

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
膜下滴灌对棉花生育后期叶片与苞叶光合特性的影响
[张超 , 占东霞, 张亚黎, 罗宏海, 勾玲, 张旺锋
, 占东霞, 张亚黎, 罗宏海, 勾玲, 张旺锋* ]
, 占东霞, 张亚黎, 罗宏海, 勾玲, 张旺锋]
|
|


第一作者联系方式: E-mail:dhtzcwlp2004@126.com
在新疆气候生态条件下, 选用新陆早33号和新陆早46号棉花品种, 设置2个膜下滴灌量处理, 测定棉花叶片和苞叶的光响应曲线、CO2响应曲线、荧光参数等相关光合生理指标, 探讨生育后期棉花叶片与苞叶光合能力的差异及对滴灌量的响应。结果表明, 棉花生育期间, 高温强光下苞叶最大羧化速率与最大电子传递速率的比值( Jmax/ Vc,max)较高, 气孔导度( Gs)较低。棉花生长发育后期, 苞叶面积、含水量、叶绿素含量、净光合速率( Pn)、光系统II实际光化学效率( ΦPSII)、Rubisco含量显著低于叶片, 但随着生育进程降幅显著低于叶片。与常规滴灌处理相比, 节水滴灌处理棉花叶片含水量降幅显著高于苞叶; 叶片叶绿素含量、 Pn和Rubisco含量显著降低, 而苞叶的变化不显著。棉花生育后期苞叶面积、含水量、叶绿素含量、 Pn、 ΦPSII、Rubisco含量的稳定性高于叶片, 表明生育后期叶片衰老较快, 但苞叶仍能保持较稳定的光合能力, 对光合物质的贡献逐渐增大, 常规滴灌下达7.22%~8.83%, 节水滴灌下达10.24%~12.53%。
The majority of cotton ( Gossypium hirsutumL.) photosynthetic products comes from leaves, but non-foliar green organs of cotton such as bracts also contain chlorophyll and may contribute to the yield. We selected Xinluzao 33 and Xinluzao 46 with two irrigation treatments (normal drip irrigation and water-saving drip irrigation) to determine physiological indices including response curves of net photosynthetic rate ( Pn), photosynthetically active radiation (PAR), intercellular CO2 concentration ( Ci), fluorescence parameters, and water content of cotton bracts and leaves and to explore the difference of photosynthetic capacity between bracts and leaves under drip irrigation conditions. The results showed that bracts showed greater RuBP regeneration/RuBP carboxylation capacity ratio ( Jmax/ Vc,max) and lower stomatal conductance under high irradiance and temperature conditions. Photosynthetic area per plant, chlorophyll content, water content, Pn, the actual photochemical efficiency of PSII ( ΦPSII), and Rubisco content were less in bracts than in leaves, but the decline of the indices showed a significantly faster in leaves than in bracts at the late growth stage. Compared with normal drip irrigation, the decreased degree of water content under water-saving drip irrigation was significantly higher in leaves than in bracts of cotton plants, and chlorophyll content, Pn, and Rubisco content decreased in leaves under water-saving drip irrigation, but there was no significant changed in bracts. The stabilities of bracts area, water content, chlorophyll content, Pn, ΦPSII, rubisco content in bracts were higher than those in leaves. It indicated the photosynthetic activity of leaves decreased rapidly. The bracts contributed approximate 7.22%-8.83% of the total photosynthate at the late growth stage under normal drip irrigation, and 10.24%-12.53% under water-saving drip irrigation. We concluded that bracts can increase the photosynthate of plant at the late growth stage under water-saving drip irrigation.
