
{kind=link}
{kind=link}
施氮期对小麦茎秆木质素合成的影响及其抗倒伏生理机制
[卢昆丽, 尹燕枰, 王振林*  , 李勇, 彭佃亮, 杨卫兵, 崔正勇, 杨东清, 江文文]
, 李勇, 彭佃亮, 杨卫兵, 崔正勇, 杨东清, 江文文]
, 李勇, 彭佃亮, 杨卫兵, 崔正勇, 杨东清, 江文文]
|
|


为明确氮肥施用模式对小麦茎秆木质素合成与积累及抗倒伏能力的影响, 2011—2012和2012—2013生长季, 选用济麦22 (抗倒伏)和山农16 (不抗倒伏)品种, 分析不同生育期追施氮肥处理的茎秆木质素含量和木质素合成相关酶活性, 及其与抗倒伏指数和产量的关系。各处理总施氮量为240 kg hm-2, 其中80 kg hm-2播前基施, 其余氮肥按4种模式追施, 分别是起身期和孕穗期各一半(N1)、拔节期全施(N2)、拔节期和开花期各一半(N3)和孕穗期全施(N4)。与N2和N3处理相比, N1和N4处理的茎秆木质素含量高, 茎秆抗折力大, 茎秆抗倒能力强。各处理对木质素合成关键酶活性有显著影响, 其中N1的苯丙氨酸解氨酶(PAL)、酪氨酸解氨酶(TAL)和过氧化物酶(POD)活性较高; N2处理第2节间形成后0~7 d时PAL、TAL和POD活性显著高于其他处理, 但第2节间形成后21 d时迅速下降; N3处理第2节间形成后0~21 d时PAL、TAL和POD活性低, 但开花期(21 d)追氮延缓了3种酶活性的降低; N4处理显著提高了第2节间形成14 d后的酶活性。在乳熟期和蜡熟期, 两品种的基部茎秆抗折力和抗倒伏指数均以N1和N4处理显著优于其他处理, 并且N1和N4处理的籽粒产量也显著高于其他处理。试验结果表明, 孕穗期追肥的N1和N4处理有助于提高茎秆中PAL、TAL、POD活性及木质素含量, 提高小麦茎秆的机械强度及抗倒伏能力, 降低小麦倒伏面积和倒伏程度。
, LI Yong, PENG Dian-Liang, YANG Wei-Bing, CUI Zheng-Yong, YANG Dong-Qing, JIANG Wen-Wen
For understanding the effects of nitrogen management on lignin synthesis in wheat stem and plant lodging resistance, a field experiment was conducted, using the lodging resistant cultivar Jimai 22 and the lodging sensitive cultivar Shannong 16, in 2011-2012 and 2012-2013 growing seasons. The lignin accumulation in stem, activities of enzymes related to lignin synthesis, lodging resistance index, and grain yield were evaluated under four nitrogen treatments. All treatments received 240 kg ha-1 nitrogen during the whole growth period, in which 80 kg ha-1 was applied before sowing. The remaining nitrogen was applied at pseudo stem erection and booting stages (1:1) for N1, at jointing stage for N2, at jointing and anthesis stages (1:1) for N3, and at booting stage for N4 treatment. Compared with N2 and N3, N1 and N4 were superior in lignin content, stem mechanical strength, and lodging resistance. Nitrogen fertilization timing had significant effects on activities of phenylalanine ammonia-lyase (PAL), tyrosine ammonia-lyase (TAL), and peroxidase (POD). Under N1 condition, the three key enzymes were in high activity. PAL, TAL and POD activities were higher in N2 treatment than in other treatments at 0-7 d after emergence of the second internode (ESI), and dropped rapidly at 21 days after ESI. Under N3 condition, the PAL, TAL and POD activities were low at 0-21 days after ESI, and the activity decline was alleviated by the nitrogen supplement at anthesis stage (21 days after ESI). Under N4 condition, the activities of these enzymes were increased after 14 days of ESI. At milk and dough stages, both cultivars had higher snapping resistance of the basal stem in N1 and N4 than in N2 and N3 treatments. Besides, the lodging resistance index and grain yield were also significantly higher in N1 and N4 than in N2 and N3. These results indicated that nitrogen applied at booting stage (N1 and N4 treatments) is propitious to increase PAL, TAL, POD activities and lignin content in stem, leading to the improved stem mechanical strength and decreased lodging area and lodging degree of wheat.
