
新疆小麦 Psy-A1、 Ppo-A1、 Ppo-D1、 TaLox-B1等位变异对面粉色泽的影响
[相吉山1, 2  , 穆培源
, 穆培源1, 2, * , 桑伟1, 2 , 徐红军1, 2, * , 聂迎彬1, 2 , 崔凤娟1, 2 , 庄丽2, 3 , 韩新年1, 2 , 邹波1 ]
, 穆培源, 桑伟, 聂迎彬|
|


第一作者联系方式: E-mail:xiangjsh@163.com
利用已开发的面粉色泽相关性状基因的分子标记, 检测了182份新疆小麦品种(系)的 Psy-A1、 Ppo-A1、 Ppo-D1、 TaLox-B1等位变异, 并结合表型鉴定结果, 分析不同等位变异对面粉色泽的影响。结果表明, 单基因等位变异对面粉色泽的影响不同, 其中4个基因等位变异L*值的差异均不显著; 对a*值的效应, Psy-A1a高于 Psy-A1b( P < 0.01), Ppo-A1b高于 Ppo-A1a( P < 0.05), TaLox-B1a高于 TaLox-B1b( P < 0.05); Psy-A1a基因型的b*值高于 Psy-A1b基因型( P < 0.01); Psy-A1a基因型的Wht值低于 Psy-A1b基因型( P < 0.01)。说明 Psy-A1是影响面粉色泽(红度、黄度和白度)的重要基因, 而 Ppo-A1和 TaLox-B1基因的等位变异只对面粉红度有显著影响。4个检测基因共有12种等位变异组合, 对a*、b*和Wht值有显著效应( P < 0.01), 但对L*值影响不显著( P > 0.05); Psy-A1b/Ppo-A1a/Ppo-D1b/ TaLox-B1b组合具有最高的Wht值和最低的a*、b*值, Psy-A1a/Ppo-A1a/Ppo-D1b/TaLox-B1a组合具有最低的Wht值和最高的a*、b*值。182份品种的Wht值平均为86.86, 其中审定品种平均为86.87, 冬小麦品种为86.42, 春小麦品种为87.32。青春5号具有优良的基因型和较高的面粉白度, 应加强利用。总体来看, 改良新疆小麦面粉的白度, 应重点加强对 Psy-A1基因的选择, 提高 Psy-A1b比例。
, MU Pei-Yuan, SANG Wei, NIE Ying-BinThe allelic variations on Psy-A1, Ppo-A1, Ppo-D1, and TaLox-B1 loci in 182 Xinjiang wheat varieties (lines) were tested using developed molecular markers that are associated with flour color. The effects of these alleles were analyzed in combination with phenotypes with the L*, a*, and b* values as well as the Wht data. On the view of a single gene, the allelic variation showed diverse effects on flour color. For L* value, the allelic effect was not significant. For a* value, the effect was Psy-A1a> Psy-A1b( P < 0.01), Ppo-A1b > Ppo-A1a ( P < 0.05), and TaLox-B1a > TaLox-B1b ( P < 0.05). For b* value, the effect of Psy-A1awas higher than that of Psy-A1b( P < 0.01). As a result, the Wht value of Psy-A1awas signlificantly lower than that of Psy-A1b( P < 0.01). These results suggest that Psy-A1 significantly affects flour redness, yellowness and whiteness, whereas Ppo-A1 and TaLox-B1 only have significant effects on flour redness. A total of 12 allelic combinations were found on the four gene loci, with significant effect on a*, b* and Wht values ( P < 0.01) and slight effect on L* value ( P > 0.05). The combination Psy-A1b/PPO- A1a/PPO-D1b/TaLox-B1b had the highest Wht value and the lowest a* and b* values, and combination Psy-A1a/PPO-A1a/PPO- D1b/TaLox-B1ahad the lowest Wht value and the highest a* and b* values. The average Wht value of total varieties (lines) tested was 86.86, and that of the released, winter, and spring varieties (lines) was 86.87, 86.42, and 87.32, respectively. Qingchun 5 should be used frequently as a promising parent in breeding programs for its excellent genotype and high flour whiteness. Our results indicate that Psy-A1is a key gene for improving flour whiteness, and the frequency of Psy-A1b should be promoted in Xinjiang wheats.
