间相比, 渍水起始于花后0 d时, 花后24~31 d β-1,3-葡聚糖含量以渍水14 d处理较高; 起始于花后10 d时, 花后24 d后β-1,3-葡聚糖含量以渍水7 d处理较高。
 | 图1 花铃期渍水对棉纤维蔗糖和β-1,3-葡聚糖含量的影响Fig. 1 Effects of waterlogging on changes of sucrose and β-1,3-glucan contents in fiber during flowering and bolling stage |
2.2.3 纤维素含量 纤维素含量呈慢-快-慢的累积动态, 用Logistic模型对纤维素累积进行模拟, 得到纤维素含量累积拟合方程均达到极显著水平。表1表明, 渍水处理的纤维素含量理论最大值随渍水持续期延长而降低; 纤维素含量快速增长期提前而持续期缩短, 渍水7 d处理纤维素最大累积速率均增加, 渍水14 d处理起始于花后0 d时最大速率增加而起始于花后10 d时降低。渍水处理间相比, 随渍水持续期延长, 纤维素快速累积持续时间延长, 但最大速率降低, 纤维素累积量降低。
2.3.1 蔗糖合酶(SS)和蔗糖磷酸合酶(SPS) 由图2可见, SS活性呈先降后升的变化, 渍水处理对其影响与渍水持续期及起始时花后时间有关。渍水起始于花后0 d时, 渍水7 d和14 d处理SS活性分别于花后38 d和24~38 d增加( P<0.05); 而起始于花后10 d时分别于花后24~31 d增加( P<0.05)和花后31 d后降低( P<0.05)。渍水处理间相比, 起始于花后0 d时, 花后24~31 d SS活性以渍水14 d处理较高; 而起始于花后10 d时则以渍水7 d处理较高。渍水对纤维SPS活性的影响与SS相似。
| 表1 花铃期渍水对纤维素累积特征值的影响 Table 1 Effect of waterlogging on the characteristics of cellulose accumulation in fiber during flowering and bolling stage |
2.3.2 蔗糖酶活性 蔗糖酶活性随花后时间增加呈下降动态(图2)。渍水起始于花后0 d时, 花后17~24 d纤维蔗糖酶活性显著降低( P<0.05); 起始于花后10 d时, 花后10~17 d蔗糖酶活性显著降低( P<0.05)。渍水处理间差异小。
2.3.3 β-1,3-葡聚糖酶活性 由图2可见, β-1,3-葡聚糖酶活性随花后时间增加呈下降动态。渍水处理降低了加厚发育期纤维β-1,3-葡聚糖酶活性。如渍水起始于花后10 d时, 渍水7 d和14 d处理β-1,3-葡聚糖酶活性分别于花后17~24 d和17~31 d显著降低( P<0.05)。渍水处理间差异与β-1,3-葡聚糖含量差异相似。