叶片是植物主要的光合器官, 叶片早衰导致叶绿素含量降低, 叶绿体的数目减少、结构和功能发生改变, 降低光合生产能力[1, 2, 3]。叶片衰老过程中, 所捕获的激发能超过其碳同化的利用能力时, 过多的激发能导致光系统II (PSII)光碳失衡, 从而降低作物的光合生产力[4, 5]; 有关分离叶绿体或类囊体膜的研究发现, 叶片衰老诱导光系统II (PSII)和光系统I (PSI)光化学活性的降低, 并且在大多数情况下PSII比PSI更容易衰老, PSII活性的下降导致光合能力降低[6, 7, 8]。类囊体膜蛋白的降解导致光合酶活性下降, 进而影响光合活性[9, 10], 其中Rubisco酶活性的降低是导致光合速率下降最主要的原因[11, 12]。
前人研究表明, 作物的非叶绿色器官, 如小麦 (Triticum aestivum L.)的穗[13, 14]、棉花(Gossypium hirsutum L.)的苞叶[15]、大豆(Glycine max)的豆荚[16]、玉米(Zea mays L.)的苞叶[17]等均含有叶绿素, 具有光合作用的潜力, 对产量形成有一定的贡献; 作物非叶绿色器官具有较强的光合抗逆性, 充分发挥非叶绿色器官的光合耐逆功能, 对提高整体光合能力具有重要意义[18, 19, 20]。棉花叶片是碳同化的主要器官[21], 但棉花苞叶、棉铃、茎秆[15]等非叶绿色器官具有光合作用。研究表明, 棉花叶片光合产物对棉铃发育的贡献是有限的, 尤其在生育后期叶片的贡献显得尤为不足[22, 23, 24, 25]。水分胁迫抑制了叶片面积的伸展[26], 降低叶片光能转化率和光化学活性[27], 增加活性氧[28], 加重了叶片的早衰。在光合“ 源” 受限制环境下, 棉花非叶绿色器官的光合作用对产量形成的作用更为突出。棉花苞叶包裹在棉铃外, 在生理位置上紧挨着产量器官棉铃, 具有较强的光合能力, 在产量形成中具有重要作用[24]。因此, 研究节水条件下棉花苞叶的光合潜力, 探讨增强棉株整体的光合抗逆能力, 减轻土壤水分亏缺对棉株产量的不利影响, 是实现棉花节水高产栽培迫切需要研究的重要内容。本研究针对新疆典型大陆性干旱气候, 比较不同水分条件下棉花生育后期叶片与苞叶光合能力的差异及对生育后期干旱逆境的适应机制, 探究苞叶光合作用对棉花产量的贡献, 为新疆棉花高产品种的选育及节水抗逆栽培提供理论依据。
2012— 2013年在石河子大学农学试验站(45.32° N, 86.05° E)种植新疆棉区主栽品种新陆早33号和新陆早46号, 2个品种生育期为124~125 d。设置2个滴灌量处理, 常规滴灌量每次滴水至田间持水量的80%; 节水滴灌量每次滴水量为常规滴灌量的60%; 采用裂区试验设计, 主区为水分处理, 副区为品种, 重复3次; 小区面积44 m2。两年试验分别于4月25日和4月23日播种, 于棉花开花期进行滴灌水分处理。田间种植方式、管道铺设方法及田间管理同一般大田膜下滴灌棉花。
