{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
内源CTK和IAA平衡对水稻分蘖芽休眠与萌发的影响
[罗宝杰* , 许俊旭* , 丁艳锋, 李刚华, 刘正辉, 王绍华*  ]
]
]
|
|


* 通讯作者(Corresponding author): 王绍华, E-mail:wangsh@njau.edu.cn; Tel: 025-84396475
** 同等贡献(Contributed equally to this work)
研究水稻抽穗期分蘖休眠与萌发转换过程中CTK和IAA含量及CTK/IAA值的变化, 探讨内源激素平衡对水稻分蘖的调控作用。采用外施6-BA打破分蘖芽休眠和去穗后外施IAA抑制分蘖芽萌发两种处理, 创建分蘖芽休眠与萌发转换的材料, 测定伸长节和伸长节分蘖芽中CTK和IAA的含量。6-BA处理打破了分蘖芽的休眠, 显著提高了倒二节和倒二芽中CTK含量, 降低了倒二节中IAA含量, 对倒二芽中IAA含量影响不大, 显著提高了节、芽中CTK/IAA值。去穗后外施IAA逆转了去穗对节、芽CTK含量提高效果, CTK含量维持在较低值, 显著提高了倒二节中IAA含量, 对倒二芽中IAA含量无显著影响, 显著降低了节、芽中CTK/IAA值。CTK和IAA共同调控分蘖发生, 较高的CTK/IAA值利于分蘖芽萌发, 较低的CTK/IAA值使其休眠, 节中CTK/IAA值在2.6~2.8, 芽中CTK/IAA值在4.8~5.0, 可作为分蘖芽休眠与萌发转换的临界指标。
The objection of this experiment was to study the changes of contents of CTK, IAA and the ratio of CTK/IAA during heading stage, and to investigate the regulation of endogenous hormones in rice tiller. Exogenous 6-BA and IAA were used to regulate the development of tiller buds, establish the transfer between dormancy and germination, and measure the contents of CTK and IAA in elongated nodes and tiller buds located at the elongated