表2表明, 与对照相比, 渍水处理花后17~38 d蔗糖、β-1,3-葡聚糖含量均值降低。渍水7 d处理起始于花后0 d时花后17~38 d蔗糖和β-1,3-葡聚糖含量均值分别降低37.44%和36.30%; 起始于花后10 d时分别降低42.64%和8.59%。渍水14 d处理起始于花后0 d时花后17~38 d蔗糖和β-1,3-葡聚糖含量均值分别降低25.04%和25.06% (2009); 起始于花后10 d时分别降低81.30%和18.32% (2009)。渍水处理间相比, 渍水起始于花后0 d时, 纤维蔗糖和β-1,3-葡聚糖含量均值以渍水14 d处理较高; 起始于花后10 d时则以渍水7 d处理较高。结合前述渍水处理对纤维比强度的影响, 可见渍水下蔗糖含量下降与纤维比强度的结果一致。表2还表明, 渍水条件下蔗糖转化率随渍水持续期延长增加; 渍水14 d处理起始于花后10 d时纤维素快速累积期最大速率降低, 其他处理均增加; 纤维素含量和纤维比强度均随渍水持续期延长降幅增大。表明渍水下蔗糖转化并非都用于纤维素的合成, 纤维素过快和过慢累积均不利于提高纤维比强度。
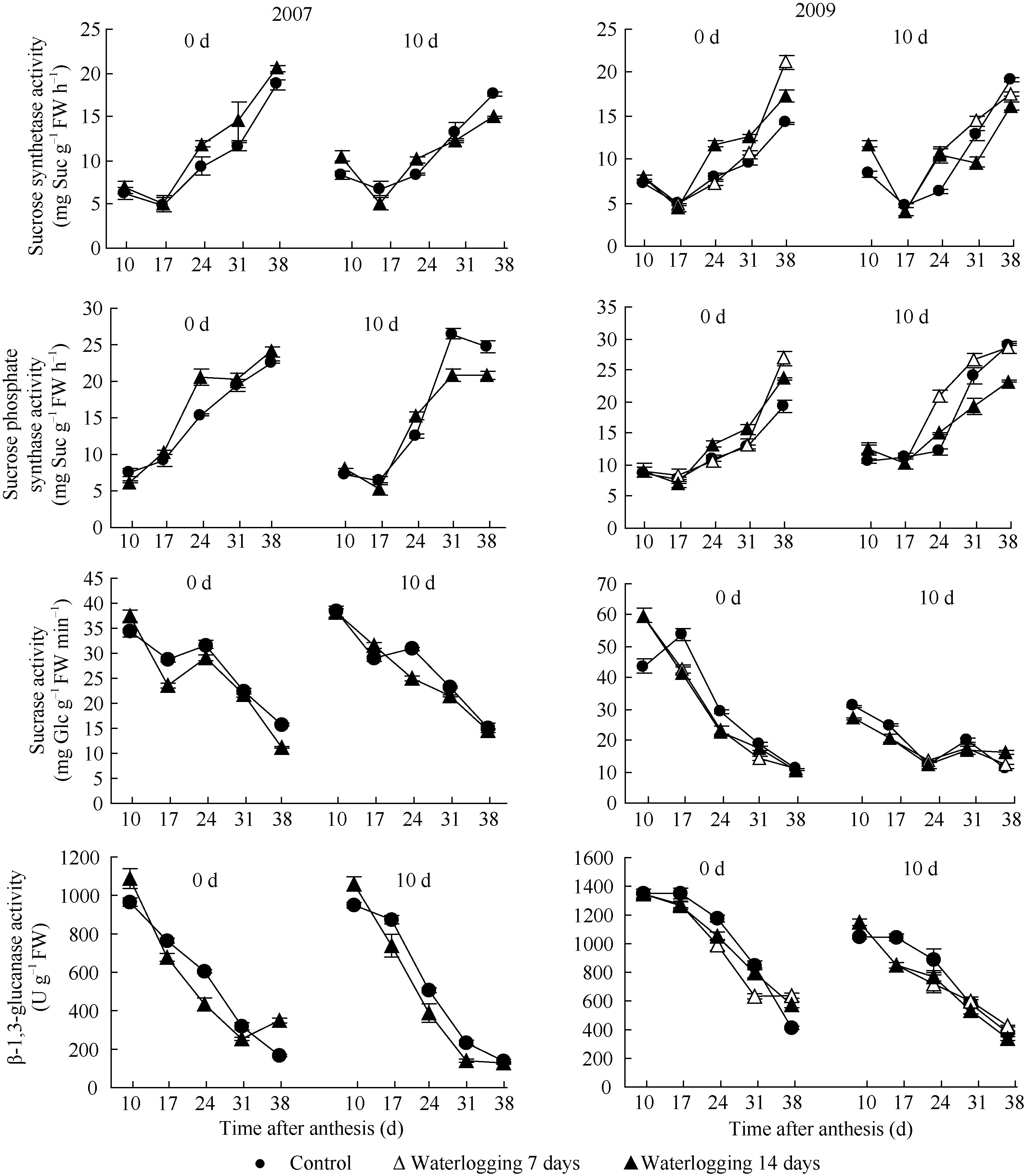 | 图2 花铃期渍水对棉纤维糖代谢相关酶活性的影响Fig. 2 Effects of waterlogging on enzyme activity associated with sugar metabolism in fiber during flowering and bolling stage |
| 表2 纤维加厚发育期(17~38 d)物质、酶活与成熟期纤维比强度的关系 Table 2 Relationship between change of key matters and enzyme activities (17-38 days after anthesis) associated with sugar metabolism and fiber strength during cotton fiber thickening stage |
与对照相比, 渍水7 d处理起始于花后0 d时花后17~38 d纤维SS和SPS活性均值分别增加19.61%和16.69%, 起始于花后10 d时分别增加8.45%和12.79%。渍水14 d处理起始于花后0 d时花后17~38 d纤维SS和SPS活性均值分别增加24.60%和18.20% (2009), 起始于花后10 d时分别降低7.06%和11.64% (2009), 表明SPS和SS活性变化与渍水持续期和起始时间有关。SPS和SS活性增加时受渍害较轻, 纤维比强度下降3.57%~6.86%; SS、SPS活性降低时受渍害较重, 比强度下降9.28%~10.03%。此外, 渍水处理的蔗糖酶和β-1,3-葡聚糖酶活性均值呈降低趋势。
蔗糖是纤维糖代谢的起始底物, 胡宏标等[ 7]研究表明纤维发育过程中蔗糖含量和转化率高有利于纤维比强度的提高。本试验渍水条件下由于棉铃对位叶碳源持续供给能力降低(另文发表), 花后17~38 d纤维蔗糖含量均值比对照降低24.86%~81.30%, 且不同处理蔗糖含量下降的幅度与纤维比强度下降的幅度一致, 纤维加厚发育期β-1,3-葡聚糖含量也下降。因此, 加厚发育期纤维蔗糖、β-1,3-葡聚糖含量降低是纤维比强度降低的原因。