于棉花盛花期植株打顶后, 在各小区分别选取长势均匀、有代表性的棉株, 对主茎倒二叶挂牌标记, 记录同时开花的倒二果枝第一果节花蕾。在棉花开花期水分处理后开始定期取样测定, 一般选择灌水后第4天测定所标记叶片及棉铃苞叶的气体交换参数、叶绿素荧光参数、环境因子及棉株各器官含水量的变化, 共取样5次, 取样时间分别在出苗后85、95、105、115和125 d前后, 即对应棉花的盛花期、盛铃前期、盛铃中期、盛铃后期、吐絮初期。
在各小区分别选取长势均匀有代表性的棉株3株, 用LI-3000 (LI-COR, Lincoln, USA)叶面积仪测定单株叶片和苞叶面积。选取所标记的叶片与相邻棉铃的苞叶, 采用Lichtenthaler[29]的方法测定叶绿素含量, 烘干法测定植株叶片与苞叶含水量。
选择晴朗无风天气, 在北京时间11:30至13:30用LI-6400便携式光合测定系统(LI-COR, Lincoln, USA)和6400-02LED红蓝光源叶室(LI-COR, Lincoln, USA), 测定所标记叶片及苞叶的光响应曲线(Pn-PAR)和CO2响应曲线(Pn-Ci)。在测定光响应曲线时, 采用开放式气路, 气温为30~35℃, 空气相对湿度50%~70%, 光强分别为2000、1800、1500、1200、1000、800、500、200、50和0 μ mol m-2 s-1。测定CO2浓度响应曲线时, 气温为30~35℃, 空气相对湿度50%~70%, 光强为2000 μ mol m-2 s-1, CO2浓度分别为2000、1000、500、400、300、200、100、50和0 μ mol m-2 s-1。采用轮回方法测定不同处理。
参考Schreiber等[30, 31]的方法, 在北京时间11:30至13:30, 采用PAM-2100便携式调制荧光仪(WALZ, Effeltrich, Germany)和2030-B光适应叶夹(WALZ, Effeltrich, Germany)测定叶片和苞叶PSII光适应下的最大荧光产量(Fm′ )和实际荧光产量(Ft)。设置光量子通量密度为1800 μ mol m-2 s-1, 每个样品照射3 min后, 打开饱和脉冲光进行荧光猝灭分析, PSII有效光化学效率(Φ PSII) = (Fm′ - Ft)/Fm′ , 所测叶片及苞叶与测定气体交换参数的相同。
应用Farquhar等[32]的模型, 对叶片与苞叶CO2响应曲线进行拟合, 在较低胞间CO2浓度下(Ci< 200 μ mol m-2 s-1), Rubisco活性和数量是净光合速率的限制因子, 此时, Pmax = Vc, max (Ci-Γ * )/ [Ci+Kc(1+Oi/ Ko)]-Rd; 根据von Caemmerer[33]的模型, 在较高胞间CO2浓度下(Ci> 200 μ mol m-2 s-1), RuBP再生将受到限制, 取决于电子传递速率, 此时Pmax= Jmax (Ci-Γ * )/ (4Ci+8Γ * ) -Rd; 其中Pmax为最大净光合速率, Vc, max为最大羧化速率, Jmax为最大电子传递速率, Ci为胞间CO2浓度, Γ * 为CO2补偿点, Kc和Ko是Rubisco羧化和氧化反应的米氏常数, 分别为460 μ bar和330 mbar, Oi是叶绿体羧化部位的氧分压, 为210 mbar, Rd为光下呼吸速率。