



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
GECROS模型在黄淮海地区模拟夏玉米生长的适应性评价
[吴玮1, 2, 3  , 马玉平
, 马玉平1, * , 俄有浩1 , 孙琳丽1, 2 , 景元书2 ]
, 马玉平, 俄有浩|
|


第一作者联系方式: E-mail:brilliant_ww@163.com
GECROS是荷兰瓦赫宁根农业大学近些年开发的机理性更强、算法更简要的作物生长模型。本文利用黄淮海地区夏玉米试验数据进行GECROS模型的适应性评价, 为模型进一步开展区域应用提供依据。结果表明, GECROS基本能够反映黄淮海地区夏玉米的发育进程。模型模拟夏玉米抽雄期的绝对偏差在6.0 d以内, 平均为2.1 d; 模拟成熟期的绝对偏差在8.0 d以内, 平均为3.4 d。GECROS描述夏玉米干物质积累和叶面积扩展过程的准确度较高。模拟雌穗总重的归一化均方根误差在7.8%~33.8%之间, 平均为18.6%; 模拟植株地上总重的归一化均方根误差在11.2%~32.6%之间, 平均为20.7%; 模拟LAI的绝对偏差在0.28~0.55之间, 平均为0.41, 模拟籽粒产量的绝对偏差在20.3~229.0 g m-2之间, 平均为80.9 g m-2。利用GECROS模型相对评价作物生长状况或环境影响基本可行。但GECROS模拟夏玉米发育进程仍存在低值偏高、高值偏低的现象; 在土壤水分胁迫较重时, 描述的生物量积累过程有偏低情况; 描述LAI扩展的总体效果差于生物量累积的效果。GECROS仍需进一步完善。
The evaluation of crop model is a key process for its application. GECROS model had been developed by Wageningen in recent years. GECROS uses stronger mechanism and more concise algorithms to summarize the current knowledge of individual physiological processes and their interactions and feedback mechanisms. To provide a foundation for the future localization and regional application of GECROS model, in this study, the field observations of summer maize from several agrometeorological stations in Yellow-Huaihe-Haihe Rivers were used to conduct the adaptability evaluation of GECROS model. The results showed that GECROS model could basically reflect the growing process of summer maize in Yellow-Huaihe-Haihe Rivers. The absolute deviations at the period from emergence to tasseling simulated by GECROS were less than 6.0 d, with an average of 2.1 d. The absolute deviations at the period from tasseling to mature were less than 8.0 d, with an average of 3.4 d. The dry matter accumulation and leaf area expansion process of summer maize were accurately described by GECROS. The normalized mean square root errors (%) of total ear weight simulated by GECROS were 7.8%-33.8%, with an average of 18.6%, and these of total plant weight were 11.2%-32.6%, with an average of 20.7%. The absolute deviations of LAI were 0.28-0.55, with an average of 0.41. The relative evaluation on crop growth and impact of environmental conditions was basically feasible. But the developmental process of summer maize simulated by GECROS had the phenomenon of low values tengding to higher and high values tengding to lower. When the soil water stress was severe, GECROS gave a lower value for the biomass accumulation process, and the overall effect of description for the LAI expansion was inferior to that for biomass accumulation. GECROS still needs to be further improved.
