




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
施氮量对滨海改良盐土棉花钾累积利用的影响
[邹芳刚1, 2  , 张国伟
, 张国伟1 , 王友华1 , 赵文青1 , 周治国1, * ]
, 张国伟]
|
|


第一作者联系方式: E-mail:fg4321@163.com
以长江流域大面积种植的转基因棉湘杂棉8号为材料, 在江苏省大丰市稻麦原种场(33.2°N, 120.5°E)滨海改良盐土上研究施氮量(0、150、300、375、450、600 kg N hm-2)对棉株钾素吸收、利用和分配的影响。结果表明, 增施氮肥提高不同生育阶段棉株钾的吸收量, 以盛花到见絮期的钾积累增量最大, 并改变生育期间的钾吸收比例, 使出苗到盛花期的钾吸收比例降低, 盛花到吐絮期的钾吸收比例升高; 同时, 增加施氮还降低生育后期中上部果枝钾浓度的下降速率, 但对下部果枝影响较小。随施氮量增加, 各部位果枝氮对钾吸收的边际效应(每增施1 kg氮促进钾的吸收量)呈先升高后降低趋势, 且果枝部位越高, 基于最大边际效应的施氮量越高。在300~375 kg hm-2施氮量范围内, 干物质和钾在经济器官中的分配比例提高, 钾浓度和钾累积量动态特征参数比较协调, 中部和上部果枝氮素对钾吸收的边际效应和钾的皮棉生产效率较高, 利于高产形成。高于375 kg hm-2的施氮量导致皮棉产量增幅下降, 氮素对钾吸收的边际效应和钾的皮棉生产效率较低; 低于375 kg hm-2的施氮量降低干物质和钾经济系数, 不利于高产形成。
A field experiment using transgenic cotton cultivar Xiangzamian 8 with different nitrogen application amounts (0, 150, 300, 375, 450, and 600 kg N ha-1) was carried out in the coastal improved saline land region of Dafeng City in 2010 and 2012. Results showed that the nitrogen application increased potassium uptake of cotton at different growth stages, with the highest increment at the peak flowering-boll opening stage, and the percentages of potassium uptake decreased from seedling to peak flowering stages, but increased from peak flowering to boll maturing stages. In addition, nitrogen application lowered the decreasing speed of potassium concentration in middle and upper fruiting branches at later growth stages, but had less influence on lower fruiting branches. The marginal effect of potassium uptake (promoted amount of potassium uptake due to 1 kg increase of N application) showed an upward-downward trend with the increase of nitrogen application rate. The higher the position of fruiting branches grown, the more the nitrogen amount demanded to meet the highest marginal effect of potassium uptake. Under the nitrogen application of 300-375 kg ha-1, the economic coefficient of biomass and potassium was relatively high, eigenvalues of dynamic model of potassium content and potassium accumulation were relatively coordinate, and the marginal effect of potassium uptake and lint production efficiency of potassium were relatively high in middle and upper fruiting branches. Excessively high nitrogen application resulted in a relatively narrow rise of yield, and the marginal effect of potassium uptake and lint production efficiency of potassium were relatively low; excessively low nitrogen application, however, also resulted in a lower yield, as economic coefficient of biomass and potassium was relatively low.