小麦倒伏常发生于灌浆中后期, 是高产、稳产、优质生产的主要限制因素之一[ 1, 2]。倒伏发生的时期越早, 倒伏程度越高, 造成的产量损失就越大[ 3, 4]。据调查, 近10年我国黄淮冬麦区高产麦田每年约有5%~10%的面积发生倒伏, 重的年份倒伏面积达10%~20%以上, 导致减产约20%~30%, 严重者减产50%, 甚至绝收。小麦植株抗倒能力与株高和茎秆机械强度密切相关, 适度降低株高是提高抗倒性的有效措施之一[ 5, 6], 但植株生物量是高产的物质基础, 单纯降低株高难以同步实现高产和抗倒伏, 而增强茎秆机械强度、提高茎秆质量成为茎秆抗倒的关键因素[ 7, 8, 9]。木质素是决定细胞壁强度和茎秆机械强度的主要次生代谢物之一[ 10, 11]。小麦茎秆中缺乏木质素易发生倒伏[ 12]。对不同抗倒性品种的研究表明, 茎秆木质素含量可作为小麦品种抗倒性评价的一个重要指标[ 13, 14]。倒伏多发生在茎秆基部第1、第2节间, 因此, 茎秆基部节间充实度高、茎壁厚、节间短则抗倒性强[ 15, 16]。
施用氮肥是小麦增产的主要栽培措施之一, 但不合理施氮容易引起倒伏, 主要原因是基肥过量或拔节前追肥多, 促使小麦大量分蘖, 群体偏大使基部节间拉长变细, 茎秆木质化受阻而发育质量变差, 在风雨天气极易发生倒伏[ 3, 17]。前氮后移施肥技术可显著减少无效分蘖, 促使个体生长健壮, 增强植株抗倒能力[ 18]。随施肥期的推迟, 穗下节间与基部节间的比值增大, 株型结构合理, 小麦植株倒伏率降低[ 3]。适当降低氮肥用量, 能增加基部节间木质素含量, 提高茎秆的抗倒性能[ 2], 而高施氮量处理则降低茎秆木质素合成相关酶的活性和木质素含量, 茎秆抗倒伏能力降低[ 19]。目前, 已有研究报道了氮肥水平对小麦茎秆木质素合成与积累规律的影响, 关于施氮期对茎秆木质素的调控作用尚无深入研究。本研究以抗倒性不同的2个小麦品种为材料, 研究施氮期对小麦茎秆木质素合成与积累规律的影响, 以期在施氮量一定的前提下, 提高氮肥利用率, 增强茎秆质量, 为提高小麦抗倒伏能力提供理论依据与技术支持。
2011--2012和2012--2013年连续2个生长季, 选用抗倒品种济麦22和非抗倒品种山农16, 在山东农业大学泰安农学试验农场进行田间试验。试验田土壤为棕壤土, 试验地耕层(0~20 cm)土壤含有机质12.3 g kg-1、全氮0.91 g kg-1、碱解氮87.2 mg kg-1、速效磷78.6 mg kg-1 和速效钾57.5 mg kg-1。小麦全生育期施纯N 240 kg hm-2、P2O5 90 kg hm-2、K2O 100 kg hm-2, 氮肥为尿素, 磷肥为过磷酸钙, 钾肥为氯化钾, 全部磷、钾肥和氮肥三分之一(80 kg hm-2)作基肥, 均匀撒施于各小区后翻耕入土, 剩余氮肥开沟追施, 设4个追施模式, 即起身期和孕穗肥1∶1 (N1)、拔节期全施(N2)、拔节期和开花期1∶1 (N3)、孕穗期全施(N4)。