nodes. The exogenous 6-BA increased the CTK contents in nodes and tiller buds, decreased the IAA content in nodes had little effect in the content of IAA in tiller buds, significantly increased ratios of CTK/IAA in both nodes and tiller buds, and promoted the transfer from dormancy to germination. External IAA after removing panicle decreased CTK contents in nodes and tiller buds, which were maintained at a low level, increased the IAA content in buds, and significantly decreased the ratio of CTK/IAA in nodes and tiller buds, causing the transfer from germination to dormancy. Tiller development is determined by the balance between CTK and IAA, higher CTK/IAA ratio is benefit to the transfer from dormancy to germination, while lower CTK/IAA ratio inhibits the transfer. The ratios of CTK/IAA at 2.6-2.8 in nodes, 4.8-5.0 in tiller buds can be considered as the critical exponent of the transfer between dormancy and germination.
分蘖是水稻等禾谷类作物最重要的农艺性状之一, 分蘖芽的萌发生长直接影响群体的构建及产量[ 1]。水稻分蘖的发生受到遗传因素、环境条件和激素等的多重影响[ 2], 有研究表明, 环境因素也是通过改变水稻内源激素含量及其平衡进而影响分蘖的发生[ 3, 4, 5]。因此, 内源激素在分蘖发生过程中起着重要作用。前人对于分蘖生长过程中内源激素含量的变化进行了大量的研究。