棉花是我国重要的经济作物, 在国民经济中占有重要地位。近年来, 随着粮棉争地矛盾的日益突出, 长江流域下游棉区的棉花种植面积逐渐向土壤条件较为贫瘠的沿海地区集中, 因此, 大力提高滨海盐碱棉区棉花产量对于长江流域下游棉区棉花可持续发展具有重要意义。
长江流域滨海盐土棉区土壤保肥保水能力差, 肥料的增产效应与非盐碱棉田土壤差异较大[1]。目前, 在生产上存在盲目施肥现象, 尤其是氮肥的施用量有不断加大的趋势。大量的研究表明, 过量施入氮肥后, 不仅氮素利用效率降低, 还影响到钾的吸收和利用[2, 3]。与氮不同, 钾在植物体内以K+形态存在, 不被有机化合物同化, 所以钾在植株体内移动性较强。充足的钾供应可以提高光合磷酸化效率, 促进棉株中碳水化合物的合成和运输, 钾营养不足则降低叶片中碳水化合物形成和养分向生殖器官的运输[4, 5, 6], 进而导致棉花早衰和品质变劣[7]。由于钾的上述生理功能, 棉株对钾的吸收、积累和分配特点关系到棉花的产量形成与施肥技术的确定。
棉花的高产以较高的生物量为前提, 而生物量的累积则以养分吸收为基础[5]。前人在棉花上的研究多以氮肥的累积规律为重点, 而较少关注钾的吸收累积规律[8, 9]。娄善伟等[10]研究了新疆地区棉花氮、磷、钾累积量动态及其分配特征的影响, 强调后期生殖器官养分的积累对产量形成具有重要作用。Dong等[11]在黄河三角洲盐土条件下研究表明, 高产棉田每生产100 kg皮棉所吸收的钾量比低产田高20.6%。但是目前关于长江流域沿海棉区氮素对棉株钾吸收, 特别是不同果枝部位果枝钾的吸收的影响研究尚未见报道。
本研究基于长江流域沿海棉区改良盐土条件下棉花种植过程中的养分运筹习惯, 探讨施氮量对滨海改良盐土棉花钾累积利用的影响, 以期为制定滨海盐土棉区合理的棉田养分管理技术体系提供理论依据。
试验于2010年和2012年在江苏省大丰市稻麦原种场(33.2° N, 120.5° E)滨海改良盐土进行, 供试土壤为潮盐土、沙性轻壤, 2年0~20 cm土壤pH值分别为7.9和7.8, 含全盐0.29%和0.26%、有机质17.9 g kg-1和18.3 g kg-1、全氮1.02 g kg-1和1.09 g kg-1、速效氮61.5 mg kg-1和60.2 mg kg-1、速效磷42.5 mg kg-1和43.3 mg kg-1、速效钾180.0 mg kg-1和182.4 mg kg-1。
以长江流域大面积种植的转基因棉湘杂棉8号为材料, 设置60、150、300、375、450、600 kg hm-2 6个施氮量水平, 重复4次, 随机区组排列。总施氮量中, 基肥、初花肥和盛花肥分别占30%、40%和30%。P2O5、K2O用量分别为150、225 kg hm-2, 全部作为基肥。小区长10 m, 宽8 m, 8行区, 行距1.0 m, 株距为0.4 m, 密度约25 000株hm-2。4月5日左右播种育苗, 5月20日左右移栽, 8月10日左右打顶。精细整枝, 其他管理同高产田块。
分别于6/29、7/15、7/30、8/15、8/30、9/15、10/5在每小区取生长发育一致的2株, 3次重复, 将棉株分为下(1~5果枝, L)、中(6~10果枝, M)、上(≥ 11果枝, U)三部位调查生物量与养分空间分布, 在105℃杀青30 min后, 80℃烘至恒重, 称生物量。样品粉碎后用原子吸收分光光度法测定各部位钾含量[12], 再根据各部位的干物质重计算棉花单株钾素累积量。收获时对每小区定点的20株棉花按照不同部位分收棉铃, 测定产量。
钾的皮棉生产效率(kg kg-1) = 皮棉产量/吐絮期整株钾积累量
生物量经济系数 = 吐絮期生殖器官生物量/吐絮期整株生物量
钾的经济系数 = 吐絮期生殖器官钾累积量/吐絮期整株钾积累量
钾的吸收比例(%) = 不同生育阶段棉株钾吸收量/吐絮期整株钾积累量