小区面积3 m × 3 m=9 m2, 3次重复随机区组排列。于2011年10月10日和2012年10月7日人工条播, 基本苗为2.1 × 106株 hm-2, 行距25 cm, 于2012年6月10日和2013年6月14日收获。其他田间管理同小麦生产田。
1.2.1 茎秆抗折力和抗倒伏指数 茎秆抗倒伏指数可作为衡量和评价小麦茎秆抗倒伏能力的一个综合指标, 抗倒伏指数越高, 作物茎秆抗倒伏能力越强[ 9]。按Zadoks的0~9级生育期系统[ 20], 分别于开花期、乳熟期和蜡熟期取长相一致植株20株, 参照李金才等[ 8]的方法测定茎秆抗折力和抗倒伏指数。取基部第2节间, 剥除叶鞘, 置两端于高50 cm、间隔5 cm的支撑木架凹槽内, 将YYD-1型茎秆强度测定仪(浙江托普仪器有限公司, 杭州)置茎秆中部并匀速下压, 茎秆折断时屏幕上显示的峰值即为茎秆抗折力(N)。抗倒伏指数 = 抗折力/植株重心高度。茎秆重心高度为茎秆基部至该茎(带穗、叶和鞘)平衡支点的距离(cm)。
1.2.2 木质素含量 参照Bhaskara Reddy[ 21]的方法。取茎秆加液氮于研钵中磨碎, 称取约0.1 g装入10 mL离心管; 加95%乙醇提取叶绿素, 5000 × g离心10 min后弃上清液; 沉淀加等量正己烷∶乙醇(2∶1)溶液, 5000 × g离心10 min弃上清液。沉淀经烘干再用25%溴乙酰冰醋酸2.5 mL溶解, 70℃水浴, 并加塞保温30 min, 然后在冷水中快速冷却, 加2 mol L-1 NaOH 0.9 mL终止反应; 再加 7.5 mol L-1盐酸羟胺0.1 mL和冰乙酸4 mL, 混匀后5000× g离心5 min, 吸取上清液0.1 mL, 加冰乙酸3.9 mL稀释, 测定波长280 nm下的OD值。以每克鲜样在280 nm处的吸光值表示木质素含量。
1.2.3 酶活性 分别于基部第2节间形成后0、7、14、21 d (开花期)、28、35 (乳熟期)和42 d, 从每小区取典型植株20株, 将去除茎鞘的基部第2节间放入液氮速冻, -40℃冰箱保存, 按已报道的方法分别测定苯丙氨酸解氨酶(PAL)[ 22]、酪氨酸解氨酶(TAL)[ 23]和过氧化物酶(POD)[ 24]的活性。
1.2.4 表观倒伏率 收获前测量实际倒伏面积, 表观倒伏率(%) = 倒伏面积/总面积 × 100。
两年度结果趋势一致, 本文采用2012—2013年