作物生长模型是研究农业生产的一种重要手段, 它不仅反映了现阶段对整个农业生产系统的认知水平, 还能用于研究和比较不同生态环境下作物生长的差异, 促进人们对作物生长机理的深化理解和研究, 反过来又提高模型模拟能力[1, 2, 3, 4]。至今, 作物生长模型虽然被不断完善, 应用性和适应性不断增强, 但由于气象等环境灾害因素对农业生产影响的多方面和复杂性, 模型的诸多生理生态过程仍需深入研究并进一步完善[5, 6]。其中, 荷兰deWit学派作为国内外众多模型研究派系之一, 先后自主研发了一系列作物生长模型, 包括ELCROS、BACROS、SUCROS、MACROS等[7, 8]。为了能更好地面向实际应用, 荷兰瓦赫宁根农业大学与世界粮食研究中心合作, 在SUCROS基础上研制了能模拟特定土壤和气候条件下的一年生作物生长动态的、解释性的WOFOST模型; 又与菲律宾国际水稻研究所(IRRI)合作, 在MACROS和SUCROS的基础上研发了水稻模型ORYZA, 再不断完善后发展成系统性、应用性更强的水稻生长模拟模型ORYZA2000[9]。这两者在国内外都有较广泛的验证和应用[10, 11]。2005年以来, 为了进一步完善作物生长模型的机制过程和克服部分模块仍利用的数理统计概括, 荷兰瓦赫宁根农业大学在之前deWit学派众多模型基础上开发了机理性更强、算法更简要的GECROS模型[12]。该模型可用于研究作物与环境的相互作用。GECROS基于“ 钟模型” [13]的改进来描述发育进程, 克服了以往积温模型的不足; 采用了Farquhar光合生理模型, 较以往光合模型的统计形式更具有机理性。GECROS较以往模型有较大改动, 其模拟和适应能力有待多方检验。目前在中国区域的应用验证研究还很少。
本文利用黄淮海地区农业气象观测站的夏玉米田间试验数据, 进行GECROS模型在潜在和水分胁迫条件下的发育进程、叶面积以及生长量的模拟检验, 以评价GECROS模型的模拟能力与适应性, 为进一步开展模型区域应用提供依据。
GECROS是分析作物品种和环境相互作用的作物生长模型[12]。它对作物发育进程、光合呼吸过程、叶面积扩展以及土壤水分平衡等主要机理描述如下。
GECROS综合考虑了温度和日长与作物发育的非线性关系, 在“ 钟模型” 基础上改进模拟发育期。它以Farquhar模型为基础, 从生理学角度出发构建光合作用模型。
PP=44× 10-6× (1-Γ * /CC) min (VC, VJ) (1)
式中, PP为潜在总光合速率(g m-2s-1); Γ * 为在没有暗呼吸下的CO2补偿点(μ mol mol-1); CC为进行羧化作用的CO2含量(μ mol mol-1)。VC为RuP2羧化速率(μ mol m-2s-1), VJ为电子传输速率(μ mol m-2s-1)。该模型从作物叶片生长过程的生理学角度出发, 根据Yin等[12]提出的叶面积指数碳氮限制原则, 将其分为碳限制(LC)和氮限制(LN)两部分。
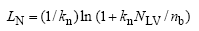
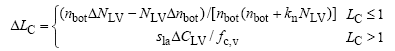
式中, kn为氮消亡系数; nb为能进行光合作用的最小叶氮含量(g m-2); NLV为活叶氮含量(g m-2), Δ LC为LC日增量, Sla为比叶面积(m2 g-1); CLV为活叶碳含量(g m-2); nbot为底层叶片氮含量。叶面积指数L=min (LC, LN)。当土壤水分充足时, 作物光合速率决定蒸腾速率。当作物根系吸收的土壤水分不能满足潜在蒸腾EP时, 实际蒸腾Ea由根活动层的土壤含水量决定, 实际蒸腾速率则决定实际光合速率[12]。潜在蒸腾EP依据彭曼公式[12]计算。
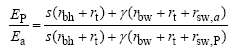