根据Hymus等[34]的理论, 通过公式Rubisco (mol m-2) = (Vc, max/1 000 000)/8Kcat估算Rubisco含量, 其中Kcat是每个活化位点的羧化能力, 为3.3。
采用SPSS17.0统计软件对数据进行单因素分析, 用最小差异性检验(LSD)法检验差异显著性, P< 0.05。用SigmaPlot10.0作图。
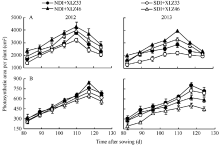
由图1可以看出, 随生育进程2个棉花品种单株叶面积(图1-A)与苞叶面积(图1-B)均表现先上升后下降的趋势, 在盛铃期达到峰值; 与正常滴灌量相比, 节水滴灌处理下棉花叶片面积显著降低, 但苞叶面积的变化无显著差异。从出苗后85~125 d, 不同水分处理下棉花苞叶面积较叶面积与苞叶面积之和变化显著, 分别增加了33.85%~39.69%和27.02%~33.10%。
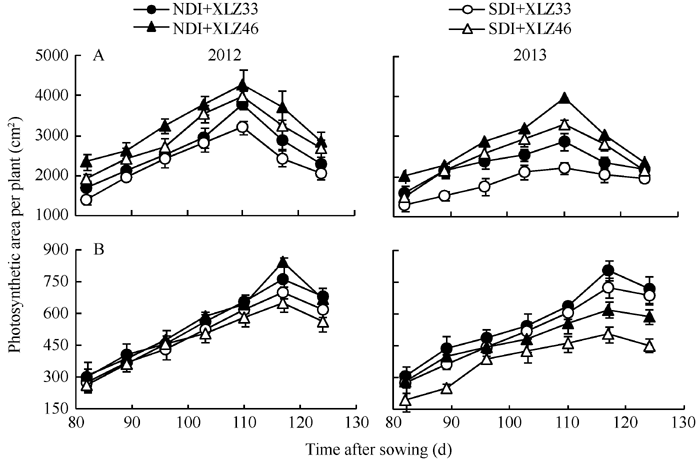 | 图1 棉花叶面积(A)与苞叶面积(B)的变化NDI: 常规滴灌量; SDI: 节水滴灌量。XLZ33: 新陆早33号; XLZ46: 新陆早46号。Fig. 1 Changes of cotton leaf area (A) and bract area (B)NDI: normal drip irrigation; SDI: saving drip irrigation; XLZ33: Xinluzao 33; XLZ46: Xinluzao 46. |
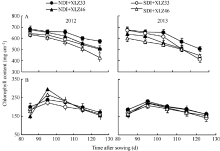
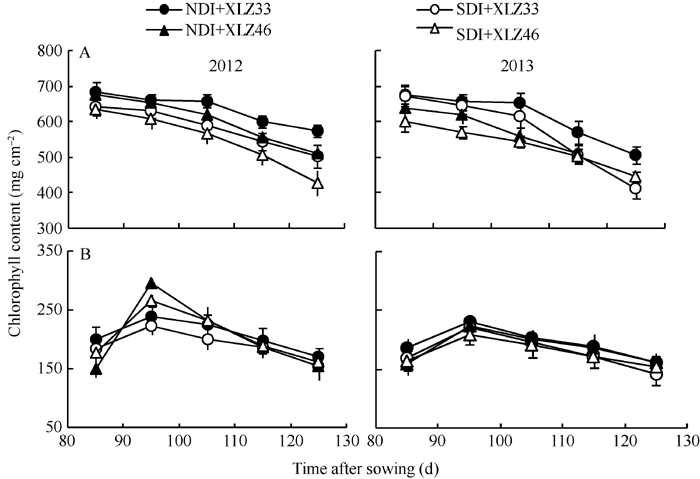
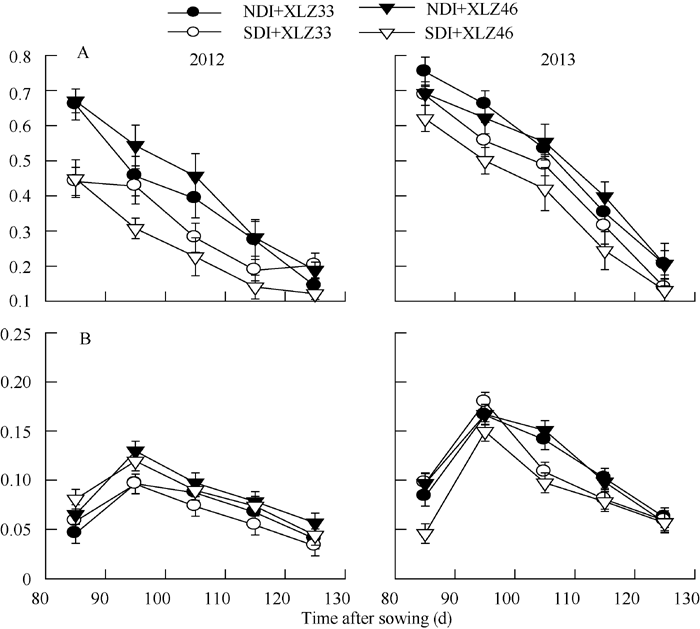
叶绿素(Chl)含量的高低影响叶片对光能的吸收、传递和转化, 生产实践中常用光合器官叶绿素的含量来判断植株遭受逆境胁迫的程度与光合生产能力等。试验表明, 单位面积苞叶的叶绿素含量显著低于叶片(图2-A, B)。出苗后85~125 d, 叶片光合机构已健全, 叶绿素含量显著下降(图2-A); 而苞叶光合机构尚未发育完全, 单位面积苞叶的叶绿素含量呈现先上升后下降的趋势, 盛铃期到吐絮期降幅较小, 差异不显著(图2-B), 但显著低于叶片。与正常滴灌相比, 节水滴灌处理下单位面积叶片的叶绿素显著降低, 但单位面积苞叶的叶绿素含量无显著性差异。
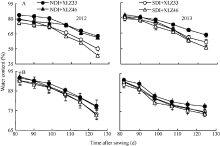
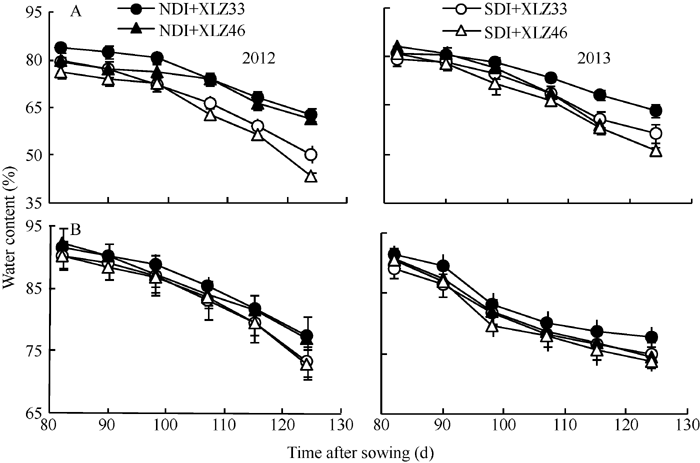