木质素含量及PAL、TAL和POD活性, 其他指标为两年度数据。用Microsoft Excel 2003整理数据并作图, 用DPS 7.05 (Date Processing System)和SPSS 18.0软件统计分析, 采用Duncan’s新复极差(SSR)法检验差异显著性。
山农16各处理在蜡熟期均发生不同程度倒伏, 倒伏率表现为N2>N3>N4>N1。表明施氮量相同情况下, 起身期或孕穗期追氮显著降低小麦实际倒伏率, 而拔节期或开花期追氮显著增加小麦实际倒伏率; 济麦22仅N2处理在2011—2012年的蜡熟末期倒伏, 其余3个处理未倒伏(表1)。表明拔节期过多追施氮肥易引发小麦倒伏。倒伏分级越大, 小麦实际倒伏率越高。
| 表1 山农16和济麦22的倒伏发生期、倒伏程度和倒伏率 Table 1 Lodging occurrence stage, degree, and percentage in Shannong 16 and Jimai 22 |
开花期至蜡熟期, 两品种小麦茎秆机械强度先增高后降低, 乳熟期达最大值。同一生育时期, 济麦22的茎秆抗折力显著高于山农16。2012—2013年度开花期, 两品种各处理的茎秆抗折力均呈N2>N1>N3>N4。与开花期相比, 山农16乳熟期N1、N2、N3和N4处理的茎秆抗折力分别高26.2%、9.5%、18.5%和41.9%, 而济麦22高32.2%、13.3%、25.6%和39.7% (表2)。表明济麦22具有较强的自身调节能力, 抗倒伏能力更强, 与N2、N3处理相比, N1、N4处理的茎秆抗折力增加显著, 即乳熟期茎秆机械强度较大。拔节期追肥量过多或追肥时期较晚, 均不利于茎秆强度的提高, 但孕穗期追氮使乳熟期茎秆机械强度提高。
| 表2 不同生育期山农16和济麦22的基部茎秆抗折力 Table 2 Snapping resistance of basal stem of Shannong 16 and Jimai 22 at different growth stages |
同一生育期, 济麦22的茎秆抗倒伏指数显著高于山农16。仅山农16 N2处理的茎秆抗倒伏指数在乳熟期显著降低, 其余各处理的茎秆抗倒伏指数均呈先上升后下降的趋势, 蜡熟期降至最低值。2012—2013年度乳熟期山农16的N1、N3和N4处理的茎秆抗倒伏指数分别比开花期提高3.4%、1.5%和11.9%, N2处理抗倒伏指数降低5.5%; 而济麦22 4个处理分别提高13.5%、1.7%、13.6%和19.2% (表3)。表明拔节期追施过量氮肥不利于茎秆抗倒能力的提高, 而孕穗期追施氮肥可提高乳熟期茎秆的机械强度, 及抗倒伏能力。