式中, rt为作物冠层湍流阻力(s m-1); rbh和rbw分别为叶片边界热量和水分传递阻力(s m-1); γ 为干湿表常数(kPa ℃-1); s为饱和水汽压随温度变化斜率(kPa ℃-1); rsw, p和rsw, a分别为潜在和实际水分条件下的叶片气孔阻力(s m-1)。
GECROS模型包含许多体现品种遗传特性及生态适应性的参数(表1), 它的输入量主要为逐日日照时数、最高、最低温度、降水量、平均水汽压和平均风速, 输出量主要包括作物碳氮元素的吸收量、蛋白质含量、冠层光合速率、蒸腾速率、叶面积指数、地上总干重、产量和收获指数等要素。
| 表1 GECROS模型的主要参数及其取值范围 Table 1 The ranges of the main parameters of GECROS model |
1.2.1 模型参数校准方法 对于GECROS模型发育参数, 根据黄淮海夏玉米实际发育期及对应温度数据推算确定。对于模型生长参数, 首先进行敏感性分析, 然后再对敏感参数依据性质分类确定其取值。
考虑到不同生长参数对作物生长影响的差异性, 在调试参数之前, 首先进行参数的敏感性分析, 以确定其影响作物生物量积累和产量形成的重要程度, 从而更有针对性地进行参数本地化。敏感性分析方法为, 改变某一参数的数值(变幅分别取-10%、-20%、10%和20%)而其他参数值保持不变, 运转模型计算生长量(逐日总重WSH、穗重WSO和叶面积指数LAI)在全生育期的平均相对变化率, 由此确定相对敏感度(Sr), 即全生育期内参数每变化1%时生长量的变化, 以判断该参数对作物生长发育的影响程度[14]。

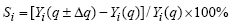
式中, Yi为第i日的模拟生长量, q为不同参数的初始值, Δ q为各个输入参数的变化量, i为日, n为发育总天数。
依据物理化学特性, GECROS模型的敏感生长参数可分为两类。第一类为生物化学参数, 主要与作物种类或品种有关, 一般具有较明确的生物学意义和普适性[12], 可以利用作物的生化分析获得参数取值。如贮存器官各类营养元素比例、25℃时RuP2最大羧化速率等。其中个别参数也可利用田间试验的物理数据进行反推校准。如根茎叶最小含氮比例。第二类为生物物理参数, 主要与耕作措施、环境条件及作物品种有关, 需要利用田间试验的物理观测数据直接计算或校准。主要包括出苗率、叶片宽度、最大株高、最大根深、粒重以及比叶面积等。
1.2.2 模拟检验方法 首先绘制作物生长模型模拟与实测结果的时间变化曲线或1∶ 1线, 进行模型模拟性能的直观检验。然后统计模拟结果的各种误差进行模拟性能的定量检验。误差分析包括绝对偏差均值(AD)、均方根误差(RMSE)和归一化均方根误差(NRMSE)[15, 16]。
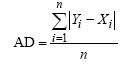
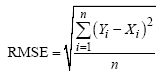
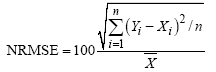
式中, Yi和Xi分别为模拟值和实测值, 
作物数据为黄淮海地区5个农业气象观测站夏玉米生长发育观测结果。一为1991— 2011年河北定州、1994— 2011年山东泰安、1991— 2010年河南郑州和2011年安徽宿州夏玉米发育期数据。二为2009— 2011年河北固城、2010年河南郑州、2011年山东泰安和2011年安徽宿州夏玉米水分控制田间试验数据。其中, 河北固城试验包含4个水分控制和1个对照处理(CK)。水分控制分别为偏多(K1)、基本适宜(K2)、轻度干旱(K3)和重度干旱(K4), 土壤湿度分别是田间持水量的80%以上、55%~80%、40%~55%和40%以下。K1~K4处理为8 m2(2 m × 4 m)池栽, 每池播种50~60株玉米; CK为大田播种, 水分供应不受限制, 视为潜在水分条件处理。河南郑州、山东泰安和安徽宿州试验均为潜在水分条件处理。供试夏玉米为近年来农业部主推黄淮海地区栽培品种, 河北定州和固城均为广源旺禾94-9, 河南郑州、山东泰安和宿州分别为浚单20、登海605和郑单958。