面粉色泽作为评价小麦品质的重要指标, 对面制品的商品性有重要影响[1]。不同面制食品对颜色的要求有一定差异, 东南亚与日本的黄碱面条(yellow alkaline noodle)和硬粒小麦通心面(pasta)要求面条保持亮黄色, 而我国的传统食品如各种白盐面条(white salted noodle)、馒头(steamed bread)、包子(steamed stuffed bun)和饺子(dumpling)等要求色泽亮白[2, 3, 4, 5]。消费市场对面粉色泽多样化的需求, 导致小麦育种目标不同。
面粉的色泽主要由色素决定。色素的总量取决于八氢番茄红素合酶(phytoene synthase, Psy)等色素类物质的积累[6]以及多酚氧化酶(polyphenol oxidase, PPO)、过氧化物酶(peroxidase, POD)、脂肪氧化酶(lipoxygenase, LOX)等对色素类物质的氧化降解[7, 8, 9]。Psy是植物类胡萝卜素合成途径中的限速酶[10], 直接影响籽粒胚乳的颜色[11]。He等[12, 13]克隆了普通小麦7AL染色体上Psy-A1基因的全长序列, 开发了共显性标记YP7A-1和YP7A-2; 结合电子克隆方法, 鉴定并克隆了7BL染色体上Psy-B1的等位变异, 开发了功能标记YP7B-1、YP7B-2、YP7B-3和YP7B-4。PPO是面条制作过程中颜色变褐的主要原因, 其表型贡献率高达50%~70%[14, 15]。张立平等[16]研究表明, 控制PPO活性的主效基因位于2AL和2DL染色体上, 分别解释37.2%~50.1%和25.1%~29.1%的表型变异。Sun等[17]开发了位于2AL上PPO活性主效基因的STS标记PPO18, 能有效地区分控制高、低PPO活性的等位基因Ppo-A1a和Ppo-A1b; He等[18]利用小麦PPO基因序列开发了位于2DL染色体上Ppo-D1a和Ppo-D1b的互补显性标记PPO16和PPO29。LOX可以催化多聚不饱和脂肪酸(polyunsaturated fatty acid, PUFA), 使之释放出高活性的氧自由基, 将面粉及面制品中色素分子的共扼双键氧化为单键, 从而对面粉颜色起到漂白作用[19, 20]。Carrera等[21]认为, 面粉色素的降解主要发生在磨粉和面制品的加工过程中, 并且面制品加工过程中色素的损失主要源于LOX的漂白作用。高LOX活性可以提高面粉和相关产品的白度, 从而提升面粉的商品价值[22, 23]。LOX活性主要受位于第4和第5同源群上的基因调控[24, 25], 第4同源群尤为重要。Geng等[1]利用电子克隆方法克隆了普通小麦4BS和5B染色体上LOX活性基因TaLox-B1和TaLox-B2的全长编码序列, 并开发了互补显性标记LOX16和LOX18, 能有效检测TaLox-B1基因[25]。
新疆拉面(Xinjiang hand-stretched noodle)属于手工鲜食白盐面条, 是新疆独具特色的面条种类。据不完全统计, 新疆小麦面粉年产量的80%都用于制作新疆拉面[26]。新疆拉面对色泽有较高的要求, 在目前的评价标准中, 色泽占新疆拉面加工品质总分的15%[27]。小麦籽粒Psy、PPO、LOX活性及面粉黄色素含量等与颜色相关的指标在小麦育种实践中已受到重视[19, 20, 24], 并已经开发了主效基因的功能标记[13, 17, 18, 25]。王亮等[28, 29]、相吉山等[30]分别对新疆小麦Psy、PPO、LOX相关基因进行了分子标记检测, 但没有探讨这些基因型对面粉色泽的影响。本研究以182份新疆小麦品种(系)为材料, 测定面粉L* 值、a* 值、b* 值, 利用已开发的功能标记对Psy-A1、Ppo-A1、Ppo-D1、TaLox-B1基因进行检测, 分析不同等位变异对面粉色泽的影响, 为利用这些分子标记辅助育种提供参考信息。
