

{kind=link}
{kind=link}
利用非条件和条件QTL解析油菜产量相关性状的遗传关系
[焦聪聪1, 2  , 黄吉祥
, 黄吉祥2 , 汪义龙3 , 张晓玉4, 2 , 熊化鑫1, 2 , 倪西源2 , 赵坚义2, * ]
, 黄吉祥]
|
|


第一作者联系方式: E-mail: jiaodacong@126.com, Tel: 15958014015
基于前期研究中构建的Sollux/Gaoyou DH群体在9个环境中的表型数据和新版遗传图谱, 对油菜角果长度进行QTL定位, 估测QTL的加性、上位性和环境互作效应。并通过条件QTL方法, 解析角果长度与角果粒数和粒重之间的遗传关系, 以期利用标记辅助, 探讨通过选择角果长度基因型以增加角果粒数、提高千粒重, 最终达到增加产量的可能性。结果共检测到在3个环境以上稳定表达的控制角果长度QTL 8个, 加性效应值在0.09~0.26 cm之间, 效应总和解释群体遗传总变异的60%。8对上位性QTL效应值在0.035~0.075 cm之间, 效应总和为加性总效应的38%。QTL与环境互作效应只在少数位点和个别环境中显著。条件QTL研究表明, qSLA2、 qSLC1-2和 qSLC8-1位点, 角果长度的变化对角果粒数影响较大; 而通过选择 qSLA7、 qSLC1-2、 qSLC8-1和 qSLC8-2长角果标记基因型, 可望同时提高角果粒数和千粒重。6个主效QTL 11个连锁标记基因型和表现型的关联分析, 验证了条件QTL分析结果, 表明通过对 qSLA2、 qSLA7、 qSLC8-1和 qSLC8-2位点6个连锁标记( ZAAS423、 SUC1-3、 ZAAS12a、 ZAASA7-28、 ZAAS433和 ZAAS437)长角果基因型的聚合, 可增长角果约2 cm, 间接增加角果粒数2粒, 同时提高千粒重0.4 g, 从而可望实质性地提高油菜产量水平。
Quantitative Trait Loci (QTLs) for silique length (SL) were mapped in the updated SG map using the phenotypic data from nine environments. QTLs with additive and epistatic effects and their interactions with environments were estimated. At QTL level, conditional QTL analysis was performed to dissect the genetic relationships between silique length and seed number per silique (SS), and between silique length and 1000-seed weight (SW). Our goal was to identify QTLs that are important for silique length, as indexed by their positive correlations with either seeds per silique or 1000-seed weight, or both of the traits. Markers linked to the target QTL can be developed for indirect selection of SS and SW. As shown by the results, we detected eight QTLs with additive effects, which together accounted for around 60% of the phenotypic variations. While the total effects of eight pairs of epistatic loci (additive × additive) ranged from 0.035 to 0.075 cm and their summation was 38% of the total additive effects. QTL by environmental interactions were significant only in few environments with small amount of genetic effects. The conditional QTL analysis revealed large impact of silique length on seed number per silique in three QTLs ( qSLA2, qSLC1-2, and qSLC8-1). Allelic selection for long silique length in qSLA7, qSLC1-2, qSLC8-1, and qSLC8-2loci could potentially increase the seed number per silique (SS) and 1000-seed weight (SW). Association analysis between genotypes linking to six related QTLs and the corresponding phenotypes of yield related traits indicated that the combination of long silique alleles from four QTLs ( qSLA2, qSLA7, qSLC8-1, and qSLC8-2) by marker assistant selection of ZAAS423, SUC1-3, ZAAS12a, ZAASA7-28, ZAAS433, and ZAAS437 significantly increased about two cm in silique length. Meanwhile, two additional seeds per silique were increased, and the 1000-seed weight was enhanced by 0.4 g. Taken together, we suggest the importance of these QTLs and markers for yield breeding purpose in Brassica napus.