随着棉花生育时期的推移, 在田间条件下棉株叶片与苞叶的含水量均表现下降趋势, 苞叶含水量显著高于叶片含水量, 不同品种间差异较小(图3-A, B)。在测定时期内, 常规滴灌量下叶片与苞叶含水量分别下降21.67%~30.78%和15.04%~17.85%, 而节水滴灌量下分别下降28.78%~43.29%和15.87%~ 19.41%。苞叶的含水量降幅小于叶片, 表明苞叶比叶片具有更强的耐旱性。
2.4.1 净光合速率(Pn)的变化 随棉花生育时期的推移, 叶片净光合速率(Pn)逐渐下降(图4-B), 不同品种间盛花期至盛铃期差异显著, 吐絮期差异不显著; 苞叶Pn呈先上升后下降的趋势, 在出苗95 d达到峰值, 这可能与苞叶光合器官逐渐建成有关, 品种间差异不显著。与正常滴灌量相比, 节水滴灌处理下叶片Pn显著降低, 但苞叶Pn无显著差异。此时常规滴灌下苞叶光合速率最大时约占叶片的27%~29% (图4-C), 而节水滴灌下这一比值可达到35%~38%。
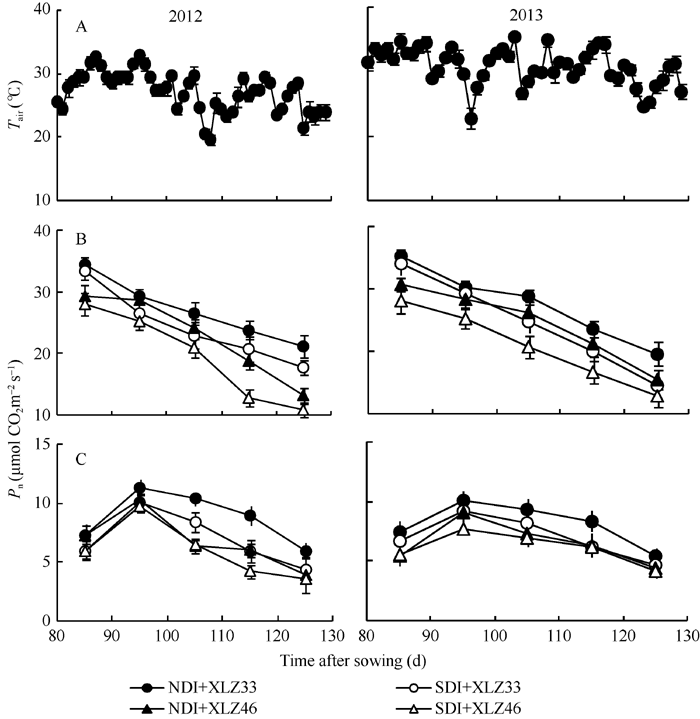 | 图4 1800 μ mol m-2 s-1光强及新疆夏季气温(A)下棉花叶片(B)与苞叶(C)净光合速率(Pn)的变化缩写同图1。Fig. 4 Changes of photosynthetic rate (Pn) in cotton leaf (B) and bract (C) under 1800 μ mol m-2 s-1 and Xinjiang’ s temperature (A)Abbreviations are the same as those given in Fig. 1. |
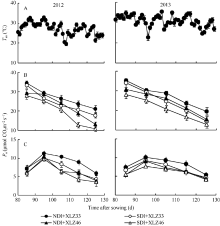
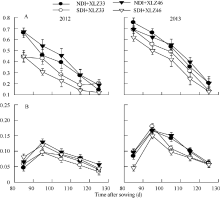
2.4.2 气孔导度(Gs)的变化 随着棉花生育时期的推移, 叶片的气孔导度(Gs)持续下降(图5-A); 苞叶的光合机构尚在发育过程中, Gs呈现由低到高的变化趋势, 在出苗后95 d左右达到峰值(图5-B), 随后逐渐下降。在新疆夏季气温(图4-A)及1800 μ mol m-2 s-1光强条件下, 叶片的Gs显著高于苞叶。与正常滴灌相比, 节水滴灌处理下叶片Gs显著降低, 不同品种间差异显著; 但不同水分处理下苞叶Gs差异不显著, 品种间苞叶Gs差异亦不显著。
 | 图5 新疆夏季棉花叶片(A)与苞叶(B)气孔导度(Gs)的变化缩写同图1。Fig. 5 Changes of of stomatal conductance (Gs) in cotton leaf (A) and bract (B) under 1800 μ mol m-2 s-1 and Xinjiang’ s temperatureAbbreviations are the same as those given in Fig. 1. |