182份新疆小麦品种(系), 均由新疆农垦科学院作物研究所小麦课题组收集保存。其中, 冬小麦110份, 包括地方品种7份、引进品种(系) 25份、自育品种(系) 78份; 春小麦72份, 包括地方品种11份、引进品种(系) 20份、自育品种(系) 41份。这些材料涵盖了新疆南北小麦主要种植区各个时期种植和使用的冬、春小麦品种, 具有较好的代表性。
2007— 2008和2008— 2009年度在新疆农垦科学院作物研究所农业试验站种植所有材料, 田间顺序排列; 5行区, 行长1.8 m, 行距0.25 m; 播量为每公顷525万粒。田间管理同大田生产, 正常成熟后及时收获, 人工脱粒。
分年度测定面粉色泽, 取每份材料脱粒后精选的1 kg种子, 在室温存放30 d后制粉, 根据硬度加水润麦, 软质麦、混合麦、硬质麦分别加水至14.0%、15.0%和16.0%, 润麦16~18 h后, 用Quandrmat Senior磨(德国Brabender公司)制粉, 用Minolta CR-310型色彩色差仪(日本Minolta Camera公司)测定面粉色泽。用D65的CIE-L* 、a* 、b* 色度系统表示面粉色泽, L* 值表示白(亮)-黑, 正值表示白色, 其值越大越白(亮); a* 值表示红-绿色, 正值表示红色, 其值越大越红; b* 值表示黄-蓝色, 正值表示黄色, 其值越大越黄。3个月内完成全部测试。
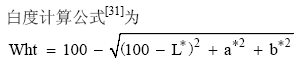
利用SAS 8.0软件对同一基因不同等位变异品种的表型进行成组数据的t测验, 对不同基因型组合
品种的表型进行随机模型的方差分析(ANOVR), 并利用LSD法进行多重比较。
从每个品种(系)选3粒有代表性的种子, 粉碎后放入2.0 mL离心管中, 按照Lagudah等[32]的方法提取基因组DNA。DNA提取液含288 mmol L-1 NaCl、200 mmol L-1 Tris-HCl (pH 8.0)、25 mmol L-1 EDTA和0.5% SDS。
利用已开发的分子标记检测Psy-A1、Ppo-A1、Ppo-D1、TaLox-B1的等位变异(表1)。PCR体系20 μ L, 含20 mmol L-1 Tris-HCl (pH 8.4)、20 mmol L-1 KCl、200 μ mol L-1 dNTPs、1.5 mmol L-1 MgCl2、每条引物10 pmol、Taq DNA聚合酶1.5 U (北京汇天东方)、模板DNA 50 ng。在MJ Research PTC-200 PCR仪(MJ Research, Inc., Waltham, MA)上进行扩增, PCR程序为94℃预变性5 min; 94℃变性45 s, 退火, 72℃延伸1 min, 40个循环; 72℃延伸10 min。
| 表1 检测目标基因等位变异的引物及其扩增信息 Table 1 Marker and amplification information for detecting alleles of target genes |
在1.5%琼脂糖凝胶上电泳分离扩增产物, 缓冲液体系为1× TAE, 电压170 V, 电泳45 min。用溴化乙锭(EB)染色, GelDoc XR System扫描成像并存入计算机。根据3粒种子的检测结果判断每个品种(系) 4个基因的等位变异。当3粒种子带型不一致时, 重新取样和检测, 以2次检测一致的结果为准。