油菜是我国最重要的食用植物油来源之一, 约占总量的50%。近年来, 随着人口数量的增长和土地使用面积的减少, 我国油菜籽的生产水平远低于百姓对菜籽油的实际需求, 当前约60%的植物食用油依赖进口, 加上近年来我国油菜单产在经历了持续上升阶段后已出现徘徊局面。因而如何提升育种技术, 进一步提高油菜作物单产水平成为育种研究者面临的重大难题。
油菜产量由单位面积角果数、角果粒数和粒重三因子构成。其中, 粒重与角果粒数较单株角果数具较高的遗传力, 受环境影响较小, 育种改良潜力相对较大[1, 2], 有研究表明选育长角果品种可望提高油菜单产水平[3, 4]。角果长度便于观察记载, 作为一种简单实用的形态指标常用来间接选择角果粒数和千粒重, 因为众多研究表明角果长度和角果粒数呈极显著正相关[5, 6], 与千粒重亦多为同向关系。Yang等[7]发现油菜A9 (uqA9)和A6 (uqA6)连锁群上的主效QTL同时控制角果长度和粒重, 效应方向一致; Li等[8]利用连锁和关联性分析, 再次证明A9和C2上控制角果长度主效QTL uqA9-1、uqA9-2和uqC02-1同时控制千粒重, 揭示了二性状显著正相关的遗传基础。其中uqA09-1解释千粒重和角果长度的遗传变异分别为20.1%和19.0%, uqA09-3分别达13.5%和13.2%。Cai等[5]结果也显示角果长度和千粒重正相关, 但也有研究表明二者呈显著负相关[6]。油菜角果长度是一个典型的数量性状, 受多基因控制, 存在基因型与环境以及基因间相互作用[5, 6]。经典遗传学认为, 油菜的角果长度主要受2个互补的主效基因控制, 同时可能受一些微效基因的影响[3]。基因以加性效应为主, 遗传力约68%[9]。随着分子标记技术的发展和QTL (quantitative trait lous)定位理论的提出, 前人对油菜角果长度QTL的研究已有不少报道。在甘蓝型油菜中, Udall等[10]在A9、C2和C8连锁群上检测到控制角果长度QTL, 其中A9和C2连锁群上的主效QTL贡献率超过10%。Chen等[11]在A10、C1、C2、C5和C7连锁群上定位到角果长度QTL, 其中C2和C7上的2个QTL分别解释高达17.1%和16.8%的遗传变异。Zhang等[6]在A3、A4、A7、A8、C2、C3、C4和C6上检出QTL, 其中A3和A8上的位点贡献率超过10%。Yang等[7]在不同群体和不同环境中对角果长度进行QTL分析, 发现A6、A7、A9、C2和C3上的QTL在多个环境和多个遗传群体中存在。Li等[12]研究发现, A2、A3、A7、A8、A9和A10上存在主效QTL, 其中贡献率最大的位于A7连锁群, 解释19%遗传变异。综合前人研究结果, 控制角果长度的主效QTL高频率地出现在A7、A9、C2和C3连锁群。对于油菜角果长度虽然已有不少研究报道, 但如何在QTL层面上解析角果长度与角果粒数和千粒重之间的关系, 通过选择角果长度达到同步改良粒数和粒重的可能性尚有较大研究空间。
本研究利用前期工作中培育的SG-DH群体[13], 在中国杭州、西安以及德国哥廷根共9个环境下的田间试验结果和新版SG图谱[14], 对角果长度、角果粒数和千粒重3个重要产量相关性状进行QTL分析。在此基础上, 引进条件遗传分析方法, 分别以群体内株系间角果粒数或千粒重恒定不变为条件时重新定位角果长度QTL, 分析比较条件和非条件QTL定位结果, 揭示角果长度与角果粒数和千粒重在不同QTL的遗传关系, 探讨通过选择角果长度基因型同时改良角果粒数和千粒重, 最终达到提高产量水平的可能性。