河北固城、河南郑州、山东泰安和安徽宿州种植密度分别为6.9、6.3、5.5和7.5株 m-2。试验观测项目包括玉米生育期(播种期、出苗期、三叶期、七叶期、拔节期、抽雄期、开花期、吐丝期、乳熟期和成熟期), 主要发育期的植株地上总干重、穗干重和LAI。另外, 气象数据包括相应年份和站点的逐日日照时数(h)、最高温度(℃)、最低温度(℃)、降水量(mm)、平均水汽压(kPa)和平均风速(m s-1), 数据来自国家信息气象中心。土壤数据包括田间持水量、凋萎湿度以及氮磷等养分含量, 由中国科学院南京土壤研究所提供。
限于试验观测数据较少, 且河北定州、山东泰安、河南郑州和安徽宿州等4个站点试验所用玉米品种特性差异较小[17, 18], 因此本文不再分玉米品种进行模型校准和检验。利用2000— 2008年河北定州、山东泰安、河南郑州和2011年安徽宿州玉米发育期数据进行GECROS模型发育参数的校准(Calibration)及回代检验。利用2009年河北固城夏玉米田间试验CK处理数据进行模型潜在生长参数校准及回代检验、K1~K4处理数据进行不同土壤水分条件下生长参数的校准及回代检验。利用1991— 1999和2009— 2011年河北定州、1994— 1999和2009— 2011年山东泰安、1991— 1999和2009— 2010年河南郑州夏玉米发育期数据进行发育模拟的独立样本检验(Validation)。利用河北固城2010— 2011年K1、K2和CK处理、河南郑州2010年、山东泰安2011年、安徽宿州2011年的田间试验数据进行水分充足条件下夏玉米生长量模拟的独立样本检验。利用河北固城2010— 2011年K3和K4处理的田间试验数据进行水分胁迫条件下夏玉米生长量模拟的独立样本检验。以模型的检验效果评价其适应能力。
2.1.1 GECROS模型发育参数校准及回代检验
GECROS的发育参数包括玉米发育下限、上限、最适温度、温度响应系数(TS)、光照敏感起止阶段、光照敏感系数(PESN)、基本营养生长日(MTDV)和基本生殖生长日(MTDR)等。玉米发育的下限、适宜和上限温度根据前人研究成果确定[19], 分别取8℃、30℃和42℃。温度响应系数反映品种感温性[20]。光照敏感起始终止阶段, 取模型默认值0.2和0.7。光照敏感系数反映品种的感光性, 与品种的最适光长有关。当实际光长大于最适光长时, 玉米的光照敏感系数为1, 当实际光长小于最适光长时, 玉米的光照敏感系数与最适光长呈负相关。基本营养生长日和基本生殖生长日为最适条件下营养生长阶段和生殖生长阶段所需天数。表2为利用黄淮海夏玉米数据校准发育参数后的取值。可以看出, 各参数取值在不同站点(品种)之间有一定差异。黄淮海北部地区夏玉米品种的基本生长日较南部更长, 光敏感系数则更低。
| 表2 GECROS模型中夏玉米发育参数的校准值 Table 2 Calibrated values of development parameters for summer maize in GECROS model |
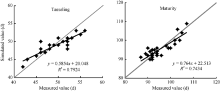
抽雄期和成熟期分别作为夏玉米营养生长阶段和生殖生长阶段的终止时期, 是评价模型模拟夏玉米发育期效果的重要指标之一。图1表明, 模拟抽雄期的绝对偏差在4 d以内, 绝对偏差均值(AD)为1.4 d, 模拟值与实测值的线性相关系数为0.89; 模拟成熟期的绝对偏差在5 d以内, 绝对偏差均值(AD)为2.3 d, 线性相关系数为0.86。
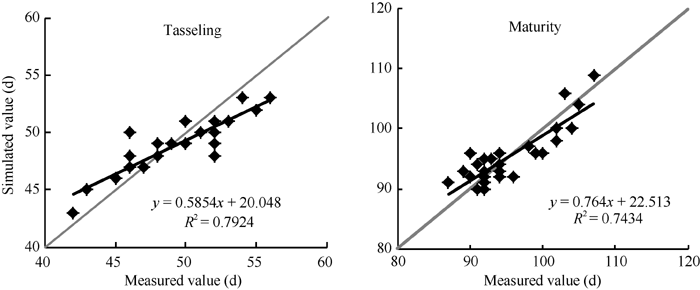 | 图1 GECROS模型模拟黄淮海夏玉米发育进程的回代检验Fig. 1 Back substitution test of GECROS simulating the development process of summer maize in the Yellow-Huaihe-Haihe Rivers |