棉株钾素累积量的增长符合Logistic曲线, 其基本模型为W=Wm/(1+aebt), 分别对模型求1阶、2阶和3阶导数, 可得相应生长曲线的最大相对生长速率(Vmax)及其出现时间(tmax), 快速累积期持续时间(T)等特征参数。
采用Microsoft Excel软件处理数据, 用SPSS 11.0软件分析方差和测验差异显著性。
由图1可见, 随施氮量增加, 总皮棉产量及上、中和下部皮棉产量均呈先上升后降低的趋势。用二次曲线拟合后可知, 2010年和2012年总皮棉产量、上部皮棉产量、中部皮棉产量和下部皮棉产量分别在397.6 kg N hm-2和379.7 kg N hm-2、456.5 kg N hm-2和419.2 kg N hm-2、417.6 kg N hm-2和424.8 kg N hm-2、276.4 kg N hm-2和246.9 kg N hm-2时最高。
 | 图1 施氮量与皮棉产量的关系TP: 整株; L: 下部果枝; M: 中部果枝; U: 上部果枝。Fig. 1 Relationship between nitrogen amount and lint yieldTP: total plant; L: lower fruiting branches; M: middle fruiting branches; U: upper fruiting branches. |
由表1可见, 施氮量显著影响棉花生物量累积分配。对于棉株生物量, 整株及中部果枝生物量在450 kg N hm-2(2010)和375 kg N hm-2(2012)处理下最高, 下部果枝在375 kg N hm-2处理下最高, 上部果枝在450 kg N hm-2处理下最高, 且达到峰值后随施氮量的增加各部位生物量降低不显著(P> 0.05)。对于生殖器官生物量, 整株、中部和下部果枝在375 kg N hm-2(2010)和300 kg N hm-2(2012)处理下最高, 上部果枝则在450 kg N hm-2处理下最高。对于生物量经济系数, 下部果枝表现为随施氮量的增加而降低, 中部果枝呈先升高后降低的趋势, 并在375 kg N hm-2(2010)和300 kg N hm-2(2012)处理下最高, 上部果枝则随施氮量的增加而增大。
| 表1 施氮量对生物量累积和分配的影响 Table 1 Effects of nitrogen amounts on biomass accumulation and distribution of cotton plant |
由表2可见, 随施氮量的增加, 下部果枝钾累积量呈先升高后降低的趋势, 并在450 kg N hm-2(2010)和375 kg N hm-2(2012)处理下下最高; 棉株整株、中部及上部果枝钾累积量则呈升高趋势, 并在升高到一定量后趋于稳定。对于生殖器官钾累积量, 随施氮量的增加, 下部和中部果枝呈先升高后降低的趋势, 分别在300 kg N hm-2和375 kg N hm-2处理下最高, 而整株和上部果枝则持续升高, 且在升高到一定量后趋于稳定。对于钾的经济系数, 随施氮量的增加, 下部和中部果枝呈先升高后降低趋势, 分别在150 kg N hm-2和300 kg N hm-2处理下最高, 上部则表现为升高到一定量后趋于稳定。
| 表2 施氮量对钾累积分配的影响 Table 2 Effects of nitrogen amounts on potassium accumulation and distribution of cotton plant |