以欧洲高含油量冬油菜Sollux和中国品种高油605选系Gaoyou杂交F1衍生的282个DH (doubled haploid)系为试验材料[13]。QTL定位图谱包含481个分子标记, 覆盖甘蓝型油菜基因组19条连锁群, 总长1948.6 cM, 标记间平均距离4.05 cM[14]。
于2000— 2001在4个试验点(中国杭州、西安以及德国哥廷根的Reinshof和Weende)及2005、2007、2008、2009和2013年度在杭州(浙江省农业科学院试验农场)共9个环境下试验。试验采用随机区组设计, 2次重复。每株系种植2行, 行长2.5~3.0 m, 行距0.33 m, 株距0.12 m。
油菜成熟时, 从每小区(株系)随机选取正常生长植株5株, 取主花序中部10个长势整齐的角果, 测量其总长度(测量范围不包括角果喙长)和角果粒数, 所得的平均值视为该株系角果长度和角果粒数表型值, 小区混收种子用于千粒重测定。
采用SPSS17.0软件统计DH群体性状表型及性状间相关系数。采用Zhu[15]提出的基于混合线性模型的数量性状条件分析方法获取条件效应值, yT1|T2表示在消除性状2的表型变异后性状1的表型值, 如ySL|SW指剔除千粒重(SW)影响后的角果长度表型值。
首先运用WinQTLCart 2.5复合区间作图法[16], 对角果长度表型值按单环境在全基因组进行QTL扫描, 采用LOD> 2.5作为阀值判断QTL是否真实存在。在每个连锁群上间隔1 cM检测QTL存在的可能性, 确定各性状QTL的数目及连锁图上QTL的置信区间、加性效应值以及对性状表型的贡献率。
进一步采用QTLNetwork 2.0软件[17], 对9个环境下的表型数据进行联合定位分析, QTL的显著性概率值为0.05, 通过1000次排列, 检验推断QTL及成对推断QTL的效应显著性, 用检验出的显著QTL构建QTL全模型, 进一步对模型进行选择, 剔除可能的假阳性QTL。用基于Gibbs抽样的Bayesian方法[18], 着重分析控制角果长度的上位性QTL以及QTL与环境的互作效应。
采用9个环境下角果长度表现型的平均值按1.4方法构建分别剔除角果粒数和千粒重影响后的角果长度表型值, 用WinQTLCart 2.5复合区间作图法[16]对角果长度进行条件QTL分析。
采用q+角果长度英文缩写SL+所在连锁群代号+QTL个数命名QTL。如对油菜第18染色体上第1个角果长度QTL, 命名为qSLC8-1。
表1表明, 2个亲本的角果长度平均值相差仅0.1 cm, 但DH群体株系间角果长度最大值达7.2 cm, 最小值约3.9 cm, 极差在3.3 cm, 出现明显的双向超亲现象, 说明增、减角果长度等位基因分散在双亲之间。长、短角果等位基因的重组和聚合使得群体中出现角果长度达到7.0 cm以上或短于3.5 cm以下株系。另值得注意的是在德国环境中, 欧洲亲本Sollux平均角果长度较中国亲本Gaoyou长1.5 cm, 但在中国杭州环境下, 则是中国亲本Gaoyou较Sollux平均长0.4 cm, 说明环境因素对油菜角果发育和最终长度有一定影响。
