{kind=link}
小麦株高相关性状与SNP标记全基因组关联分析
[陈广凤1, 2  , 陈建省
, 陈建省1 , 田纪春1, * ]
, 陈建省]
|
|


第一作者联系方式: E-mail: wywdz@sina.cn
株高是影响小麦产量和控制倒伏的重要因素, 研究小麦株高相关性状的遗传机制对高产育种具有指导意义。以205份中国冬麦区小麦品种(系)为材料, 利用分布于小麦全基因组的24 355个单核苷酸多态性(SNP)标记对株高相关性状进行关联分析。共发现38个与株高相关性状显著关联( P < 0.0001)的SNP, 分布在1B、2A、2B、3A、3B、3D、4A、4B、5A和6D染色体上。其中, 11个位点至少在2个环境中稳定表达, 可用于开发CAPS标记。同时, 发掘了一批株高性状相关基因的优异等位变异, 如降低株高的等位变异BobWhite_c48009_52, 平均降低株高12.9 cm; 控制穗下节间长的等位变异BS00039422_51-C和IAAV1698-A, 分别调控穗下节间长5.9 cm和6.6 cm。本研究发掘的控制小麦株高基因位点为在分子水平上研究小麦株高复杂性状提供了有价值的参考。
Plant height (PH) has great influence on yield potential and lodging occurrence in wheat ( Triticum aestivumL.). In this study, the diverse panel of 205 elite wheat lines was genotyped with the 90k SNP array (24 355 SNPs) based on the Illumina Infinium assay to disclose the PH genetic mechanism. A total of 38 SNPs were found to be associated with PH ( P < 0.0001), which were located on chromosomes 1B, 2A, 2B, 3A, 3B, 3D, 4A, 4B, 5A, and 6D, including 11 SNPs detected in two or more environments. These stable SNPs can be used to develop CAPS markers for PH. In which, a few elite alleles were identified, such as BobWhite_c48009_52 to reduce PH by 12.9 cm, and BS00039422_51-C and IAAV1698-A to decrease length of the first internode below spike by 5.9 cm and 6.6 cm, respectively. These results will facilitate further researches in PH-related traits in wheat.
小麦株高不仅影响产量潜力, 而且与抗倒伏性密切相关, 调控株高以提高抗倒伏性一直是小麦重要的育种目标。株高由数个节间长度构成, 不同节间在发育的不同时期伸长, 共同影响株高性状。研究表明, 小麦穗下节长度和倒二节长度与倒伏性密切相关[1, 2]。迄今为止, 已有许多国内外学者在不同遗传背景、不同环境条件下对小麦株高的遗传机制进行了研究, 定位的株高QTL几乎遍布小麦的全部21条染色体。Cadalen等[3]利用DH群体定位了9个与小麦株高相关的QTL, 其中的2个RFLP标记Xfba1-4B和Xfba211-4D分别与矮秆基因Rht1和Rht2连锁; 周淼平等[4]利用一套重组自交系群体检测到4个控制小麦株高的QTL, 位于1D、2B、4A和4D染色体上, 单个QTL对表型变异的解释率为10.3%~33.8%; 刘宾等[5]构建了花培3号和豫麦57杂种F1的DH群体, 利用动态QTL定位方法, 检测