2.4.3 最大羧化速率(Vc, max)与最大电子传递速率(Jmax)的动态变化 在新疆夏季气温(图4-A)及光强1800 μ mol m-2 s-1条件下, 棉花叶片与苞叶的最大羧化速率(Vc, max)变化趋势不同, 出苗后85 d叶片的光合机构已健全, Vc, max呈快速下降趋势(图6-A), 而苞叶Vc, max呈现先上升后下降的趋势, 且变化幅度较小(图6-C), 表明苞叶光合器官尚在逐渐发育过程中。田间条件下棉花叶片最大电子传递速率(Jmax)变化趋势与Vc, max相似(图6-B, D)。与正常滴灌相比, 节水滴灌条件下叶片Vc, max与Jmax显著降低, 但苞叶Vc, max与Jmax无显著差异, 品种间差异亦不显著。叶片Vc, max是苞叶的3.42倍左右, Jmax是苞叶的1.89倍左右, 均达显著水平。
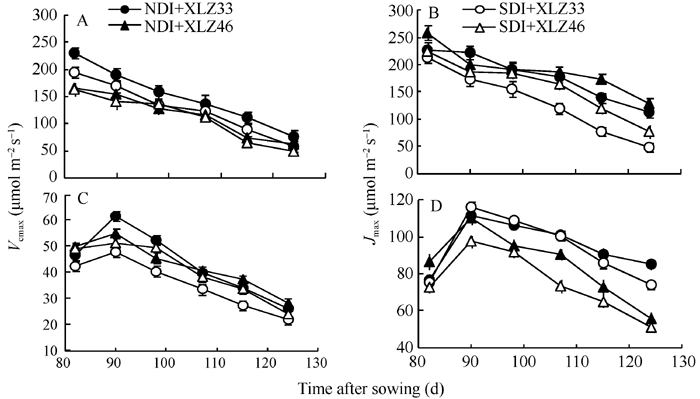 | 图6 棉花叶片Vc, max(A)和Jmax(B)与苞叶Vc, max(C)和Jmax(D)的动态变化缩写同图1。Fig. 6 Changes of the maximal velocity of RuBP carboxylation (Vc, max) and the maximum rate of electron transport driving RuBP regeneration (Jmax) in cotton leaves (A, B) and bracts (C, D)Abbreviations are the same as those given in Fig. 1. |
最大电子传递速率与最大羧化速率的比值(Jmax/ Vc, max)可用来评价Rubisco氧化和羧化作用的蛋白分配情况[35, 36]。随棉花生育时期的推移, 苞叶的Jmax/Vc, max均显著高于叶片(图7-A, B)。与正常滴灌量相比, 节水滴灌条件下叶片Jmax/Vc, max显著降低, 品种间差异显著; 但苞叶Jmax/Vc, max无显著差异, 品种间差异亦不显著。
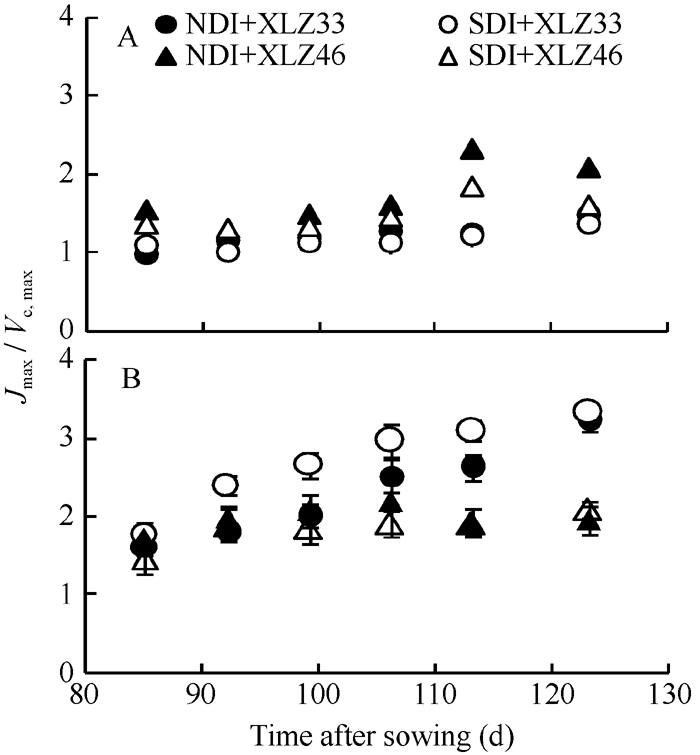 | 图7 棉花叶片(A)与苞叶(B)Jmax/Vc, max的变化缩写同图1。Fig. 7 Changes of Jmax/Vc, max in cotton leaves (A) and bracts (B)Abbreviations are the same as those given in Fig. 1. |
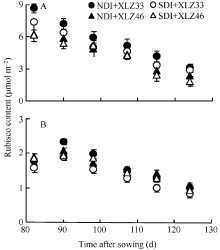
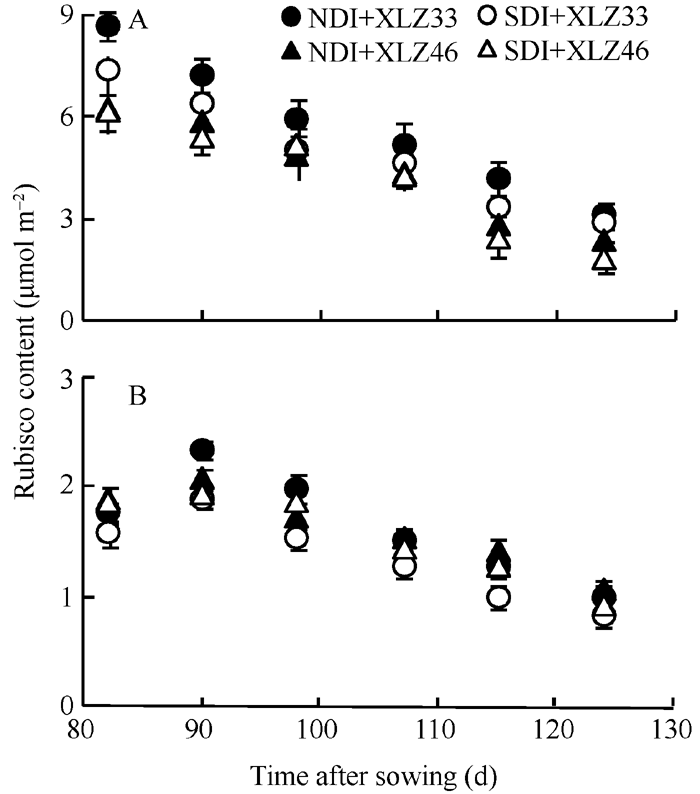
2.4.4 Rubisco含量的动态变化 在棉铃发育过程中, 单位面积叶片Rubisco含量呈降低趋势; 而苞叶光合机构尚在发育过程中, 单位面积苞叶Rubisco含量在出苗后95 d时达到峰值, 是叶片Rubisco含量的19%~24% (图8), 此后苞叶的Rubisco含量开始缓慢下降。出苗后85~125 d, 常规滴灌下叶片的Rubisco含量下降58.91%~60.25%, 苞叶的Rubisco含量下降40.49%~41.07%; 而节水滴灌下叶片Rubisco含量降低69.25%~74.71%, 苞叶降低45.17%~47.65%。