细胞分裂素(CTK)和生长素(IAA)共同调节分蘖芽的休眠与萌发, 细胞分裂素促进分蘖芽的萌发[ 6, 7, 8], 而生长素则抑制分蘖芽的萌发[ 9, 10, 11]。李经勇等[ 12]认为水稻叶片和穗中IAA含量降低, 叶鞘、茎秆和再生芽中IAA含量升高, 有利于再生芽萌发。马兴林等[ 13, 14]研究发现, 在冬小麦衰亡前和衰亡过程中, IAA/(Z+ZR)值持续增大。李春喜等[ 15]研究表明小麦分蘖发生与分蘖节中Z+ZR含量呈显著正相关。刘杨[ 16]的研究结果显示, 在齐穗期去除顶穗, 水稻节和芽中Z+ZR和IP+IPR含量显著提高, 是分蘖芽休眠向萌发转换的前提条件。前人对水稻分蘖芽休眠与萌发转换过程中内源激素的变化虽然进行了大量研究, 但在CTK和IAA平衡与水稻分蘖芽休眠和萌发的关系方面研究较少, 且未能明确水稻分蘖芽实现休眠与萌发相互转换的临界CTK/IAA值。本研究通过叶面喷施不同浓度外源激素, 调控水稻内源CTK和IAA水平, 分析了CTK和IAA平衡与分蘖芽休眠和萌发的关系。旨在探明水稻分蘖芽休眠与萌发转换的CTK/IAA值, 为更好地调控水稻分蘖发生提供生理依据。
供试品种为南粳44 (常规粳稻)和扬稻6号(常规籼稻)。南京农业大学丹阳试验基地供试土壤质地为黏土, 含有机质21.0 g kg-1、碱解氮80.3 mg kg-1、速效磷15.6 mg kg-1、速效钾80.9 mg kg-1。2011年5月27日播种, 6月23日移栽; 2012年5月27日播种, 6月26日移栽。行株距为30.0 cm × 13.3 cm, 双苗移栽。小区面积为5 m × 4 m, 随机区组排列, 3次重复, 田间管理与大田生产相同。在抽穗期分别进行正、反两组试验。正试验以叶面喷施6-BA诱导伸长节休眠分蘖芽萌发。即在留穗的条件下, 设置叶面喷施15 (BA-15)、20 (BA-20)、25 (BA-25)、30 (BA-30) mg L-1 6-BA四个处理, 以叶面喷施清水为对照。反试验参照刘杨[ 16]的方法, 先去穗诱导伸长节休眠分蘖芽转入萌发, 去穗12 h后再叶面喷施IAA逆转去穗效应, 使已转入萌发的分蘖芽再度转入休眠。即在去穗12 h的条件下, 设置叶面喷施30 (IAA-30)、40 (IAA-40)、50 (IAA-50)、60 (IAA-60) mg L-1 IAA四个处理, 以叶面喷施清水为对照。