角果长度与角果粒数或千粒重之间均存在极显著正相关, 相关系数分别为0.231和0.188 (P< 0.01), 说明通过对角果长度的选择有可能同时增加角果粒数和千粒重, 但角果粒数和千粒重之间极显著负相关(-0.277, P< 0.01), 与许多前人研究结果相符。
利用WinQTLCart 2.5软件对9个环境下的角果长度进行QTL扫描, 共检测到在3个以上环境中稳定表达的QTL 8个(表2), 分别是qSLA2、qSLA4、qSLA7、qSLC1-1、qSLC1-2、qSLC2、qSLC8-1和qSLC8-2, 分布在6条相应的染色体上, 加性效应值在0.09~0.26 cm之间。当所有QTL位点纯合并存时, 角果长度可相应增长2.60 cm, 解释群体内遗传总变异的60%, 单个QTL平均贡献率在3.7%~10.5%。其中, qSLA2、qSLC2和C8上的2个主效QTL分别在5个或以上环境中稳定表达, 加性效应平均值为0.18~ 0.19 cm (图1)。相距20~30 cM的2个QTL qSLC8-1和qSLC8-2贡献率最大, 超过10.0%; qSLA2、qSLC1-1、qSLC1-2和qSLC2 4个QTL的增效基因来源于亲本Sollux, 而qSLA4、qSLA7、qSLC8-1和qSLC8-2增效基因来源于亲本Gaoyou。
| 表1 SG-DH群体在9个环境中角果长度的表型变异 Table 1 Phenotypic variation of silique length in SG-DH population across nine environments |
| 表2 SG-DH群体在9个环境中检测到控制角果长度QTL Table 2 QTL information of silique length in SG population over nine environments |
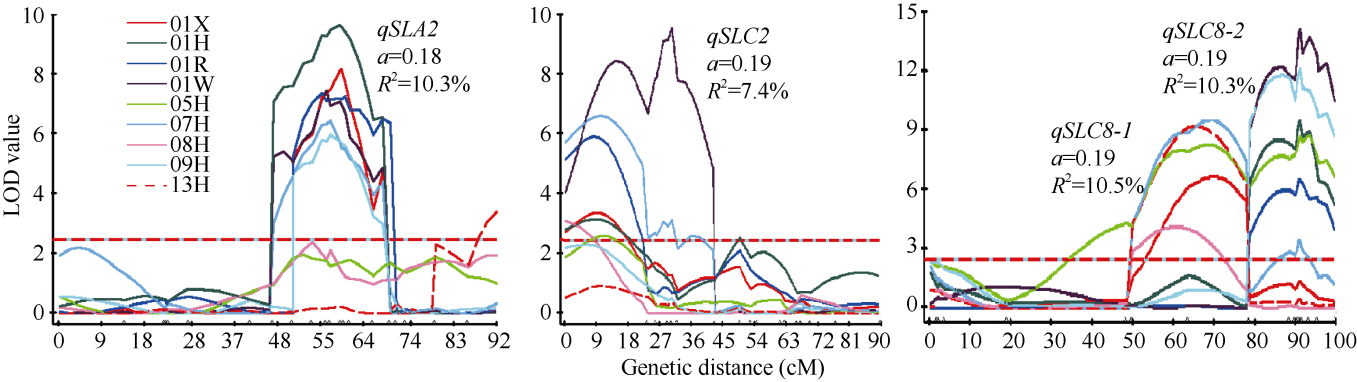 | 图1 SG-DH群体中4个控制角果长度主效QTL在连锁群上的位置Fig. 1 Scanning results of four major QTLs for silique length over nine experiments in SG DH population |
为解析角果长度QTL与环境以及QTL间的相互作用, 进一步采用QTLnetwork 2.0软件对9个环境下的角果长度进行QTL联合分析, 共检测到8对上位性QTL。其中2个主效QTL qSLC2和qSLC8-2分别与A1和A3上的标记位点ZAASA1-73和ZAAS613互作, 其余6对均发生在2个标记位点之间(表4)。所有上位性QTL的效应值在0.035~0.075 cm之间。当所有位点纯合并存时, 增减角果长度的效应值达到0.87 cm, 是加性总效应的38%左右。8对上位性QTL, 除ZAAS202/ DGD2_1为C× C基因组间互作, 其余7对均发生在A和C之间。角果长度QTL与环境互作(QE)只发生在2个QTL位点, 效应值很小。qSLA2在2001西安和2005杭州环境中存在显著的QE互作, 效应值在0.04~0.05 cm。qSLC2在2001Weende, 2008和2009杭州三个环境中被检测到QE互作, 互作效应在0.05~0.11 cm之间, 未检测到其余6个QTL显著的QE互作。所有上位性QTL均未被检测到有显著的环境互作效应。