到控制株高的18个非条件QTL和10个条件QTL。这些研究多为基于双亲本遗传群体连锁作图的传统QTL定位, 仅涉及2个特定亲本, 同时在构建分离群体时发生重组的次数有限, 因而鉴定的QTL的置信区间大、分辨率低, 易发生目标基因的丢失或连锁累赘, 难以在分子标记辅助选择(marker- assisted selection, MAS)中加以利用[6]。
近年来, 利用遗传多样性丰富的自然群体进行全基因组关联分析(genome-wide association study, GWAS)成为热点。GWAS直接利用基因本身或基因附近微小区域的分子标记与性状表型的关联来实现基因的精细定位[7, 8]。单核苷多态性(SNP)标记作为目前最具发展潜力的分子标记, 在基因组中具有遗传稳定、数量多、分布广且易于检测等特点, 适合于数量庞大的检测分析, 满足全基因组关联分析对于大样本、高密度标记的要求, 可以大大提高关联分析的统计效力[9, 10]。基于SNP标记的全基因组关联分析所定位的QTL, 解析率高, 在MAS中可提高选择的目的性和准确性, 进而提高育种效率[11], 已成功应用于人类[12]、果蝇[13]、水稻[14]、玉米[15]等多种生物的遗传研究。小麦是异源六倍体, 基因组庞
大, 与水稻、玉米等作物相比, 小麦SNP标记的开发与利用较晚, 基于SNP标记的小麦株高的全基因组关联分析的研究未见报道。本研究以中国冬麦区主要小麦品种(系)为材料, 利用SNP标记进行株高相关性状/标记的关联分析, 以求找到与株高相关性状紧密连锁的SNP标记, 在小麦育种中为聚合和利用控制株高的有利基因/QTL提供参考。
205份供试材料包括中国冬麦区20世纪80年代以来的推广品种或骨干亲本132个, 高代品系73份, 其中高代品系全部来自中国山东省(表1)。
2012— 2013和2013— 2014年度, 分别将供试材料种植于山东农业大学(山东泰安)和德州市农业科学院(山东德州), 每份材料(小区)种3行, 2次重复, 行长2 m, 均匀播种70粒, 行间距25 cm。常规田间管理, 生长期间没有发生严重病虫害和倒伏。灌浆后期按每品种取10株在田间调查株高、穗下节间长以及倒二、倒三和倒四节间长。采用SPSS 17.0软件统计分析数据, 计算相关系数。利用SAS 8.0软件对5个株高相关性状做方差分析, 获得遗传方差(σ g2)和环境方差(σ e2), 计算广义遗传力(hB2)。hB2=σ g2/(σ g2+σ e2)[16]。
| 表1 供试小麦材料 Table 1 Wheat varieties used in this study |
参照略有改动的Triticarte Pty. Ltd (http://www.triticarte.com.au/)方法提取DNA, 用0.8%琼脂糖电泳检测DNA质量和浓度。委托美国加利弗尼亚大学戴维斯分校植物科学系生物技术检测中心, 使用美国Illumina公司和美国堪萨斯州立大学共同开发的小麦90k基因芯片(81 587个SNP)进行群体DNA的基因分型, 利用GenomeStudio软件读取分型数据, 并以文本文件形式导出保存。用PLINK v1.07[17]对获得的基因型数据进行质量控制, 剔除检出率小于80%和低频基因频率(minor allele frequency, MAF)小于5%的SNP标记, 最终获得24 355个SNP用于株高相关性状关联分析。