由图2可见, 不同施氮量下棉株各部位的钾浓度均随生育进程逐渐降低, 变化范围为1.55%~ 3.79%。回归分析表明, 不同果枝部位钾浓度随时间的变化趋势均可用负指数函数方程(y = ae-bx)拟合(表3)。分析表3可知, 随施氮量增加, 各果枝部位钾浓度衰减方程的a值(初始值)均持续升高, 即钾浓度升高, b值(下降速率)变化相对复杂, 对于下部果枝, 施氮对b值影响较小, 对于中部和上部果枝, 低于150 kg N hm-2处理对b值影响较小, 高于此浓度的氮肥处理则降低了b值。
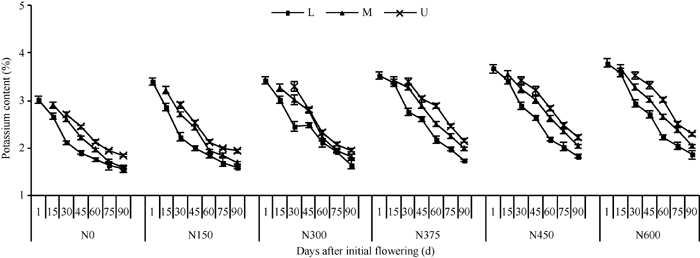 | 图2 施氮量对棉花不同部位钾浓度的影响(2010年)L: 下部果枝; M: 中部果枝; U: 上部果枝。Fig. 2 Effects of nitrogen amounts on potassium content of cotton plant in 2010L: lower fruiting branches; M: middle fruiting branches; U: upper fruiting branches. |
| 表3 不同氮素处理下棉花不同果枝部位钾浓度拟合方程参数(2010年) Table 3 Parameters of equations for potassium content of different fruiting branches in cotton under different nitrogen amounts in 2010 |
随着生育进程, 棉株各部位钾累积量的变化符合Logistic生长曲线(图3)。对图3数据拟合分析可得到钾累积动态模型的特征值(表4), 棉株各部位钾最大累积速率和最大累积速率出现时间变化一致, 下部果枝以375 kg N hm-2施氮量最高, 中部和上部果枝则随施氮量的增加而增加; 下部果枝钾快速累积期持续时间随施氮量的增加而延长, 中部果枝以300 kg N hm-2施氮量最短, 上部果枝则随施氮量的增加而缩短。
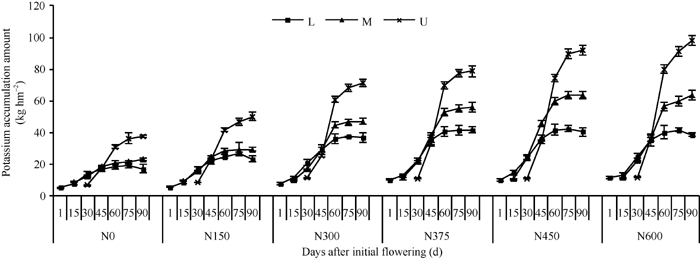 | 图3 施氮量对棉株不同部位钾累积量的影响(2010年)L: 下部果枝; M: 中部果枝; U: 上部果枝。Fig. 3 Effects of nitrogen amount on K accumulation amounts of cotton plant in 2010L: lower fruiting branches; M: middle fruiting branches; U: upper fruiting branches. |
| 表4 钾累积动态特征值(2010年) Table 4 Eigenvalues of potassium accumulation models in different parts of cotton plant in 2010 |
分析表5可知, 下部果枝生物量和钾的经济系数与钾含量回归方程a值(初始值)和钾快速累积期持续时间呈显著负相关, 中部果枝生物量和钾的经济系数仅与钾快速累积期持续时间显著正相关, 上部果枝生物量和钾经济系数与钾含量回归方程a值和最大累积速率显著正相关、与钾含量回归方程b值(下降速率)和快速累积期持续时间显著负相关。
| 表5 不同部位生物量和钾经济系数与钾累积特征参数的关系(2010年) Table 5 Correlation coefficients of economic coefficient of biomass and potassium with eigenvalue of potassium accumulation in cotton in 2010 |
分析表6可知, 不同生育时期间钾的吸收比例以盛花到见絮最高, 初花到盛花次之, 出苗到初花和吐絮期最低。随着施氮量的升高, 钾吸收量在吐絮期逐渐增加, 其余时期均呈先升高后降低趋势, 且均在450 kg N hm-2时最高; 钾吸收比例变化规律差异较大, 在出苗到初花期和初花到盛花期呈降低趋势, 在盛花到见絮期表现为升高到一定程度后趋于稳定, 在吐絮期则表现为升高趋势。