| 表3 SG-DH群体中角果长度的上位性效应 Table 3 Epistatic interactions estimated for silique length in SG-DH population |
以9个环境角果长度的平均值和剔除千粒重或角果粒数变异后的条件平均值进行条件前、后的QTL分析和比较(表4), 结果显示5个角果长度QTL, 角果粒数和千粒重的变化对其有不同程度的影响。当排除角果粒数和千粒重因素后, qSLA2的效应值略有下降但幅度不大, qSLC8-2位点则分别下降约30%和50%, 贡献率从12.8%降至4.6%和2.5%。在qSLA7、qSLC1-2和qSLC8-1位点, 角果长度对角果粒数和千粒重的影响是另一种情况, 在假定角果粒数不变情况下(SL|SS), qSLA7基本无影响, qSLC1-2和qSLC8-1效应值和贡献率略有增加。但给定千粒重不变时(SL|SW), 这3个QTL的效应值均较大幅度提高(qSLA7从0.08至0.13, qSLC1-2从0.13至0.17, qSLC8-1从0.09至0.16), 增幅分别达63%、31%和78%, qSLC2条件前、后效应值基本无变化; 除此, 千粒重被条件后, 涉及6条连锁群的7个位置上出现新的角果粒数QTL, 但效应值均较小, 在0.08~ 0.12 cm之间, 角果粒数被条件后, 只在qSLC7位点检测到一个新的微效角果长度QTL。条件QTL结果显示角果长度与千粒重的关系似大于和角果粒数的关系。
| 表4 角果长度条件和非条件QTL的加性效应和贡献率 Table 4 Additive effect and genetic contribution of conditional and unconditional QTLs for silique length |
为验证主效角果长度QTL的遗传效应, 探讨通过标记辅助选择和聚合长角等位基因, 间接获得多粒大粒材料的可能性, 从而筛选高效实用分子标记应用于育种。我们从SG群体282个株系中选取角果最长和最短的株系各20个, 用与定位在5条连锁群上的6个主效QTL (qSLA2、qSLA7、qSLC1-1、qSLC2、qSLC8-1和qSLC8-2)连锁的11个标记进行标记基因型和性状表现型的关联分析(表5)。在平均角果长度> 6.3 cm的20个株系中, 除qSLA7外, 其他5个位点的9个连锁标记携长果等位基因的比例在75%以上。qSLC1-1和qSLC22个QTL, 虽然长角果组中, 4个标记的符合率均高于95%, 但在平均角果长度< 4.54 cm的短果组中, 含长果等位基因的比例也高达45%~ 70%; 而与qSLA2、qSLC8-1和qSLC8-2 3位点连锁的5个标记, 在长、短2组中的符合概率均在75%以上, 适合于育种应用。另长角果组的20个株系, 平均角果粒数较短角果组多2粒, 千粒重高约0.4 g, 差异均达极显著(P< 0.001)。表明通过对长角果等位基因的标记筛选可望获得角果粒数多且千粒重高的育种材料。