利用Wang等[18]对6个DH遗传群体(BT-Schomburgk × AUS33384、Young × AUS33414、Chara × Glenlea、W7984 × Opata M85、Sundor × AUS30604和Westonia × Kauz)进行图谱整合的位点信息, 获得本研究群体SNP位点遗传信息(表2)及整合复合遗传图谱。
| 表2 SNP复合遗传图谱信息 Table 2 Information of SNP in the integrated linkage map |
应用PowerMarker V3.25软件[19]计算每个SNP位点的等位变异数、等位基因频率、多态性信息量(PIC)。, pij表示位点i的第j个等位变异出现的频率。
选择在小麦的21条染色体长臂和短臂上均匀分布的3297个SNP标记, 用Structure 2.0软件进行群体结构分析, 采用admixed model, 设K值为2~15, 参数iterations为100 000, burn-in period为100 000。每个K值重复运行5次。以设定的K所对应的最大似然值为目标, 来选择合适的K值作为亚群数目[20]。
应用TASSEL 3.0软件(http://www.maizegenetics.net/)中的MLM (mixed linear model)进行性状和标记之间的关联分析, 利用Structure 2.3.1软件计算Q值, 用TASSEL 3.0软件计算Kinship值。当标记的P≤ 0.0001时认为标记与性状存在关联。
在4个环境中, 5个株高相关性状均表现较大的变异范围, 遗传力以株高最大, 倒四节间长最小(表3)。株高相关性状之间存在不同程度的正相关, 除穗下节间长与倒四节间长不相关以外, 其他性状都呈极显著正相关, 相关系数介于0.061~0.730之间(表4)。
利用24 355个分子标记, 在205份小麦材料中共检测到48 710个等位变异, 每个标记均检测到2个等位变异; 多态性信息含量PIC值介于0.05~0.38之间, 平均0.31, 主要分布于0.35~0.38之间。在全部SNP中, 低频等位基因频率大于0.20的占59.9% (14 597/24 355), 低频等位基因频率接近0.5的占9.6% (2337/24 355)。
群体遗传结构分析表明, 当K=4时, Δ K出现明显峰值(图1-A)。因此, 本试验采用的自然群体应分为4个亚群(图1-B)。第1亚群共43份, 以河南品种为主, 还包括北京、安徽品种及5个山东品系; 第2亚群共32份, 以河北品种为主, 还包括山西、安徽品种及13个山东品系; 第3亚群最大, 共有105份材料, 以山东品种和品系为主, 江苏和甘肃品种以及2个引进品种也分在此亚群; 第4亚群共25份, 包括山西、江苏、宁夏品种及2个山东品系。
| 表3 小麦自然群体株高及相关性状的表型变异 Table 3 Phenotypic variations of plant height and its related traits in the natural wheat population |
| 表4 株高相关性状之间的相关性 Table 4 Pairwise correlation coefficients among plant height related traits |
将SNP标记结果与4个环境下的表型数据进行关联分析, 共检测到38个株高及各节间性状/标记显著关联位点(P< 0.0001), 分布在1B、2A、2B、3A、3B、3D、4A、4B、4D、5A和6D染色体上, 单个关联位点表型变异贡献率(R2)范围为12.79%~24.15% (表5)。