两品种茎秆木质素含量呈S曲线变化, 且济麦22节间木质素含量显著高于山农16。成熟期N1处理茎秆中木质素含量显著高于N2、N3和N4处理(除济麦22的N4与N1无显著差异外)。茎秆木质素积累量, 第2节间形成后0~14 d, N2显著高于其他处理, 但14 d后差异不显著; 第2节间形成后28 d内N3低于N2, 但开花期追施氮肥对积累速度的降低起到缓冲作用, 使N3成熟期茎秆中的木质素含量显著高于N2; N4比其他处理延迟了木质素的快速积累, 但提高了第2节间形成14 d之后的积累速度, 延长了木质素的快速积累期, 使成熟期木质素含量显著提高(图1)。表明追肥时期后移至孕穗期有利于第2节间形成14 d之后茎秆中木质素的积累, 有利于增强茎秆的抗倒伏能力, 而拔节期追肥量过多或开花期追肥不利于成熟期茎秆中木质素含量的提高。
| 表3 不同生育期山农16和济麦22的茎秆抗倒伏指数 Table 3 Stem lodging resistance index of Shannong 16 and Jimai 22 at different growth stages |
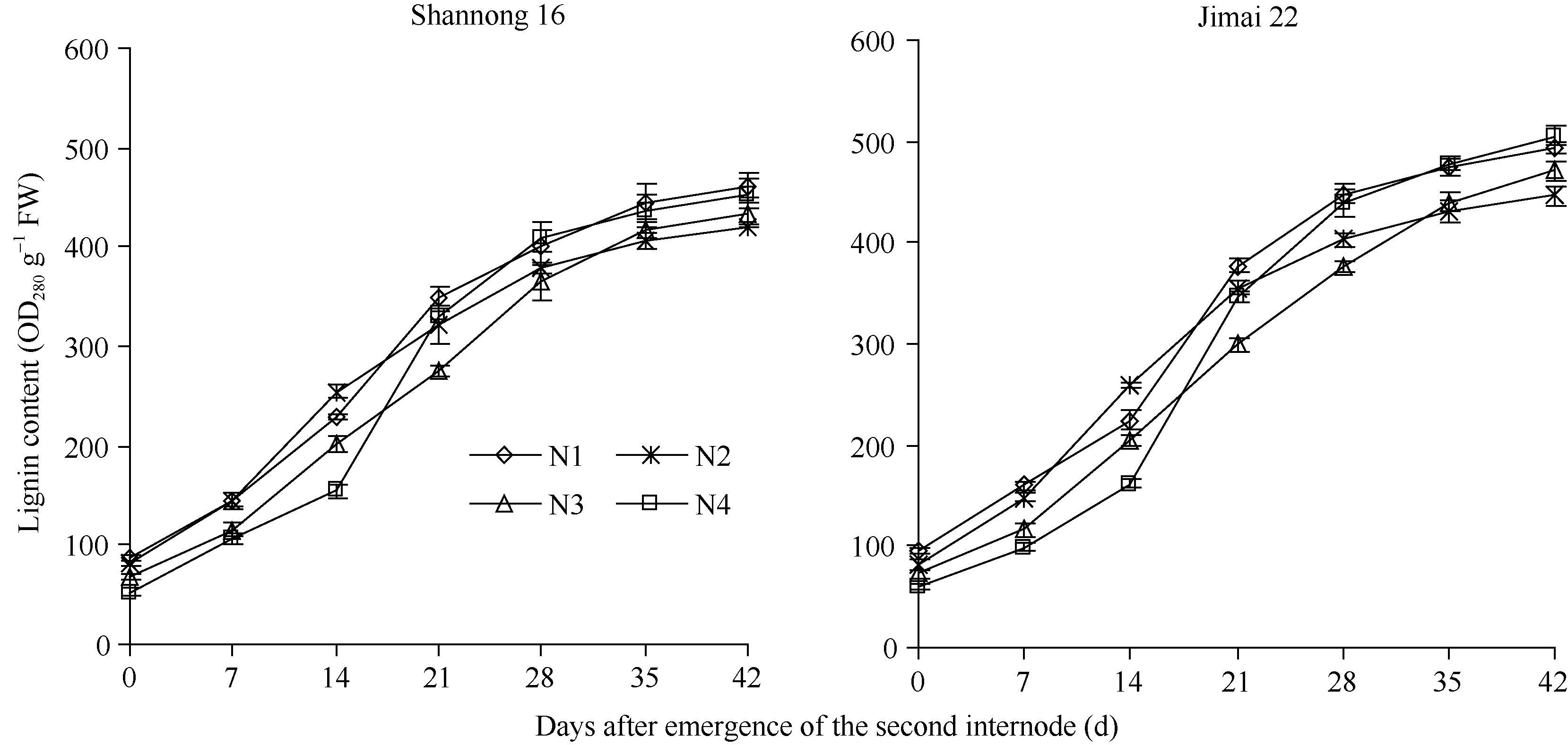 | 图1 不同施氮处理小麦茎秆木质素含量的变化(2012-2013)Fig. 1 Changes of lignin content of wheat stem in different nitrogen treatments (2012-2013) |
随茎秆基部第2节间生长发育进程, 各处理PAL活性呈下降趋势, 第2节间形成后0~21 d是PAL的活跃期, 济麦22茎秆的PAL活性显著高于山农16 (图2-A, B)。第2节间形成后14~21 d, N1的PAL活性显著高于其他处理; 第2节间形成后0~7 d, N2的PAL活性明显高于其他处理, 此后呈缓慢下降再迅速下降; 与N2处理相比, N3的PAL活性降低, 但开花期施开花肥对第2节间形成后35~42 d PAL活性的降低有缓冲作用; 第2节间形成后0~14 d, 两品种N4处理的PAL活性显著低于其他处理, 此后PAL活性缓慢下降, 说明酶活性高活跃期被延长。PAL活性的高活跃期与木质素的快速积累期同步, 追氮时期后移有利于提高第2节间形成14 d后茎秆中PAL的活性。
两品种各处理茎秆基部第2节间TAL活性均呈单峰曲线变化, 第2节间形成后14 d达活性高峰, 而后迅速下降。济麦22第2节间形成后0~7 d的TAL活性上升迅速, 且高峰显著高于山农16。第2节间形成后0~14 d, 两品种N1的TAL活性显著高于N4, N2的TAL活性显著高于N3, N3的TAL活性显著高于N4 (图2-C, D)。
两品种各处理茎秆基部第2节间POD活性亦呈先上升再降低的趋势, 第2节间形成后14 d达活性高峰, 且N1>N2>N4>N3。济麦22的POD活性显著高于山农16。第2节间形成后0~7 d, 山农16各氮肥处理POD活性为N2>N1>N3>N4, 而济麦22各处理POD活性为N1>N2>N3>N4。第2节间形成后14~21 d, 两品种POD活性为N1>N2>N3 (图2-E, F)。