1.2.1 分蘖芽长度 从叶面喷施6-BA或去穗开始取样, 每间隔12 h取一次, 共取7次, 直至处理72 h分蘖芽完全实现休眠与萌发转换[ 16]停止取样。每次每小区取40个单茎, 快速剥去叶和鞘, 用刀片割下倒数第2叶着生的节和分蘖芽, 简称倒二节和倒二芽, 用直尺测量芽长, 然后将倒二节和倒二芽分别装入样品盒, 以液态氮速冻保存, 备测激素含量。
1.2.2 激素含量测定 按照杨建昌等[ 17]的方法提取和纯化样品中IAA和CTK, 其中CTK组分包含玉米素+玉米素核苷(Z+ZR)和异戊烯基腺嘌呤+异戊烯基腺嘌呤核苷(ip+ipR), 采用酶联免疫法测定其含量[ 18]。试剂盒购自中国农业大学, 使用酶标仪为Spectramax plus型。
使用Microsoft Excel 2003软件处理数据与统计分析。由于2011年和2012年试验结果的变化趋势基本相同, 本文将2年数据合并分析。
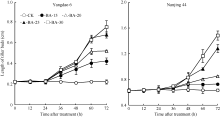
在正试验中, CK倒二芽在72 h观察期内未出现生长现象, 倒二芽的长度始终保持在0 h水平(图1)。不同浓度6-BA处理倒二芽的长度从24 h (扬稻6号)或36 h (南粳44)开始明显增长, 并显著高于同期CK, 且随喷施6-BA浓度提高, 倒二芽的芽越长。表明叶面喷施不同浓度6-BA均打破倒二芽的休眠, 实现分蘖芽从休眠向萌发的转换。
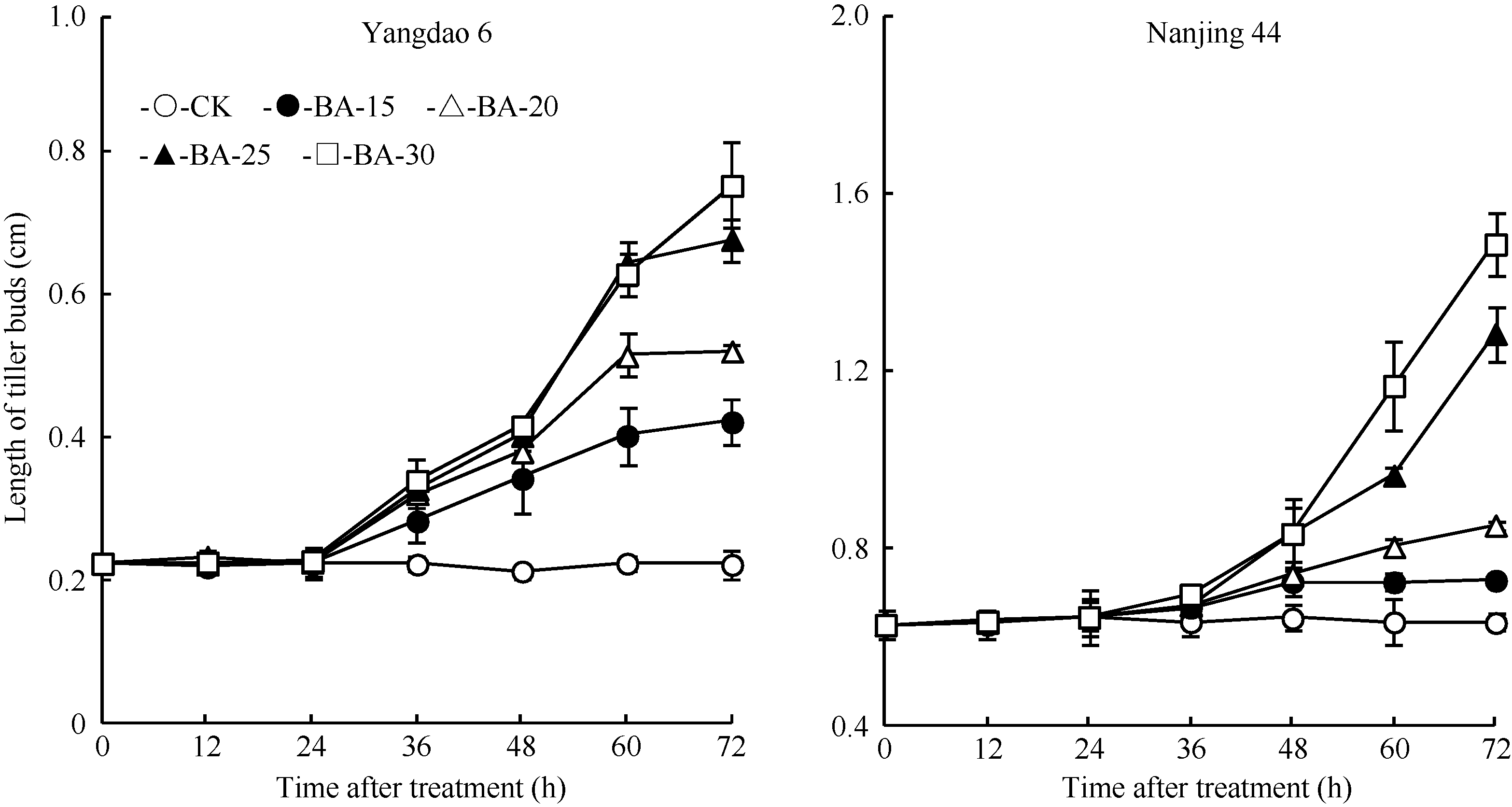 | 图1 留穗施用外源6-BA对倒二芽长度的影响Fig. 1 Effect of external 6-BA on the length of tiller buds located at elongated nodes |
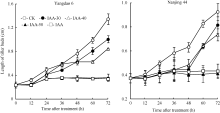
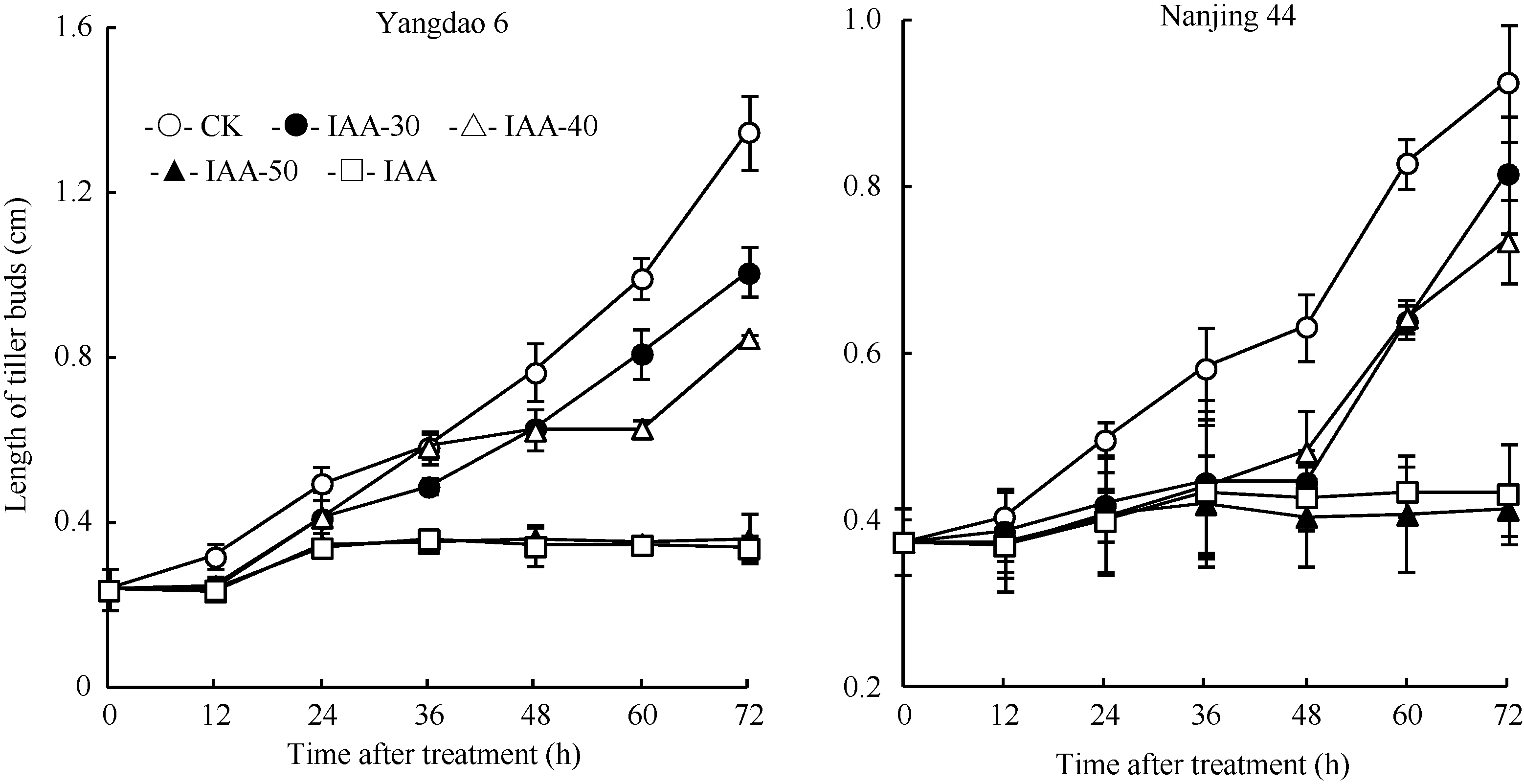 | 图2 去穗12 h再施用外源IAA对倒二芽长度的影响Fig. 2 Effect of external IAA on the length of tiller buds located at elongated nodes after panicle removing |
在反试验中, CK倒二芽在去穗后12 h的芽长已开始明显长于0 h, 分蘖芽转入萌发生长状态(图2)。叶面喷施不同浓度IAA处理对转入萌发态倒二芽的伸长生长具有明显抑制作用, 且浓度越高抑制作用越强。其中, IAA-50和IAA-60两处理在喷施IAA后倒二芽的芽长未明显增长, 去穗诱导倒二芽萌发生长的效应几乎被完全抑制; 而IAA-30和IAA-40两处理由于喷施浓度较低, 喷施IAA后倒二芽仍然有一定程度的增长, 未能完全逆转去穗的效应。表明叶面喷施50 mg L-1或60 mg L-1 IAA可以完全逆转去穗的萌发生长效应, 实现分蘖芽从萌发向休萌转换。