2.1.2 GECROS模型生长参数校准及回代检验
利用河北固城2009年潜在条件下夏玉米试验数据进行GECROS 模型生长参数的敏感性分析, 获得各参数对模型不同状态变量(逐日总重WSH、穗重WSO和叶面积指数LAI)的相对敏感度及其平均值(表3)。总体来看, 初始叶片含氮比例(ncri0)的敏感性最高, 对3个状态变量的平均相对敏感度达0.85%。比叶面积(Sla)次之, 为0.72%。再次为粒重(Sw)和出苗率(Eg), 均为0.52%。但参数对不同状态变量的影响程度有差异, 如出苗率对LAI的影响较大, 平均相对敏感度可达0.80%, 而对WSO的影响较小, 为0.22%。参数的正负变化对变量的影响也可能存在区别。如茎干物质密度(ρ )增大时, 对LAI的平均相对敏感度可达0.30%, 而当其降低时仅为0.03%。
| 表3 GECROS模型生长参数的相对敏感度 Table 3 The relative sensitivity of crop growth parameters in GECROS (%) |
根据生长参数物理化学特性及敏感性, 利用2009年河北固城田间试验潜在水分条件的夏玉米数据计算获得最大根深(Dmax)、叶片宽度(w)、茎干物质密度(ρ )、最大株高(Hmax)、出苗率(Eg)、粒重(Sw)、比叶面积(Sla)和种植密度(NPL)等参数的初始取值。再利用“ 试错法” 对初始叶片含氮比例(ncri0)、出苗率(Eg)和茎干物质密度(ρ )等参数进行适当调整, 最终确定了各参数的取值(表4)。利用水分胁迫条件下的夏玉米生长数据调试确定了作物每日供水量Wput (表4)。可以看出, 校准后的参数取值与缺省值相比均有一定变化。如黄淮海夏玉米的最大株高更高, 可达2.8 m, 种植密度略偏低, 为8株 m-2。出苗率(Eg)的变化最为明显, 黄淮海地区可达0.7 g g-1。夏玉米每日供水量约为3 mm d-1。
| 表4 GECROS模型中夏玉米主要生长参数的校准值 Table 4 Calibrated values of growth parameters for summer maize in GECROS |
由图2可以看出, 模拟地上总重、穗重与实测值非常接近; 模拟LAI在玉米生长前期增长迅速, 与实际情况比较一致, 但在中后期由于受碳氮限制而下降幅度较实际稍大。表5为模拟结果的误差统计。结果表明, 模拟夏玉米植株地上总重和雌穗总重的平均值略低于实测值, 模拟LAI的平均值略高于实测值。模拟植株地上总重和雌穗总重的绝对偏差均值(AD)与均方根误差(RMSE)均小于80 g m-2, 模拟LAI的AD与RMSE均小于0.7, 模拟籽粒产量与实测值相差132.1 g m-2。所有模拟结果的归一化均方根误差(NRMSE)均小于25%。t检验结果显示, P(t)值都大于0.05, 表明模拟值与观测值无显著差异。
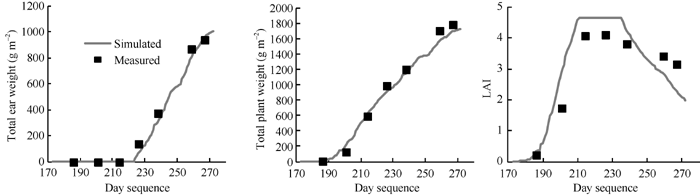 | 图2 GECROS模型模拟河北固城2009年潜在条件下夏玉米生长量和实测值的比较Fig. 2 Comparison of biomass simulated by GECROS and measured for summer maize under potential condition (CK) at Gucheng of Hebei in 2009 |
| 表5 GECROS模拟河北固城2009年潜在条件下夏玉米生长量的误差分析 Table 5 Errors of GECROS simulating summer maize growth under potential condition at Gucheng in 2009 |