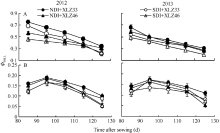
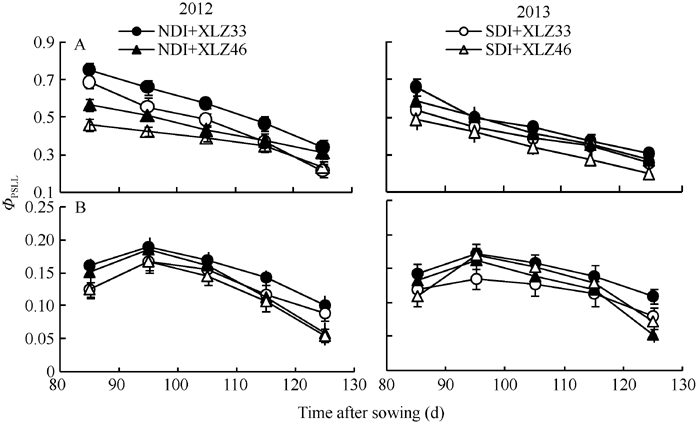
在环境胁迫下, 实际光化学效率(Φ PSII)的变化能反映PSII光合能力的高低[27, 37, 38, 39]。在本试验条件下, 随生育进程, 当光强为1800 μ mol m-2 s-1时, 叶片的Φ PSII呈下降趋势(图9-A), 品种间差异显著; 至出苗后125 d, 常规滴灌与节水滴灌下叶片的Φ PSII分别降低45.36%~53.38%和58.57%~68.72%。棉花苞叶的Φ PSII变化趋势与叶片存在明显差异, 出苗后85 d苞叶Φ PSII呈先上升后下降的趋势, 表明苞叶光合器官尚在发育过程中, 至出苗后95 d达到峰值(图9-B), 变化趋势与Pn一致, 光系统II Φ PSII降低直接导致其较低的Pn; 苞叶的Φ PSII下降缓慢且幅度较小, 品种间差异不显著; 至出苗后125 d, 常规滴灌与节水滴灌下苞叶的Φ PSII分别降低29.25%~34.71%和33.17%~37.65%。
本研究结果表明, 2个棉花品种在出苗后85~ 125 d, 叶片Rubisco含量快速下降, 而苞叶仍能保持较稳定的Rubisco含量(图8-B); 叶绿素降解是叶片衰老的表现, 随棉铃的快速生长, 苞叶的叶绿素含量下降幅度显著低于叶片(图2-B)。在棉铃发育后期, 苞叶Rubisco含量和叶绿素含量下降幅度均小于叶片, 这表明苞叶的衰老较慢。节水滴灌条件下叶片Pn、Gs、Jmax与Vc, max下降显著, 而苞叶Pn、Gs、Jmax与Vc, max变化不显著, 表明土壤水分不足, 叶片的光合能力受到明显抑制, 而苞叶能保持相对稳定光合能力。相关分析表明, 苞叶能保持相对稳定光合能力与其具有较稳定的含水量和相对稳定的光合组分(叶绿素含量、Rubisco含量)呈显著正相关。与叶片相比, 苞叶具有较低的气孔导度和较高的水分利用效率[40, 41, 42], 使得苞叶具有较为抗旱的形态特征和生理机制。前人研究表明, 在棉铃发育的初期, 主茎倒二叶的光合作用能为倒二果枝第一果节的棉铃提供足够的碳水化合物, 但在棉铃发育中后期, 主茎倒二叶衰老、光合能力下降不能再满足棉铃的发育[15, 43], 棉铃发育所需的能量主要由新的“ 光合源” 提供。在棉花生育后期, 叶片逐渐衰老脱落, 苞叶表面积的相对增加且其光合能力维持的时间较长, 因此苞叶的光合作用可能是棉铃发育所需能量的新“ 源” 之一。