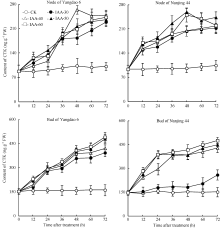
正试验表明(图3), CK倒二芽和倒二节中的CTK含量基本维持稳定; 不同浓度6-BA处理12 h后倒二节和倒二芽中CTK含量持续显著升高, 到分蘖芽开始伸长的36 h, 6-BA各处理倒二节和倒二芽中CTK含量分别比CK显著高77.7%~156.3%和86.7%~174.7%不等。反试验的结果与正试验相反(图4), 去穗后倒二节中CTK含量显著增加, 并在36 h达到高峰, 比0 h显著高426.4%; 不同IAA处理均减缓了倒二节中CTK含量的升高, 其中IAA-50和IAA-60两处理的CTK含量在处理后24 h达到最高值, 仅比0 h高114.9%左右, 随后开始下降, 呈现先升高再降低的趋势, 并最终维持在一个较低的水平, 与0 h含量无显著差异。倒二芽中CTK含量变化幅度略小, 但趋势与倒二节基本一致。这说明较高浓度的IAA处理完全逆转了去穗对CTK含量增加的促进作用, 2个品种呈现相同的趋势。
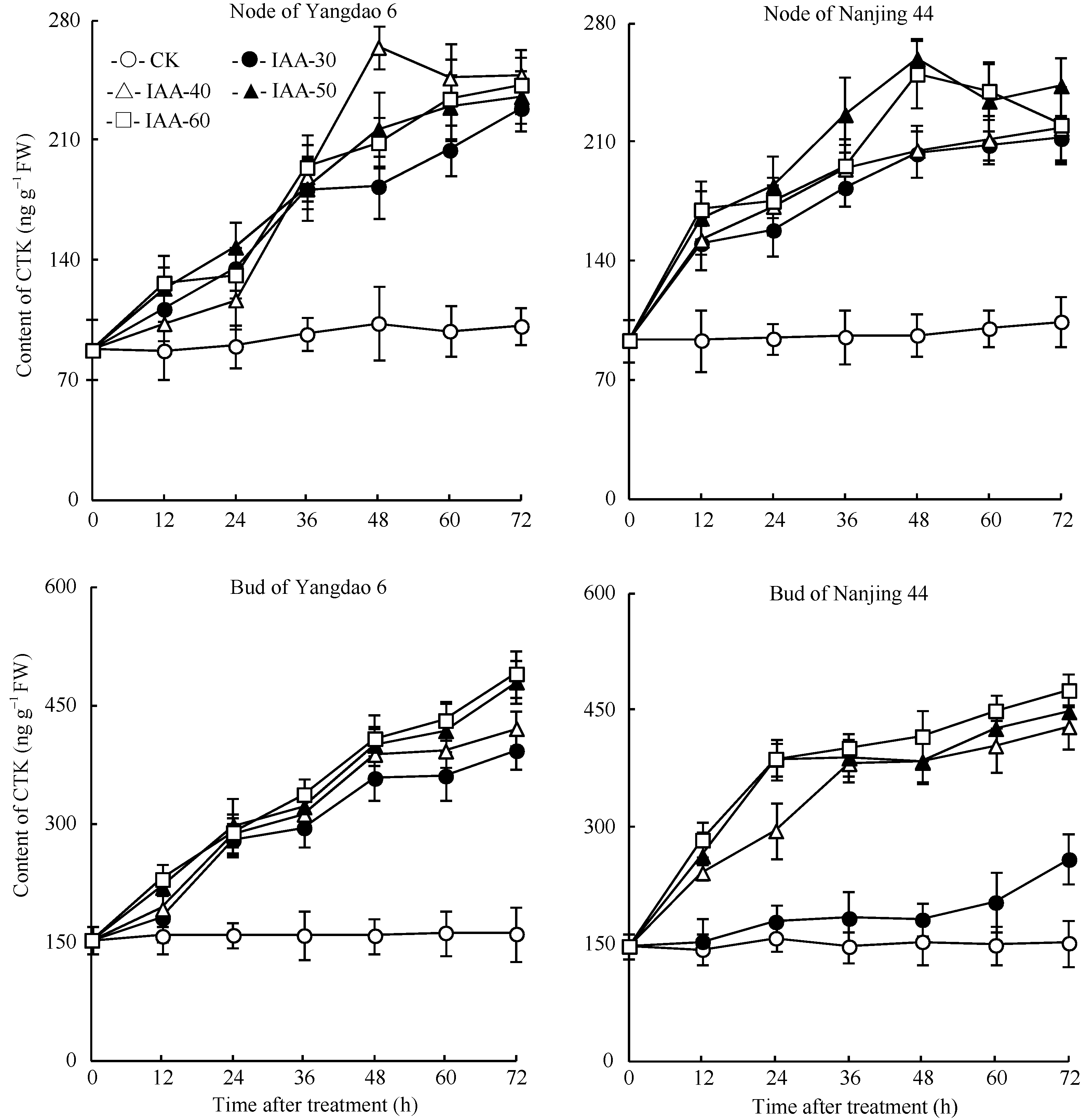 | 图3 留穗施用外源6-BA对节和芽中CTK含量的影响Fig. 3 Effect of external 6-BA on CTK contents of nodes and tiller buds |
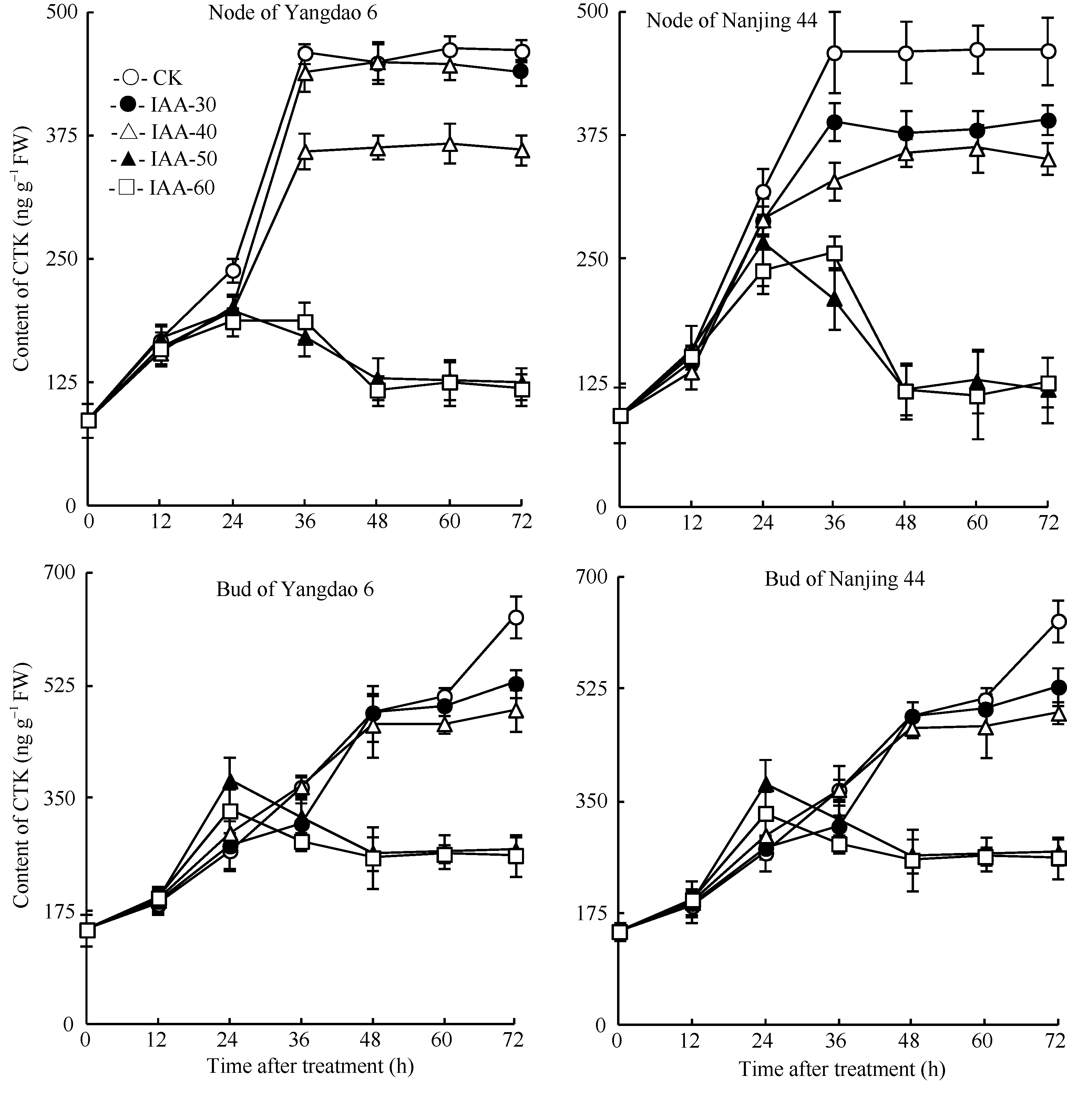 | 图4 去穗12 h再施用外源IAA对节和芽中CTK含量的影响Fig. 4 Effect of external IAA on CTK contents of nodes and tiller buds after 12 hours of panicle removing |
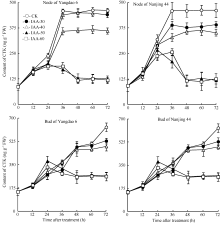
正试验表明(图5), 在观测期内CK的IAA含量基本保持不变, 不同浓度6-BA处理12 h后倒二节中IAA含量持续显著下降, 并于48 h后维持在一个较低的水平, 扬稻6号和南粳44的最低含量分别仅为初始值的51.7%和71.0%。与倒二节不同, 不同浓度6-BA处理倒二芽中IAA含量在观测期内无显著变化, 基本维持在一定的水平。反试验结果与正试验相反(图6), 去除顶穗后, CK倒二节中IAA含量显著下降, 72 h后倒二节中IAA含量仅为初始值的31.7%。