由图3可以看出, 雌穗和植株地上总重的总体模拟效果良好, 线性相关系数均大于0.97; LAI的模拟效果稍差, 水分充足和水分胁迫条件的相关系数分别为0.89和0.88。表6为模拟结果的误差统计。总体来看, 各种水分条件下模拟生长量及LAI与实测值无显著差异[P(t)> 0.05], NRMSE均小于26%, 植株地上总重与LAI的模拟均值略高于实测值, 雌穗总重与籽粒产量的模拟均值略低于实测值。另外, 水分充足条件下模拟植株地上总重整体有所偏大。
 | 图3 GECROS模型模拟河北固城2009年不同土壤水分条件夏玉米生长量和实测值1∶ 1图a和b分别代表水分充足和水分胁迫条件。Fig. 3 The 1:1 diagram of biomass simulated by GECROS and measured for summer maize under different water conditions at Gucheng of Hebei in 2009a: adequate soil moisture; b: soil water stress. |
| 表6 GECROS模拟河北固城2009年不同土壤水分条件下夏玉米生长量的误差分析 Table 6 Errors of GECROS simulating summer maize growth under different water conditions at Gucheng of Hebei in 2009 |
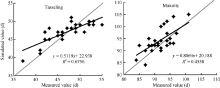
2.2.1 GECROS模型模拟夏玉米发育进程的适应性评价 图4表明, 模型模拟抽雄期、成熟期与实测值的线性相关系数分别为0.82和0.67。但模拟抽雄期存在低值偏高、高值偏低的现象; 模拟成熟期略偏长。模拟误差统计表明, 模拟的绝对偏差在6 d以内, 绝对偏差均值(AD)为2.1 d; 模拟成熟期的绝对偏差在8 d以内, 绝对偏差均值(AD)为3.4 d。t检验结果显示, 抽雄期和成熟期的P(t)均大于0.05, 模拟值与观测值无显著差异。
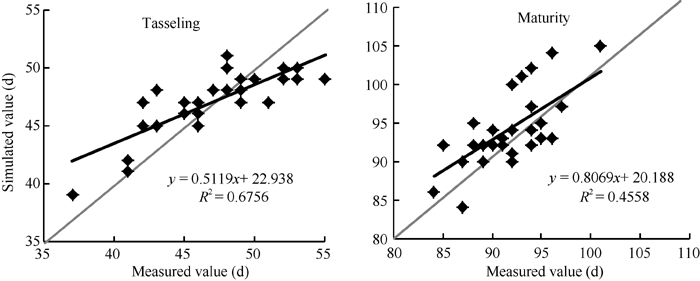 | 图4 GECROS模型模拟黄淮海夏玉米发育进程的独立样本检验Fig. 4 Validation of GECROS simulating development process of summer maize in the Yellow-Huaihe-Haihe Rivers |
2.2.2 GECROS模型模拟土壤水分充足条件下夏玉米生长的适应性评价 从图5可以看出, 植株地上总重、雌穗总重和LAI模拟值与实测值总体较接近, 线性相关系数分别为0.98、0.96和0.93。由模拟误差统计结果(表7)可知, 各生长量的模拟值与实测值均无显著差异[P(t)> 0.05], 模拟效果良好。总体来看, 植株地上总重和雌穗总重的模拟效果要好于LAI的效果。其中, 除河南郑州外, 模拟植株地上总重和雌穗总重的NRMSE (%)均小于21%, 而模拟各站LAI的NRMSE (%)均大于23%, 安徽宿州和河南郑州两站LAI的NRMSE (%)均大于30%, 模拟籽粒产量的绝对偏差在41.6~229.0 g m-2之间。单独看, 河北固城2010— 2011年和山东泰安2011年的模拟效果较好, 模拟植株地上总重、雌穗总重和LAI的NRMSE (%)均小于30%; 安徽宿州除模拟LAI外, 其余模拟生物量的NRMSE (%)均小于30%; 河南郑州除模拟植株地上总重, 其余模拟生物量的NRMSE(%)均大于30%。
 | 图5 GECROS模型模拟土壤水分充足条件下夏玉米生长量与实测值1∶ 1图Fig. 5 The 1:1 diagram of biomass simulated by GECROS and measured for summer maize under adequate soil moisture condition |