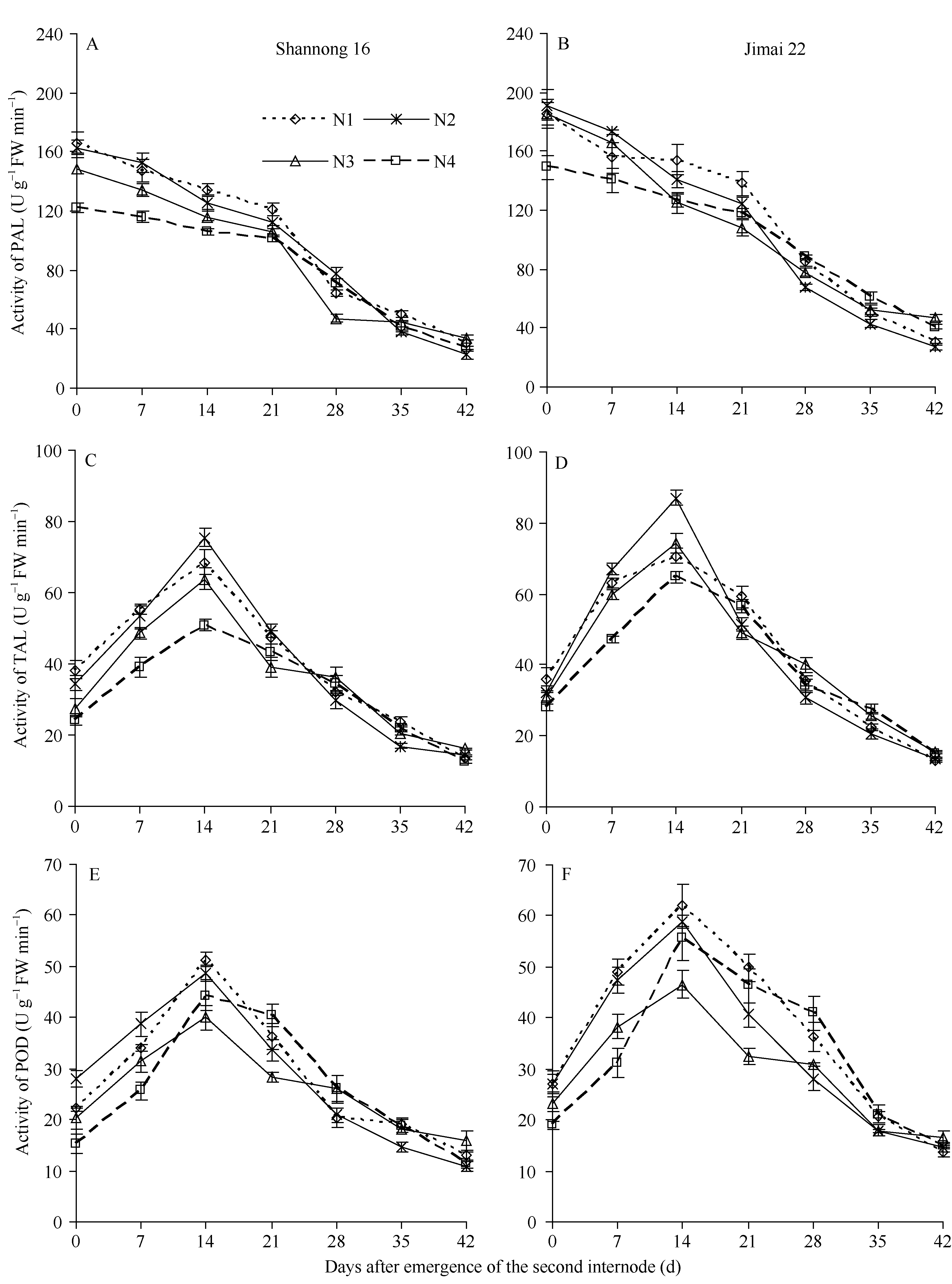 | 图2 不同施氮处理对苯丙氨酸解氨酶(PAL)、酪氨酸解氨酶(TAL)和过氧化物酶(POD)活性的影响(2012-2013)Fig. 2 Effects of different nitrogen treatments on activities of phenylalanine ammonia-lyase (PAL), tyrosine ammonia-lyase (TAL), and peroxidase (POD) (2012-2013) |
山农16的籽粒产量呈N1>N4>N2>N3的规律; 而济麦22的籽粒产量, 以N1处理最高, N3处理最低, N2和N4处理相近, 差异不显著。两品种穗数均有N1>N2>N3>N4的趋势, 但济麦22处理间的差异明显大于山农16。总体来看, 千粒重以N4处理最高(济麦22 2011—2012年度N1较N4略高, 但差异不显著), 且显著优于其他处理; 穗粒数以N1处理显著高于N3处理; N4处理有较高的单穗结实粒数和千粒重, 但由于受成穗数的限制, N4处理产量低于N1处理(表4)。可见, 过晚追肥或重施孕穗肥均不利于高产。
| 表4 不同施氮处理的籽粒产量及产量构成因素 Table 4 Grain yield and yield components in different nitrogen treatments |
茎秆基部第2节间木质素含量与茎秆抗折力、抗倒伏指数均呈极显著正相关, 表明茎秆木质素含量高, 小麦茎秆抗倒伏能力强; 同时木质素含量与3种木质素合成相关酶的活性也显著正相关, 说明提高茎秆中PAL、TAL和POD活性可增加木质素含量(表5)。
| 表5 木质素含量及相关酶活性与抗倒伏能力相关分析(2012-2013) Table 5 Lignin content and related enzyme activity and their relationships with lodging resistance (2012-2013) |
为明确影响茎秆基部第2节间木质素含量的关键酶及其差别, 建立了茎秆基部第2节间中PAL ( x1)、TAL ( x2)和POD ( x3)活性对木质素含量( y)的回归方程 y= 1.569 x1 - 1.635 x2 + 8.962 x3 + 237.670 ( R2 = 0.870)。可见, 对木质素含量的影响表现为POD>TAL>PAL。