不同浓度IAA处理的倒二节中IAA含量均高于同期CK, 表明外源IAA减缓了倒二节中IAA含量的下降, 其中IAA-30和IAA-40两处理内源IAA含量也持续下降, 与CK相比降幅减缓, 但未完全抑制IAA下降; IAA-50和IAA-60两处理在去穗后0~48 h (外施IAA后0~36 h)内源IAA含量持续下降, 于去穗后48 h (外施IAA后36 h)停止下降, 之后维持在较高水平, 扬稻6号和南粳44分别为初始值的70.0%和68.3%, 表明高浓度的IAA处理有效地抑制了去穗对内源IAA含量的下降作用。与倒二节不同, 外施不同浓度IAA倒二芽中IAA含量变化无明显规律, 2个品种表现一致。
正试验结果表明, 留穗外施15、20、25和30 mg L-16-BA, 倒二分蘖芽在24 h已从休眠转入萌发, 15 mg L-1 6-BA为诱发这种转换的最低剂量(图1)。在15 mg L-1 6-BA处理后24 h, 南粳44和扬稻6号倒二节中CTK/IAA比值已升高到2.8和2.7, 明显高于CK的1.5和1.5 (表1); 倒二分蘖芽内CTK/IAA比值已升高到5.0和4.7, 明显高于CK的2.7和2.7 (表2)。反试验结果说明, 去穗后48 h倒二分蘖芽已充分萌发, 外施50 mg L-1和60 mg L-1 IAA到48 h已萌发分蘖芽的生长明显被抑制, 转入休眠状态, 50 mg L-1 IAA为诱发这种转换的最低剂量(图2)。从表3和表4可以看出, 在50 mg L-1 IAA处理后48 h, 南粳44和扬稻6号倒二分蘖芽内CTK/IAA比值已降低到4.4和4.9, 明显低于CK的8.9和7.6; 节中CTK/IAA比值已降低到2.6和2.8, 明显低于CK的13.2和12.8。
因此, 芽内CTK/IAA比值在4.8~5.0, 节中CTK/IAA比值在2.6~2.8, 可作为分蘖芽休眠与萌发转换的临界值。
| 表1 留穗外施6-BA对倒二节CTK/IAA的影响 Table 1 Effect of external 6-BA on ratio of CTK/IAA in nodes |
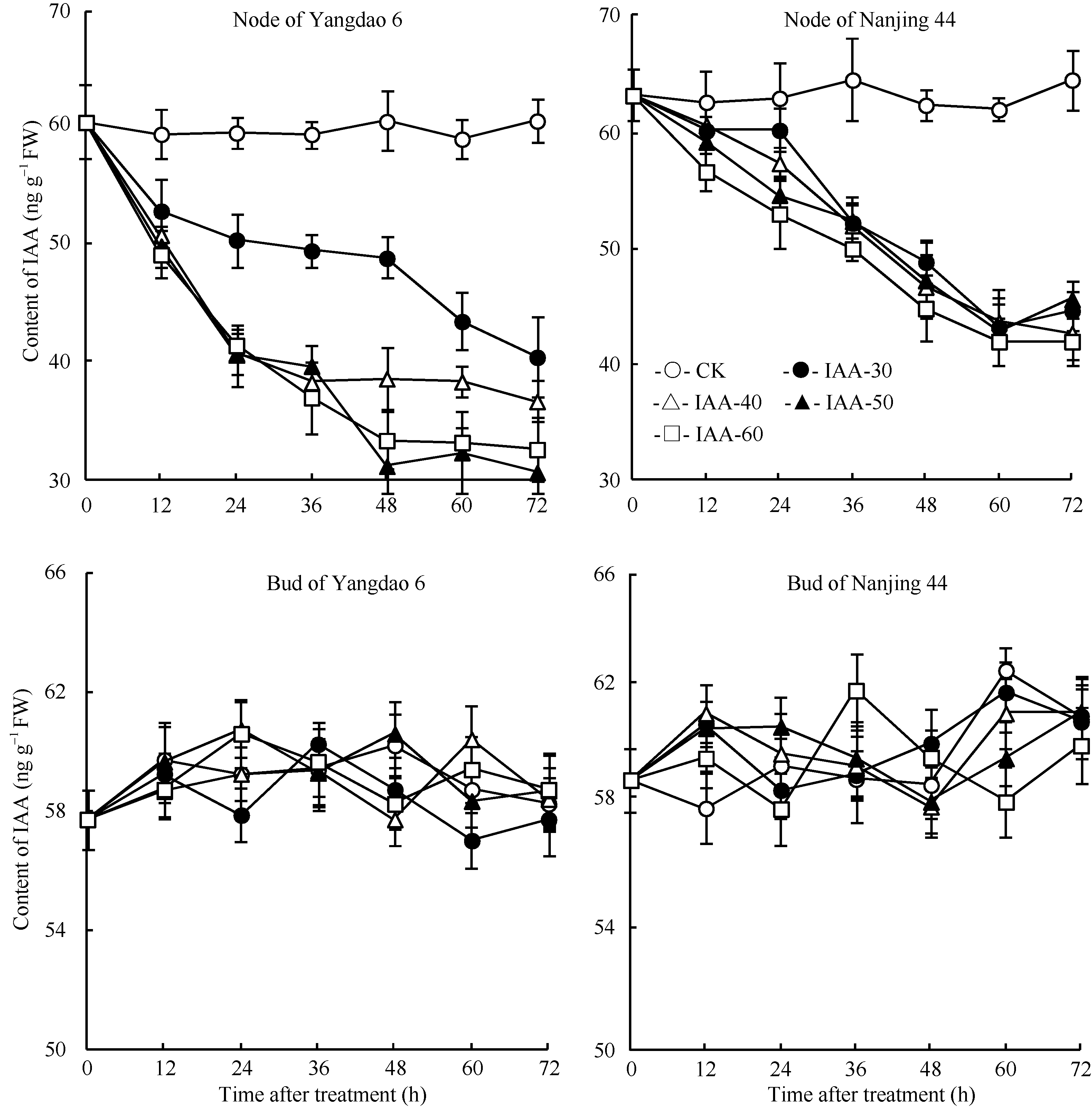 | 图5 留穗施用外源6-BA对节和芽中IAA含量的影响Fig. 5 Effect of external 6-BA on IAA contents of nodes and tiller buds |