| 表5 SG群体极端株系6个主效QTL标记基因型和角果长度表现型的关联分析 Table 5 Marker defined genotypes of individuals with extreme phenotypes for silique length in SG population |
本研究利用SG群体9个环境下的油菜角果长度表型变异, 共检测到在3个以上环境中效应显著QTL 8个。其中, qSLA2、qSLC1、qSLC8-1和qSLC8-2 4个主效QTL在5个或以上环境下稳定表达, 单个贡献率大于7.8%。对油菜角果长度QTL的研究, 前人已有众多报道, 19条连锁群中, 除A1、A5和C9外, 其余16条上均曾检测到控制角果长度QTL [6, 7, 10, 11, 19]。因实验材料和试验环境不同, 定位结果不尽一致。Zhou等[20]通过对3个DH和1个RIL群体角果长度定位结果的整合分析, 提出A1、A9和C8上各存在1个共性QTL, C2和C3上各有4个和3个共性QTL。我们将本研究结果与前人结果比较(图2), 初步判断qSLC2、SL12[7]和qSLC2-1a[11]; qSLA7、qSLA7[11]和qSLN7-1[6]位于大致相同基因组区域; qSLC2峰值位置在约9.0 cM附近, 相应物理距离与Zhou等[20]报道的约9.2 Mb位置比较一致, 很可能是相同QTL。qSLC8-1和qSLC8-2在本研究中是效应值最大的2个控制角果长度QTL, 其峰值物理距离在30 Mb和38 Mb附近, 因此qSLC8-1很可能与Zhou等[20]提出的位于C8连锁群约27.4 Mb的共性QTL一致。定位在A2上的角果长度QTL鲜有报道, 但在本研究中qSLA2在多个环境中效应极显著, 加性主效达到0.18 cm, 贡献率大于10%。其他QTL由于所用标记各异, 较难和前人结果比较。
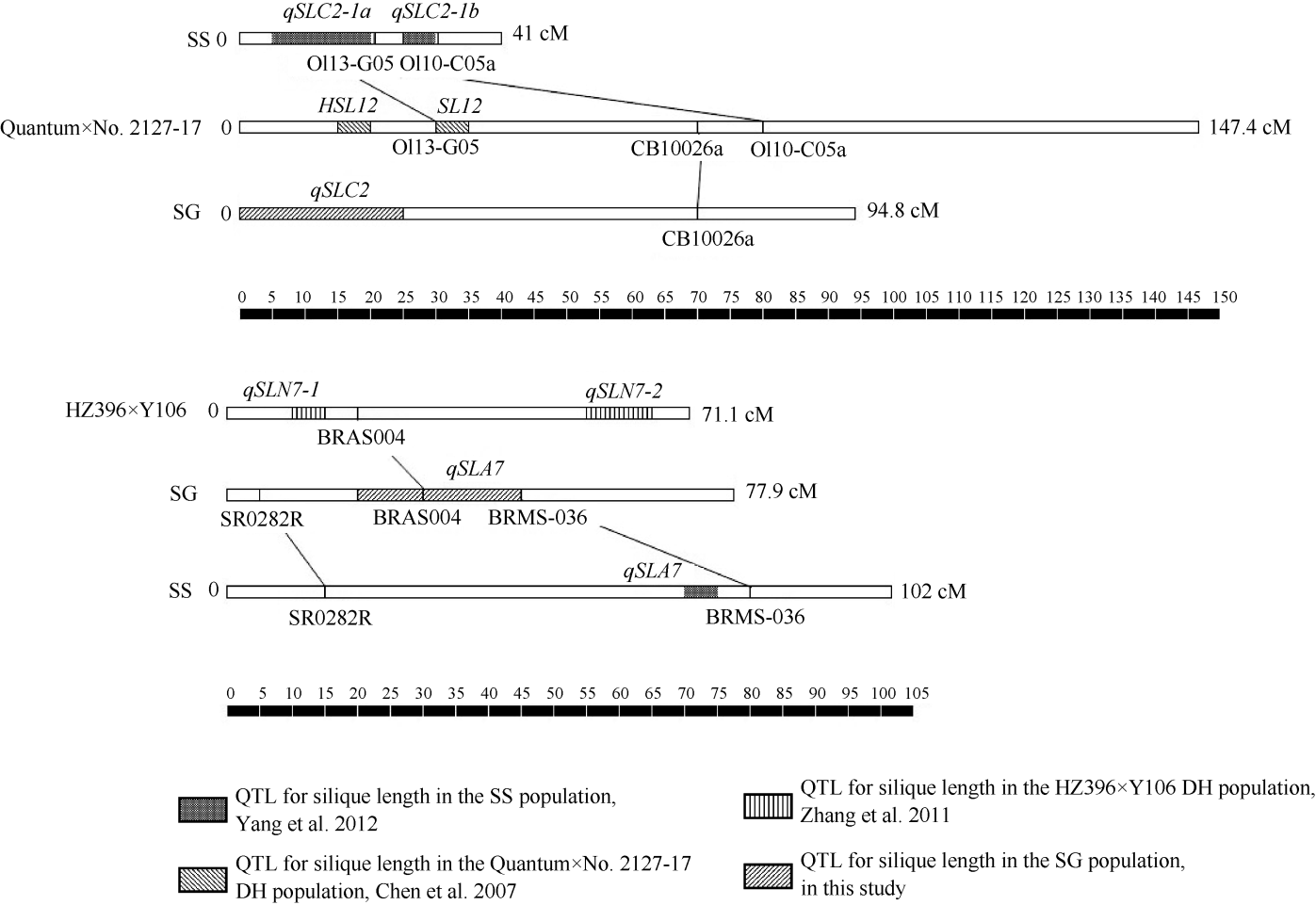 | 图2 SG群体2个角果长度QTL在连锁群上的位置以及与前人发表的研究结果比较Fig. 2 Genomic position of two major QTLs for silique length in SG-DH population and their alignment with reference QTLs |
9个环境下检出的8个角果长度QTL, 除qSLA7和qSLC2分别在2个和3个环境中被检测到微效QE互作外, 其余QTL与环境的互作效应均不显著, 说明角果长度是一个相对稳定的性状, 这与该性状具有较高遗传力相一致[9], 也与前人的研究结果相吻合[19]。上位性分析检出8对互作QTL, 每对效应均较微小, 累计只达加性主效的约1/3。但C2和C8上的2个主效QTL参与上位互作, 因此, 标记辅助时要避开可能产生的不利互作而影响选择效果。