木质素是植物苯丙烷代谢途径的主要终产物之一, 是维管植物细胞壁的重要组成成分。木质素与半纤维素以共价键结合, 具有加固细胞壁机械强度和增强植物抗倒伏性的生物学功能[ 25]。研究表明, 抗倒伏能力越强的品种, 其木质素含量越高[ 12, 14, 26], 但也有学者研究发现, 倒伏品种中茎秆木质素占干物质的百分率总是高于不倒伏品种[ 27, 28], 或者不同抗倒性品种的茎秆木质素含量无明显差异[ 29]。本研究表明, 木质素含量与茎秆抗倒伏指数呈极显著正相关。山农16的倒伏率显著高于济麦22, 且倒伏面积大, 茎秆中木质素的含量也较低。本研究支持抗倒性品种茎秆木质素含量高的论点。
PAL、TAL和POD是禾本科植物木质素合成的关键酶。相关分析表明, 小麦茎秆木质素含量与PAL和POD活性均呈极显著正相关, 与TAL活性呈显著正相关(表5), 表明较高的PAL、TAL和POD活性有利于木质素的合成与积累。PAL催化L-苯丙氨酸脱氨生成反式肉桂酸, 是苯丙烷类代谢途径的关键酶和限速酶, 其表达及其丰度直接影响木质素生物合成的整个过程[ 30]。Sewalt等[ 31]报道, PAL活性下降时, 植物体中的木质素含量下降2倍。本研究中, PAL的高活性期与木质素含量的快速增长期(快速拔节期)基本同步, PAL活性降低, 木质素的增加量也降低。TAL催化酪氨酸生成香豆酸, 仅存在于禾本科植物中[ 32]。我们观察到TAL活性在小麦生长发育过程中呈单峰曲线变化, 第2节间形成后14 d达到峰值, 此时正值木质素含量的快速增长期; 抗倒能力强的处理, 其植株TAL活性高。这与陈晓光等[ 14]的研究结果一致。TAL活性高为小麦茎秆基部第2节间的物质充实提供了保障。茎秆的PAL、TAL活性能在一定程度上反映茎秆的抗倒伏能力[ 14]。POD是植物活性氧代谢中活性氧清除系统的一种重要酶类, 同时也是木质素生物合成中最后一步的关键酶。POD在H2O2参与下催化木质素单体聚合成木质素, 提高组织木质化程度, 从而增强茎秆的抗倒伏能力[ 32, 33]。与薛应钰等[ 34]的试验结果一致, 本研究中POD活性与木质素含量之间也呈极显著正相关。茎秆生长发育过程中, 除个别时期外(0 d), 济麦22中PAL、TAL和POD 酶活性均高于山农16。说明无论是木质素合成特异途径, 还是木质素单体合成阶段, 济麦22茎秆中酶催化木质素相关反应的能力总体高于山农16。3种酶中POD对茎秆基部第2节间木质素合成的影响最大, 其活性高是小麦抗倒伏能力强的原因之一。