| 表6 施氮对棉株不同生育阶段钾吸收量和吸收比例的影响 Table 6 Effect of nitrogen amount on potassium uptake and percentages of uptake at growth stages in cotton |
随施氮量增加, 棉株钾吸收量变化规律可用Logistic方程拟合[13], 对表2中棉株不同部位钾吸收量随施氮量的变化采用Logistic曲线拟合, 拟合方程如图4。通过对方程求导, 可以得出氮对钾吸收的边际效应(每增施1 kg氮促进钾的吸收量)。可以看出, 随着施氮量的增加, 钾吸收的边际效应呈先升高后降低趋势, 上、中、下部果枝和整株钾吸收的边际效应分别在363.3、210.5、88.0和146.9 kg N hm-2时达到最大。
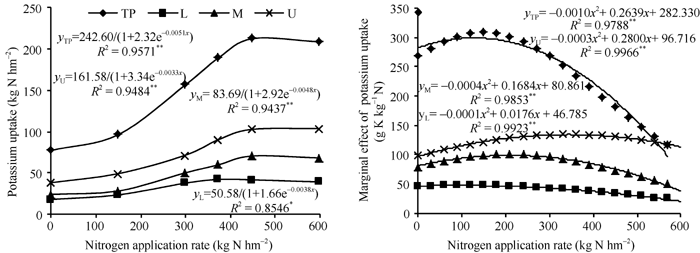 | 图4 施氮量与钾吸收量和钾吸收边际效应的关系(2010年)TP: 整株; L: 下部果枝; M: 中部果枝; U: 上部果枝。Fig. 4 Relationship between nitrogen amount and accumulation and marginal effect of potassium in 2010TP: total plant; L: lower fruiting branches; M: middle fruiting branches; U: upper fruiting branches. |
皮棉产量对钾吸收量的响应可用二次曲线拟合(图5), 上、中、下部果枝和整株达到最高皮棉产量时的理论吸钾量分别为81.8、52.9、29.8和162.1 kg K hm-2。对二次方程求导可得到钾的皮棉生产效率(每吸收1 kg钾可生产的皮棉产量), 随钾吸收量的升高, 上、中和下部果枝及整株的钾的皮棉生产效率均呈线性降低, 其降低趋势表现为下部果枝最大, 中部果枝次之, 上部果枝最低。
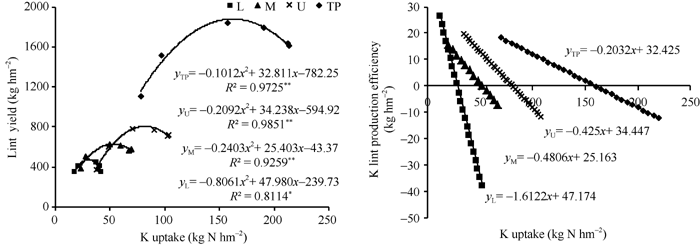 | 图5 钾吸收量与皮棉产量及钾皮棉生产效率的关系(2010年)TP: 整株; L: 下部果枝; M: 中部果枝; U: 上部果枝。Fig. 5 Relationship between potassium accumulation and lint yield and lint production efficiency of potassium in 2010TP: total plant; L: lower fruiting branches; M: middle fruiting branches; U: upper fruiting branches. |
本研究中随施氮量升高, 总皮棉产量及上、中和下部皮棉产量均呈先上升后降低趋势, 在达到理论最大值时, 整株需氮量(370~400 kg N hm-2)低于中部和上部需氮量(400~430 kg N hm-2和410~460 kg N hm-2), 高于下部需氮量(240~280 kg N hm-2), 表明在滨海盐碱改良土上, 基于产量最高时的施氮量促进了中、上部果枝产量形成, 却限制了下部果枝产量形成。这可能与高氮处理下棉花群体叶面积过大、冠层通风透光差、下部荫蔽影响下部果枝光合作用有关[14]。同时, 薛晓萍等[15, 16]研究认为, 长江下游棉区适宜的施氮量水平是240 kg N hm-2, 远低于本研究的结果, 其原因可能是本试验地区为沙性土壤, 保肥能力较差, 养分流失较重, 并且已脱盐改良的土壤含盐量虽较低, 但其中盐离子可能仍影响棉株对养分的吸收, 鲁雪林等[17]的研究则表明冀东滨海盐碱地的适宜施氮量为337.5 kg N hm-2, 远高于非盐碱棉区, 这与本研究结果相似。