比较角果长度在非条件和分别给定角果粒数和千粒重不变为条件时QTL定位结果, 可以看出角果长度与角果粒数和千粒重遗传相关出现不同作用模式。第一种, 当角果粒数和千粒重被条件后, QTL效应值发生较大改变, 如qSLA2、qSLC1-2和qSLC8-1位点, 角果长度的变化对角果粒数影响较大; 而通过选择qSLA7、qSLC1-2、qSLC8-1和qSLC8-2长角标记基因型, 可同时提高千粒重。这些QTL的效应部分受条件性状所左右, 同时作用于角果长度和条件性状, 为一因多效QTL。这一遗传特点在一定程度上解释了角果长度和角果粒数以及千粒重之间的显著正向相关现象, 其QTL区间可能同时存在控制角果粒数和千粒重的基因。第二种情况, QTL在条件前后效应值变化不大, 方向相同, 如qSLA4、qSLA7和qSLC2基本不受角果粒数变化的影响, 但与千粒重有一定关系。第三种情况, 只在条件QTL分析中出现的新位点, 通常效应值比较低, 这是因为当2个性状同时动态并存时, 其效应被条件性状所掩盖而未能检出。这些QTL效应微小, 对性状影响不大, 但从侧面解释了角果长度和千粒重之间复杂遗传关系。
根据SG群体282个株系9个环境下角果长度平均值, 筛选出角果最长和最短的各20个极端株系, 用与5个显著性最高效应值最大QTL连锁的11个高质量标记, 进行标记基因型和表现型的一对一验证(表5)。与qSLC1-1和qSLC2连锁的各2个标记(ZAASRF27、ZAAS637、ZAAS1004和CN32a)虽然在长角果组(> 6.31 cm)中符合率达95%~100% (95%以上株系含长果等位基因), 但在低角果组中(< 4.54 cm), 符合率不够理想, 20个株系中只有6~11个携短角果等位基因。2个qSLA7连锁标记在2个组中的符合率处于中上水平, 标记ZAAS12a的符合率为65%~75%, 可考虑育种应用。其余3个QTL的5个标记, 与qSLA2连锁的2个标记(ZAAS423和SUC1-3)在长角果组里的准确率为75%, 短角果组中的符合率达90%以上; qSLC8-1和qSLC8-2峰值位置相近处的3个连锁标记(ZAAS433、ZAAS437和ZAASA7-28)在长角和短角果2个组中的标记检测符合率均在85%以上, 长角果组为85%~90%, 短角果组中是95%~ 100%。说明这些标记在油菜育种中作为辅助手段用于长角果材料筛选可望获得预期结果。另值得注意的是我们发现增加角果长度不但使角果粒数增多, 千粒重也显著提高。通过对4个控制角果长度QTLqSLA2、qSLA7、qSLC8-1和qSLC8-2的6个连锁标记的筛选和聚合长角果等位基因, 可间接增加角果粒数同时提高粒重, 从根本上提高油菜产量水平。
共检测到在3个环境以上稳定表达的控制角果长度QTL 8个, 加性效应值在0.09~0.26 cm之间, 效应总和解释群体遗传总变异的60%。8对上位性QTL效应值在0.035~0.075 cm之间, 效应总和为加性总效应的38%。QTL与环境互作效应只在少数位点和个别环境中显著。在qSLA2、qSLC1-2和qSLC8-1位点, 角果长度的变化对角果粒数影响较大; 而通过选择qSLA7、qSLC1-2、qSLC8-1和qSLC8-2长角果标记基因型, 可望同时提高角果粒数和千粒重。通过对qSLA2、qSLA7、qSLC8-1和qSLC8-24个QTL位点的6个连锁标记(ZAAS423、SUC1-3、ZAAS12a、ZAASA7-28、ZAAS433和ZAAS437)长角果基因型的聚合, 可增长角果约2 cm, 间接增加角果粒数2粒, 同时提高千粒重0.4 g, 从而可望实质性地提高油菜产量水平。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|