合理的氮肥运筹模式能显著增加茎秆木质素含量, 改善其抗倒伏性[ 9]。适当减少氮肥用量, 能够增加基部节间木质素含量, 提高茎秆的抗倒性能[ 2]。高施氮量处理则降低了茎秆木质素合成相关酶的活性和木质素含量, 茎秆抗倒伏能力降低[ 19]。地力条件较好的情况下, 实施氮肥后移技术, 增加小麦生育中后期的追肥比例, 有利于优化群体结构, 增强抗倒伏能力[ 35]。徐本生等[ 17]发现拔节、孕穗期施用等量的氮肥, 倒伏程度以孕穗期施肥的较轻, 推测其原因是此时茎节细胞的分生组织活动能力减弱, 机械组织已进入稳定状态。随施肥期的推迟, 尤其推迟至药隔期和四分子期, 穗下节间与基部节间的比值增大, 株型结构合理, 小麦植株倒伏率降低, 产量高[ 3]。在黄淮麦区小麦基施氮肥过多, 造成冬前小麦叶蘖生长旺盛, 甚至徒长, 分蘖多, 群体过大, 传统的春季麦田采用与返青水“一炮轰”的追肥法, 氮肥施用偏早, 增加了春季无效分蘖, 刺激小麦底部茎节的生长, 使基部茎节细长、脆弱, 易倒伏。因此, 需要通过合理氮肥运筹等措施提高小麦茎秆强度, 以增强其抗倒性。
4种氮肥追施模式下, N1处理(起身期和孕穗期各一半)中持续的氮肥供应可以满足植株对氮的需求, 整个生长发育过程中PAL、TAL和POD活性及木质素含量较其他处理处于较高的水平; N2处理拔节期一次性追施氮肥, 在第2节间形成初期氮肥供应充足, 其PAL、TAL和POD活性及木质素积累速度显著高于其他处理, 但第2节间形成21 d后, 可能由于开花后营养器官的物质大量运往生殖器官, 用于籽粒的形成和灌浆, 又无外源的氮肥补充, 茎秆中氮含量低, PAL、TAL和POD活性迅速下降, 木质素积累量降低, 茎秆抗倒伏能力下降; N3处理(拔节期和开花期各一半)因拔节期氮肥施用量降低, PAL、TAL和POD活性及木质素积累量明显降低, 而开花期追氮后充足的氮肥供应对第2节间形成28 d后3种酶活性及木质素积累速度的降低起到缓冲作用, 成熟期茎秆中木质素含量较低, 茎秆抗倒伏能力较差; N4处理全部追施氮肥后移至孕穗期, 虽然第2节间形成初期PAL、TAL和POD酶活性低且木质素含量积累慢, 但显著提高了第2节间形成14 d后的酶活性及木质素的积累量, 提高了茎秆机械强度和抗倒伏指数。说明孕穗期追肥的N1和N4模式有助于提高PAL、TAL及POD活性, 有利于小麦茎秆中木质素的积累, 从而提高茎秆的抗倒伏能力。
在总施氮量240 kg hm-2、基施80 kg hm-2条件下, 起身期和孕穗期按1∶1追施其余氮肥有助于提高茎秆中木质素含量和茎秆抗倒伏指数。氮肥施用模式通过调控茎秆PAL、TAL和POD活性及木质素含量, 影响小麦的抗倒伏能力。在本试验区的小麦生产中, 建议适当减少氮肥基施量, 中、后期追施各占总氮量的30%左右, 可以满足小麦生长氮需求, 同时有利于提高茎秆中的木质素含量, 增强抗倒能力, 为高产打下基础。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|