棉花具有无限生长习性, 受所处环境条件和自身生理年龄的影响, 施氮量对不同部位钾累积及向生殖器官转运的影响存在差异, 进而影响不同部位生长发育和产量器官形成[18, 19]。用于生殖器官发育并形成碳水化合物的钾的减少是导致棉花早衰和品质变劣的主要因素之一[7]。本研究表明, 尽管棉株各部位及其生殖器官钾吸收量变化趋势一致, 但是钾的经济系数变化差异较大。对于下部和中部果枝, 钾的经济系数随施氮量的增加呈先上升后降低趋势, 分别在150和300 kg N hm-2处理下达到最大, 而上部果枝则表现为升高到一定量后趋于稳定, 这说明低于150和300 kg N hm-2处理分别促进了钾向下部和中部生殖器官分配, 高于此水平则倾向于向营养器官分配, 导致下部和中部营养生长过旺, 生殖器官生物量和产量降低; 对于上部果枝, 施氮则促进了钾向生殖器官分配, 这与产量和生物量变化趋势基本一致, 也导致上部果枝产量占总产比例的增加。
植物体内钾参与众多生理代谢过程, 棉株中钾浓度的变化与碳水化合物的形成和养分向生殖器官的运输密切相关[20]。本研究表明, 随生育进程, 棉株各部位的钾浓度变化可用负指数函数方程拟合, 对于下部果枝, 钾浓度衰减方程a值(初始值)与生物量和钾经济系数显著负相关, 对于中部果枝, 钾浓度的变化对生物量和钾的生殖分配影响不显著; 对于上部果枝, 生物量的生殖分配与初始钾含量(a值)显著正相关, 而与钾含量下降速率(b值)显著负相关, 说明钾含量的变化过程可以通过影响下部和上部生物量和钾的生殖分配而影响棉花产量。进一步分析表明, 随着棉花果枝部位的上升, 钾吸收峰值出现时间推迟, 钾累积速率峰值增加, 快速累积期持续时间缩短, 说明果枝部位越高, 其钾的吸收越集中。对于下部和中部果枝, 较短的快速累积期持续时间有利于光合产物和钾向生殖器官转运; 对于上部果枝, 较高的最大累积速率、较迟的最大累积速率出现时间和较短的快速累积持续时间利于光合产物和钾向生殖器官的转运。
棉株钾的吸收随施氮量的动态变化可用Logistic曲线描述[13], 对其求导可以得到氮对钾吸收的边际效应。本研究中, 随施氮量增加, 各部位氮对钾吸收的边际效应均呈先升高后降低趋势, 且果枝部位越高, 基于氮对钾吸收的最大边际效应的施氮量越高。通过比较基于皮棉产量理论最大值时施氮量(379.7 kg N hm-2)和基于上、中和下部氮对钾吸收的最大边际效应的施氮量(363.3、210.5和88.0 kg N hm-2)可知, 滨海改良盐碱土高产的形成与上部果枝氮对钾吸收的边际效应关系最为密切。
棉花生育前期吸收的钾主要是促进棉株营养器官发育, 后期吸收的钾则对产量形成至关重要[21]。施氮量过低增加了盛花期之前钾的吸收比例, 降低了盛花期之后的钾吸收比例, 导致后期钾供应不足; 过量施氮虽然增加了棉花吐絮期钾的吸收比例, 使吐絮期钾的浓度和吸收量显著增加, 但是过高钾浓度反馈促进了氮的吸收[22], 导致棉花贪青晚熟, 品质下降, 这可能是过量施氮导致棉花产量降低的原因之一。
施氮量通过调控棉花不同果枝部位钾含量和钾累积量的动态影响钾和生物量的累积和转运, 进而影响棉花产量。在300~375 kg hm-2施氮量范围内, 干物质和钾在生殖器官中的分配比例提高, 钾浓度和钾累积量动态特征参数比较协调, 中部和上部果枝氮素对钾吸收的边际效应和钾的皮棉生产效率较高, 利于高产形成。高于375 kg hm-2的施氮量导致皮棉产量增幅下降, 氮素对钾吸收的边际效应和钾的皮棉生产效率较低; 低于375 kg hm-2的施氮量降低干物质和钾经济系数, 不利于高产形成。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|