| 表7 GECROS模拟土壤水分充足条件下夏玉米生长量的误差分析 Table 7 Errors of GECROS simulating summer maize growth under adequate soil moisture condition |
2.2.3 GECROS模型模拟土壤水分胁迫条件下夏玉米生长的适应性评价 图6表明, 植株地上总重、雌穗总重和LAI模拟值与实测值总体较接近, 线性相关系数分别为0.98、0.93和0.94。由模拟误差的统计分析结果(表8)可知, 2010年和2011年各生长量模拟值与实测值无显著差异[P(t)> 0.05], 2011年模拟检验结果整体好于2010年。2011年各生长量NRMSE (%)均小于30%, 而2010年植株地上总重、雌穗总重NRMSE (%)均大于30%。另外, 土壤水分充足的模拟效果总体好于水分胁迫条件。其中, 土壤水分充足下模拟植株地上总重、雌穗总重和籽粒产量的NRMSE (%)均小于20%, LAI均小于30%; 而水分胁迫条件下模拟植株地上总重、雌穗总重和籽粒产量, 除2011年雌穗总重NRMSE (%)小于20%以外, 其余均大于20%, 水分胁迫条件下模拟LAI的NRMSE (%)也均大于水分充足条件。
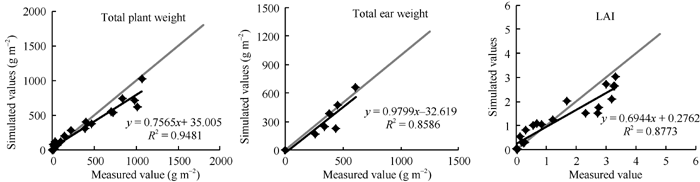 | 图6 GECROS模型模拟土壤水分胁迫条件下夏玉米生长量与实测值的1∶ 1图Fig. 6 The 1:1 diagram of biomass simulated by GECROS and measured for summer maize under soil water stress |
| 表8 GECROS模拟土壤水分胁迫条件下夏玉米生长量的误差分析 Table 8 Errors of GECROS simulating summer maize growth under soil water stress condition |
发育期是作物的生理年龄, 它与作物的形态变化相联系, 是作物生长达到质变的标志性阶段, 准确描述发育进程是作物生长模型的首要任务。基于WOFOST修改而成的“ 东北玉米生长模型” (NEC_ MaGM)在模拟东北春玉米时, 其模拟抽雄期和成熟期的绝对偏差均值分别为3.6 d和5.9 d [3]。本研究中, GECROS模拟华北夏玉米抽雄期和成熟期的绝对偏差均值分别为2.1 d和3.4 d。尽管东北春玉米和华北夏玉米的生育期长度和发育进程有差异, 但根据模拟误差基本可以认为, GECROS在玉米发育进程方面的性能与NEC_MaGM相当或略优。尽管GECROS模型考虑了温度与作物生长发育的非线性关系, 是钟模型[13]的改进, 但它描述夏玉米发育进程的灵敏性仍偏低。本文在模型发育参数校准和独
立样本检验中都发现, GECROS模拟发育期的变化范围较实测小, 模型的敏感性偏低。由此导致模拟结果出现低值偏高而高值偏低的现象, 模拟抽雄期尤其如此。这一现象在“ 东北玉米生长模型” (NEC_ MaGM)所采用发育单位法描述玉米发育进程的模拟中也曾出现[3]。但在华北冬小麦发育模拟中, 即使最简单的积温模型也未显现[21]。而在ORYZA2000水稻模型使用的热量单位法模拟中也较少见[22, 23]。究其原因, 主要可能是玉米品种基因型间的差异及基因型与环境互作较大。一般情况下, 尽管一个地区种植的玉米品种类型不但多且更新快, 但它们应该都是能够适应当地气候环境条件的品种。主栽品种之间的生育特性差异不会太大[17, 18]。本研究分析黄淮海地区玉米发育期也发现, 各站点玉米全生育期多年平均差不超过2 d, 种植起始日均值不超过3 d。因此, 为了全面体现一地的玉米品种型, 本研究综合长年代多个品种发育期数据进行作物生长模型发育参数的校准和检验, 以扩展模型的时空应用范围。这种情况下, 当利用年份较短且品种不变的田间试验数据进行模型参数校准和检验时, 作物发育模型的描述效果较好; 而当数据年份较长时, 玉米品种基因型与环境互作使得同一品种的生育特性可能随环境发生较大变化, 或者种植品种发生改变使基因型间的差异显现出来, 而作物生长模型本身对发育进程进行了简化描述, 对于多因素协同影响以及作物品种对逆境(如土壤水分亏缺)的反应力也未予考虑, 因此模拟发育的灵敏度有所下降。总之, 这一现象还需要更多数据、更多模型以及更多作物之间的比较才能得出合理结论并改进发育模型。