| 表2 留穗外施6-BA对倒二芽CTK/IAA的影响 Table 2 Effect of external 6-BA on ratio of CTK/IAA in buds |
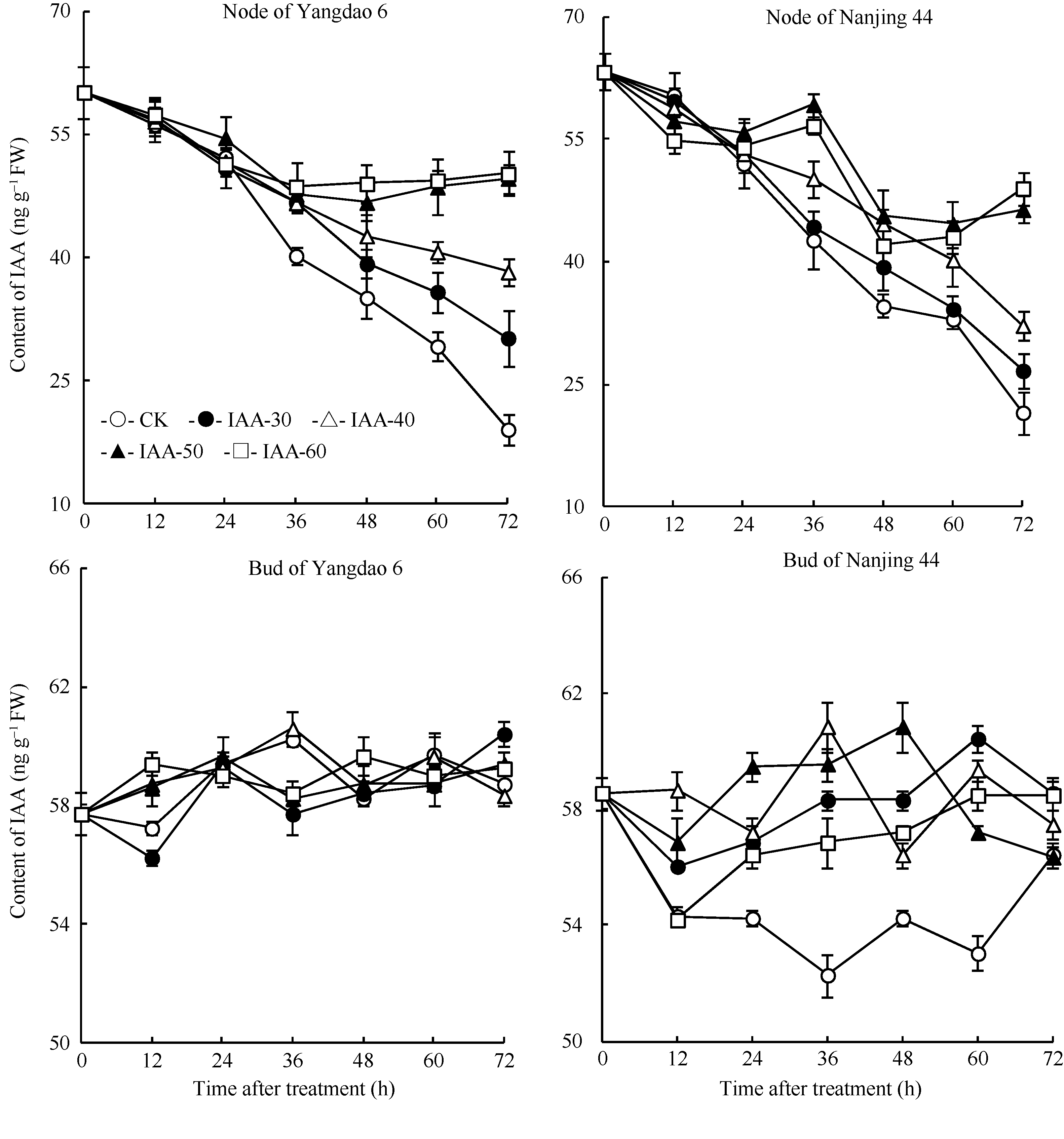 | 图6 去穗12 h再外源施用IAA对节和芽中IAA含量的影响Fig. 6 Effect of external IAA on the IAA contents of the nodes and tiller buds after 12 hours of panicle removing |
| 表3 去穗12 h再外施IAA对倒二节CTK/IAA的影响 Table 3 Effect of external IAA on ratio of CTK/IAA in nodes after panicle removing |
| 表4 去穗12 h再外施IAA对倒二芽CTK/IAA的影响 Table 4 Effect of external IAA on ratio of CTK/IAA in buds after panicle removing |
植物激素可以调控分蘖发生, 本研究显示, 外施6-BA可以打破分蘖芽的休眠, 使其向萌发转换; 较高浓度的IAA可以逆转去穗对分蘖芽的伸长效应, 使其转向休眠, 说明外源激素改变了分蘖芽的休眠与萌发状态, 转换材料创建成功。除遗传因素[ 19, 20]和外界环境[ 21]外, 植物激素对分蘖发生有重要作用。Thorne等[ 4]研究发现, 环境因素是通过改变内源激素含量进而影响分蘖生长。CTK在根部或茎部合成, 运输至侧芽, 促进细胞分裂和扩大。刺激叶片中蛋白质和核酸的合成, 直接促进分蘖发生[ 3, 22]。外源施用CTK促进小麦侧芽生长[ 23], 较高的CTK含量有利于小麦、玉米等分蘖的发生[ 13, 24], 小麦分蘖衰亡过程与Z+ZR含量密切相关[ 13, 14]。本试验研究结果显示, 分蘖芽开始萌发之前, 倒二节和倒二芽中CTK含量均显著上升, 且随着分蘖的生长, CTK含量持续上升, CTK含量的降低不利于分蘖芽萌发。这表明CTK含量的变化在水稻分蘖芽休眠与萌发转换过程中起重要作用, 较高的CTK含量有利于分蘖萌发, 而较低的CTK含量则会导致分蘖休眠。IAA在顶端合成并通过极性运输运至基部抑制侧芽生长[ 6], 消除或抑制IAA的活性可以打破侧芽的休眠[ 22], 尚玉磊等[ 25]研究发现, 在稻麦的分蘖发生过程中, IAA含量显著下降并维持在较低值; 李经勇等[ 12]试验结