在182份材料中, 4个基因位点各检测到2种等位变异, 均对L* 值无显著效应; 同一基因不同等位变异对a* 值、b* 值、Wht值的影响表现不同, Psy-A1等位变异对三者都有显著影响, Ppo-D1位点对三者均无显著效应, 在Ppo-A1和TaLox-B1b位点, 仅a* 值在等位变异间有显著差异(表2)。说明Psy-A1位点等位变异会引起面粉红度、黄度、白度的极显著变化, 而Ppo-A1、TaLox-B1位点等位变异会引起面粉红度的显著变化。
| 表2 4个基因不同等位变异对面粉色泽的影响 Table 2 Effect of alleles of four genes on flour color |
在182份品种中共检测到Psy-A1、Ppo-A1、Ppo-D1和TaLox-B1基因等位变异组合12种, 其L* 值的差异不显著, a* 值、b* 值、Wht值的差异极显著。其中, 基因型Psy-A1b/Ppo-A1a/Ppo-D1b/TaLox-B1b的a* 值、b* 值均最低, Wht值最高; 基因型Psy-A1a/Ppo-A1a/Ppo-D1b/TaLox-B1a的a* 值、b* 值均最高, Wht值最低(表3)。说明Psy-A1、Ppo-A1、Ppo-D1、TaLox-B1基因等位变异组合会引起面粉红度、黄度、白度的极显著变化。
| 表3 4个基因不同等位变异组合及其色泽差异 Table 3 Flour colors of different allelic combination on Psy-A1, Ppo-A1, Ppo-D1, and TaLox-B1loci |
182份供试材料的Wht值在81.76~91.51之间, 平均为86.86, 说明新疆小麦品种资源具有较高的面粉白度。其中Wht> 90的品种(系)有8份(4.4%), Wht> 85的有166份(91.2%)。在审定品种中, 春小麦品种34个(表4), Wht值平均为87.32, 在4个基因位点上主要等位变异为Psy-A1a(91.2%)、Ppo-A1a(82.4%)、Ppo-D1a(76.5%)和TaLox-B1b (82.4%); 冬小麦品种31个(表4), Wht值平均为86.42, 主要等位变异类型与春小麦相同, 但其比例分别为96.8%、71.0%、93.5%和64.5%。
| 表4 新疆主要审定品种的Wht值及基因型 Table 4 Wht value and genotypes ofmainly released varieties in Xinjiang |
小麦面粉的颜色性状对面条和馒头等食品的外观品质具有重要影响[33], 消费者首先通过面制品的颜色来判断面食品质, 其次才会考虑烹饪后的食味。面粉中黄色素、氧化酶类和蛋白质等均会影响面粉色泽[34, 35]。类胡萝卜素是黄色素的主要成分[14], 主要由叶黄素和黄酮等物质组成[36]。Psy基因在类胡萝卜素家族各种成分的生物合成环节中起着重要作用[6, 37], 是影响小麦籽粒黄色素含量的关键基因[12]。Psy-A1基因位于小麦7AL染色体, 可解释黄色素含量变异的20%~28%, Psy-A1a的黄色素含量显著高于Psy-A1b[13]。本研究发现, 新疆小麦品种中, Psy- A1a的面粉红度和黄度极显著高于Psy-A1b、白度极显著低于Psy-A1b, 说明Psy-A1基因等位变异通过影响黄色素含量[13], 会引起面粉黄度和红度的显著变化, 导致白度发生显著变化。因此, 在要求面粉白度较高的小麦育种中, 应尽量选择含有Psy-A1b基因的材料。
LOX会偶联氧化小麦面粉中的类胡萝卜素进而使小麦面粉变白[38], 其活性是影响面粉黄色素含量的另一主要因素[39], 与磨粉品质及面条、馒头等食品的颜色密切相关[40]。TaLox-B1基因位于小麦4BS染色体, TaLox-B1a的LOX活性极显著高于TaLox-B1b[1, 25]; LOX活性与类胡萝卜素含量呈极显著负相关, 与全麦粉白度呈显著正相关, 与全麦粉黄度相关性不显著[32]。本研究发现, 新疆小麦品种中, TaLox-B1a的面粉红度显著高于TaLox-B1b, 而面粉亮度、黄度、白度的差异均不显著, 这与以往的研究结果[25, 32]略有不同。这与我们所选用的新疆小麦材料有一定的关系, 其原因可能是LOX以酶的形式对色素类物质起催化作用, 具有酶作用的高效性特征。推测LOX活性在某一特定范围内的差异就可以引起黄色素含量或面粉白度的显著变化, 但这一推测还有待验证。