在玉米生长方面, 首先, 各器官生物量是作物生长模型所重点描述的状态变量, 其中玉米雌穗干重直接与产量形成相关。孙琳丽等[24]通过WOFOST模型与华北地区玉米观测数据的同化得到了模型最优参数组合和取值, 然后模拟了玉米生长状况。其模拟地上总干重和雌穗干重的均方根误差(RMSE)在30.3~144.2 g m-2之间, LAI的RMSE在0.60~0.95之间。本文利用GECROS模型模拟华北夏玉米地上总干重和雌穗干重的RMSE在42.8~195.7 g m-2之间, LAI的RMSE在0.35~0.76之间。初步比较表明两模型模拟能力的差异并不大。其次, 土壤水分是影响作物生长最重要的因素之一, 描述土壤水分的胁迫是作物生长模型的重要内容。GECROS模型不但将作物的光合作用与水分在土壤、作物及冠层间传输的阻力相联系, 而且考虑了土壤水分对作物根系吸收营养元素的影响[12]。这比以往模型仅用相对蒸腾校正光合速率的理论基础要更加扎实[25]。但本研究表明, GECORS模拟土壤水分充足条件下夏玉米生长的效果仍要好于水分限制条件的效果。可见水分限制对玉米叶片生理结构的影响机制仍需进一步探讨。第三, 叶片是作物进行光合作用生成干物质的重要器官, 叶面积扩展也成为作物生长模型的重要组成部分。GECROS模型模拟LAI是以叶片生长营养元素积累和损失的方式进行, 在作物生长模型对LAI模拟的众多方法中[26], 属生理法[27], 理论基础较好。但该模型模拟LAI的总体效果仍差于生长量的效果。这一方面是由于GECROS模型描述叶片营养元素积累和损失时未考虑不同发育阶段的差异。如比叶面积等参数总是保持不变, 而这与实际情况不符。另一方面, 模型中LAI的消亡仅由氮素转移速率决定, 而未考虑玉米生长后期叶片碳素的限制、氮素增长微弱以及LAI形态性消亡滞后于功能性消亡等因素的制约[12], 由此导致模拟LAI在后期下降幅度过大。另外, 与发育模拟类似, 本研究中GECROS模拟生物量的误差部分可能也归因于玉米品种基因型间的差异及基因型与环境间较大的互作。
当然, 在作物生长模型适应性评价中, 参数的校准方法以及观测数据的质量也可能是影响模型准确率的重要因素。这需要首先对众多作物生长参数进行敏感性分析, 以便明确对模型变量影响最大的参数。然后还要根据参数的生物学意义选取尽可能少的参数开展调试。参数的取值范围也要根据试验观测或前人研究严格确定。对不同来源的观测数据需要进行一致性和质量检查。
GECROS基本能够反映黄淮海地区夏玉米的发育进程。模型模拟夏玉米抽雄期的绝对偏差在6 d以内, 平均为2.1 d; 模拟成熟期的绝对偏差在8 d以内, 平均为3.4 d。GECROS描述夏玉米干物质积累和叶面积扩展过程的准确度较高。模拟雌穗总重的归一化均方根误差在7.8%~31.3%之间, 平均为18.6%; 模拟植株地上总重的归一化均方根误差在11.2%~33.6%之间, 平均为20.7%; 模拟LAI的绝对偏差在0.28~0.55之间, 平均为0.41。模拟籽粒产量的绝对偏差在20.3~229.0 g m-2之间, 平均为80.9 g m-2。利用GECROS模型进行作物生长状况或环境影响条件优劣的相对评价基本可行。但GECROS模拟夏玉米发育进程仍存在低值偏高、高值偏低的现象; 在土壤水分胁迫较重时, 描述的生物量积累过程有偏低情况; 描述LAI扩展的总体效果差于生物量累积的效果。GECROS仍需进一步完善。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|