果表明, 水稻叶片和穗中IAA含量的降低, 有利于再生芽萌发; 另有研究结果显示, IAA是在木质部和束间厚壁组织中起作用, 进而间接影响分枝生长。本试验结果显示, 留穗和IAA处理的分蘖芽中IAA含量较高, 分蘖芽处于休眠状态, 分蘖芽从休眠向萌发转换过程中, 倒二节中IAA含量显著降低, 试验结果还显示IAA含量的变化先于分蘖芽伸长, 说明节内IAA含量降低可能是分蘖芽萌发的前提条件, 而较高的IAA含量诱导分蘖芽处于休眠状态, 这与前人研究结果一致。倒二芽中IAA含量在分蘖休眠与萌发转换的过程中无明显变化, 说明IAA并没有直接作用在水稻的分蘖芽上, 分蘖芽中IAA绝对含量不是影响分蘖发生的关键因素, IAA对分蘖芽生长的抑制作用是间接的, 推测IAA是通过控制CTK在倒二节中运输的通道, 进而影响分蘖芽中CTK含量来调控分蘖发生。前人更多的研究表明, 几种激素的平衡互作在分枝生长的调控过程中起重要作用[ 6, 13, 15, 26, 27]。李春喜等[ 15]发现, 小麦、水稻和玉米分蘖节中ZR/IAA值升高时, 作物分蘖能力强。冬小麦衰亡分蘖中IAA/ZR值在分蘖开始衰亡之前就开始降低, 未衰亡分蘖中IAA/ZR则维持稳定[ 13, 14]。较高的IAA/ZR值抑制玉米分蘖发生, 较低的IAA/ZR值则促进玉米分蘖发生[ 24]。刘杨[ 16]也发现随着IAA/ZR值升高, 不利于分蘖芽继续生长。本试验表明, 休眠材料CTK/IAA值维持在较低的水平, 倒二节和倒二芽中CTK/IAA值分别为1.5~1.7和2.5~2.8, 在分蘖芽从休眠向萌发转换的过程中, 分蘖节、芽中CTK/IAA值均显著升高, 说明CTK和IAA的相对含量是调控侧芽生长的关键因素, 提高CTK/IAA值可以促进分蘖芽萌发, 且CTK含量的增幅显著高于IAA含量的变化, 推测IAA是通过调控CTK的含量进而调控分蘖生长。可以推断分蘖芽休眠与萌发相互转换时, 倒二节中CTK/IAA值在2.6~2.8, 倒二芽中CTK/IAA值在4.8~5.0, 为分蘖芽休眠与萌发转换的临界值。
水稻分蘖芽的休眠与萌发受CTK和IAA共同调控, 外施植物激素可以改变内源激素含量及其平衡, 进而影响分蘖芽萌发。在分蘖芽从休眠向萌发转换的过程中, 节、芽内的CTK含量均显著上升, 节内IAA含量显著下降, 而芽中IAA含量无显著变化, 节、芽中CTK/IAA值均显著提高, 节中CTK/IAA值在2.6~2.8, 芽中CTK/IAA值在4.8~5.0, 可作为分蘖芽休眠与萌发转换的临界指标。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|