强光下绿色器官捕获的激发能超过其碳同化的利用能力时, 过剩光能会导致PSII的光抑制[39]。Demmig和Adams[44]认为光合器官通过降低PSII实际光化学效率以减轻光抑制。本研究结果表明, 随生育进程叶片与苞叶的Φ PSII均呈下降趋势(图9), 苞叶的下降幅度显著低于叶片; 与正常滴灌量相比, 节水滴灌条件下叶片Φ PSII显著降低, 而苞叶变化不显著, 叶片Pn下降幅度显著高于苞叶, 表明苞叶具有一定的抗旱性。棉花苞叶的Pn(图4)与Gs (图5)显著低于叶片, 对Pn、Gs与胞间CO2浓度(Ci)进行相关性分析表明, Pn、Ci与Gs呈显著正相关, 较低的Gs可能是苞叶Pn的限制因素, 另一方面, 较低的叶绿素含量也是苞叶Pn较低的重要原因。
Farquhar等[32]认为当环境中的CO2浓度较高时, RuBP的氧化速率将成为限制光合速率的因子; Hu等[25]研究发现, 棉铃具有较高的呼吸速率, 对其包裹在外的苞叶形成了一个高浓度CO2的微环境。本试验表明, 苞叶具有较高的Jmax/Vc, max比值(图7)和Gs (图5), 表明苞叶对较高浓度CO2的适应能力高于叶片。节水滴灌条件下叶片Jmax/Vc, max比值和Gs显著降低, 但苞叶Jmax/Vc, max比值和Gs无显著性差异, 暗示节水滴灌条件下苞叶具有适应较高浓度CO2的能力, 同时维持稳定的光合作用。
前人研究表明, 小麦穗的光合作用占麦粒所需碳水化合物的40%~50%[40, 45], 水稻花序的光合作用占谷粒所需碳水化合物的20%~30%[46]。因此, 作物非叶绿色器官光合作用对产量形成具有重要的生理意义。在棉花盛铃中期至吐絮初期叶片快速脱落, 相对而言, 苞叶面积增加, 苞叶面积占光合面积的比例显著增大(图1-B)且能保持较稳定的光合速率(图4-B)。因此, 随着叶片的衰老, 苞叶的光合作用可能对整个棉株的光合作用起着重要补充作用。Hu等[24]研究发现在棉铃的整个发育时期内, 茎秆能保持相对稳定的表面积, 约占棉株总面积的15.6%~ 17.3%, 但茎秆单位面积的叶绿素含量较低, 光合能力有限; 随着棉铃的发育, 棉铃面积逐渐增加, 吐絮期棉铃面积约占棉株总面积的6.9%~8.1%, 但棉铃单位面积的叶绿素含量较低, 光合能力有限。根据前人对棉花非叶器官光合作用的估算[15, 24, 40, 43], 分别将出苗后85~125 d叶片、苞叶、铃壳和茎秆的净光合速率平均值与其面积平均值相乘, 估算出常规滴灌下苞叶的光合能力占整个棉株光合能力为3.65%、4.71%、5.56%、7.22%和8.83%; 而节水滴灌下这一比值可达到4.78%、6.64%、7.87%、10.24%和12.53%。
棉花经济产量主要在盛铃期至吐絮期形成的, 生育后期叶片衰老较快, 而苞叶能保持较稳定的光合能力, 在节水滴灌条件下, 棉花苞叶的光合作用对植株整体光合能力的贡献率增加。因此, 在水分不足条件下, 可通过膜下滴灌合理的水肥管理, 适当控制叶面积指数的发展, 促进苞叶等非叶绿色器官面积的发展, 调节叶铃空间分布, 充分发挥非叶绿色器官的光合抗逆功能, 提高棉花产量。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|