与Pettigrew等[ 19]遮阴下纤维加厚发育期(花后21 d)蔗糖含量降低影响最终纤维强度的观点一致, 与低温下蔗糖转化率降低[ 12]是棉花纤维素合成受阻的主要原因结果不太一致。本试验渍水处理蔗糖转化率增加, 致使纤维素快速累积期提前、但持续期缩短, 不同处理纤维素累积最大速率增加或降低, 推测与受渍害程度有关。表明纤维素平缓累积有利于纤维比强度提高, 而过快[ 4, 7]、过慢累积均不利于纤维比强度提高。渍水对纤维物质的影响与渍水持续期及起始时间有关。渍水起始于花后0 d时, 纤维蔗糖和β-1,3-葡聚糖含量以渍水7 d处理受影响较大; 起始于花后10 d时, 则以渍水14 d处理受影响较大, 而纤维素含量、纤维比强度随渍水持续期延长而降低。植物在逆境条件下体内的碳水化合物首先用于维持呼吸和生存能量需求[ 20], 推测渍水下纤维物质并非都用于纤维素合成, 随渍水持续期延长, 用于维持生存代谢的消耗增多。
纤维发育相关物质受酶活调控。已有的研究表明纤维加厚发育期SS活性增加的同时SPS活性也增加, 直接或间接影响纤维素合成[ 8]。另有研究发现, SS是渍水条件下蔗糖降解的主要酶[ 21, 22, 23], 可为植物在低氧胁迫下提供足够的糖[ 24]。本试验中渍水7 d与渍水14 d (起始于花后0 d)处理花后17~38 d纤维SS和SPS活性均值分别增加8.45%~24.59%和12.79%~18.20%, 但最终纤维素累积量降低, 表明有纤维素合成以外的碳源消耗途径存在。Haigler等[ 8, 25]推测逆境下碳源供应减少且膜结合型SS向可溶型转化, 增加细胞质中游离UDPG浓度用于维持基本代谢过程, 纤维素停止高速合成。纤维发育中蔗糖主要来源于棉铃对位叶, 渍水下棉铃对位叶蔗糖代谢酶活性均被激活, 蔗糖累积而铃重降低(另文已发表)。可见, 渍水条件下棉铃对位叶、棉铃中蔗糖代谢均被激活但生成碳源并非都用于棉铃发育。花铃期渍水条件下, 通过栽培措施补给碳源、促进碳源有效利用有利于改善铃重和纤维品质。本试验中, 渍水7 d处理起始于花后10 d时纤维SS、SPS活性响应快于起始于花后0 d时; 渍水14 d处理起始于花后0 d时, 花后17~38 d SS和SPS活性均值增加而起始于花后10 d时降低。推测SS与SPS活性响应渍水的变化与受渍害程度有关。受渍害较轻时SS和SPS活性响应快且酶活增加、纤维比强度下降幅度较小; 受渍害较重时酶活性降低, 纤维比强度下降幅度大。此外, 渍水条件下根系是直接受害的器官, 根部的碳水化合物大部分用于储存, 是恢复生长阶段根和叶重新生长的重要物质来源[ 26]。因此, 生产中及时清沟理墒, 促进根部恢复生长是产量和品质提高的保证。
花铃期渍水下, 纤维加厚发育期蔗糖、β-1,3-葡聚糖含量降低; 纤维SS和SPS活性受渍较轻条件下被激活而受渍较重条件下降低, 蔗糖酶、β-1,3-葡聚糖酶活性也降低。渍水下碳源供应不足, 糖代谢消耗增加或受抑制是最终纤维比强度降低的原因。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|