PPO活性与磨粉品质及面条、馒头等食品的颜色密切相关[40], 可以解释面条(团)颜色褐变变异的50%~70%, 是引起面条(团)颜色变褐的主要原因[15]。Ppo-A1基因位于小麦2AL染色体, 可解释PPO活性变异的28%~43%, Ppo-A1a的PPO活性显著高于Ppo-A1b[17]。Ppo-D1基因位于小麦2DL染色体, 可解释PPO活性变异的9.6%~24.4%, Ppo-D1a的PPO活性显著高于Ppo-D1b[18]。本研究发现, Ppo-A1a的面粉红度显著低于Ppo-A1b, 而面粉亮度、黄度、白度的差异均不显著; Ppo-D1a和Ppo-D1b面粉色泽的差异均不显著。可以看出, 虽然Ppo-A1a与Ppo-A1b的PPO活性差异极显著, 但这种差异并不一定能够引起面粉亮度、黄度、白度的显著变化。PPO主要在面粉加工过程中起作用, 是面条制作过程中颜色变褐的主要原因[14, 15, 40], 对面粉色泽(红度除外)的影响并不显著。
使用D65的CIE-L* 、a* 、b* 色度系统表示面粉色泽时, a* 为正值代表红色, 为负值代表绿色[41]。本研究182份材料的a* 值均为负值, 说明新疆小麦面粉色泽偏绿。以往的研究发现, 面粉红度与灰分含量和面粉颗粒度[41]以及籽粒蛋白质含量显著正相关[42], 说明影响面粉红度的因素较多。在本研究中, Psy-A1、Ppo-A1、TaLox-B1基因的等位变异均可导致面粉红度的差异显著, 说明黄色素含量、PPO和LOX活性均能显著影响面粉红度, 进一步印证影响面粉红度的因素较多。
新疆小麦品种资源中, Psy-A1、Ppo-A1、Ppo-D1、TaLox-B1基因共有12种等位变异组合类型, 其面粉亮度无显著差异, 但组合间红度、黄度、白度均有极显著差异, 以Psy-A1b/Ppo-A1a/Ppo-D1b/TaLox- B1b基因型的白度最高、红度和黄度最低, 而Psy-A1a/Ppo-A1a/Ppo-D1b/TaLox-B1a基因型具有最低的白度、最高的红度和黄度。说明这4个基因等位变异对面粉亮度的效应不大, 但对面粉红度、黄度、白度存在极显著影响, 并且红度、黄度的提高会降低白度[32]。在新疆小麦审定品种中, 春小麦品种的面粉白度略高于冬小麦, 其中青春5号的白度最高。据报道, 新疆小麦品种中Psy-A1a为Psy-A1位点的主要等位变异[28, 29, 30]; 在本试验中, 无论冬小麦还是春小麦, Psy-A1a的比例均超过90%, 因此, 降低Psy-A1a的比例是进一步提高新疆小麦面粉白度的重要途径。
Psy-A1、Ppo-A1、Ppo-D1、TaLox-B1基因等位变异对面粉色泽的影响存在较大的区别, 其中Psy-A1对面粉红度、黄度、白度均有极显著影响, 是影响面粉色泽的重要基因; Ppo-A1和TaLox-B1对面粉红度有显著影响。等位变异组合Psy-A1b/Ppo-A1a/ Ppo-D1b/TaLox-B1b是面粉白度的优质基因型, 而Psy-A1a/Ppo-A1a/Ppo-D1b/TaLox-B1a是劣质基因型。由于各基因等位变异对面粉色泽影响程度的不同, 改良新疆地区小麦品种面粉色泽应以利用Psy-A1b为主攻目标, 可利用Psy-A1基因功能标记辅助选择。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|