检测到28个与株高显著关联的位点, 分布在1B、2A、2B、3B、3D、4A、4B、4D、5A和6D染色体上。其中, 6D染色体上的2个标记(wsnp_Ra_ c27787_37299352、BobWhite_c14066_403)在4个环境同时被检测到; 3B染色体上的5个标记(wsnp_Ex_ c6065_10623213、wsnp_BE498786B_Ta_2_1、BobWhite _c48009_52、Excalibur_c48368_217、Excalibur_c11001_ 134)和5A上的1个标记(Excalibur_c1208_72)在2个环境同时被检测到。单个关联位点表型变异贡献率为12.79%~23.82%, 贡献率最大的是BS00039422_51, 但仅在环境E2中被检测到。
与穗下节间长显著关联的位点有10个, 分布在2A、3A、3B和6D染色体上, 其中BS00039422_51和IAAV1698在3个环境中同时被检测到。单个关联位点表型变异贡献率为15.57%~20.86%, Ra_ c106454_621的表型贡献率最大, 只在环境E4中被检测到。
检测到10个与倒二节间长显著关联的位点, 分布在2A、2B、3B和4A染色体上。其中, RFL_ Contig5495_464在2个环境中同时被检测到, 对表型变异贡献率分别为17.93%和14.36%。单个关联位点表型变异贡献率为14.36%~24.15%, BS00039422_ 51虽然贡献率最大, 但仅在环境E2中被检测到, 且其P值最小。
与倒三和倒四节间长显著关联的位点较少, 其中倒三节间长的关联位点只有E3环境下的1个, 位于4B染色体, 能解释15.79%的表型变异; 倒四节间长的关联位点有2个, 在环境E1和E4被检测到, 分别分布在3A和3B染色体, 对表型变异的贡献率为15.20%和17.50%。
在多个环境下均能检测到的关联位点为稳定位点, 本研究发现控制株高的稳定位点最多, 共有8
个, 此外分别有2个和1个位点控制穗下节间长和倒二节间长。在4个环境中同时检测到的wsnp_Ra_ c27787_37299352和BobWhite_c14066_403, 都位于6D上, 与株高关联; 2A上的BS00039422_51和IAAV1698与穗下节间长关联, 在3个环境中同时被检测到; 共有7个关联标记在2个环境中被检测到, 其中6个控制株高, 包括3B上的5个(wsnp_Ex_ c6065_1062321、wsnp_BE498786B_Ta_2_1、BobWhite_ c48009_52、Excalibur_c48368_217和Excalibur_ c11001_134)和5A上的1个(Excalibur_c1208_72), 还有1个位点是2B上的RFL_Contig5495_464, 控制倒二节间长。
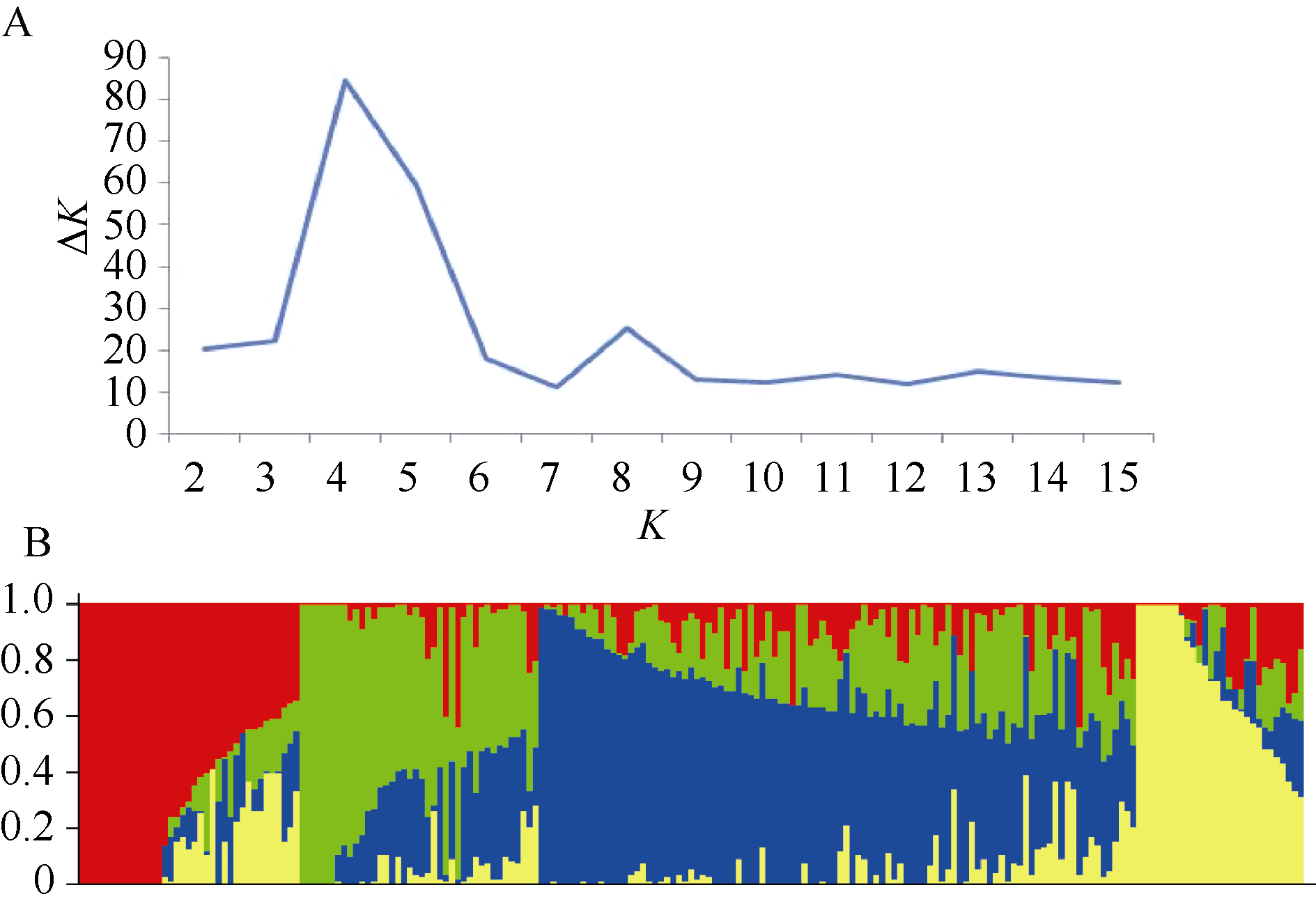 | 图1 205份小麦材料的群体结构Fig. 1 Population structure of 205 wheat accessions |
| 表5 株高性状关联位点及其对表型变异的贡献率(R2) Table 5 Loci associated with plant height-related traits and percentage of phenotypic variation explained (R2) |
| (续表5) 株高性状关联位点及其对表型变异的贡献率(R2) Table 5(Continued) Loci associated with plant height-related traits and percentage of phenotypic variation explained (R2) |
在这些稳定表达的关联标记中, 存在一因多效现象, 如2A上的BS00039422_51和IAAV1698, 同时与株高、穗下节间长和倒二节间长相关联; 6D上的wsnp_Ra_c27787_37299352和BobWhite_c14066_ 403, 同时与株高和穗下节间长关联。这一现象与相关分析结果相吻合。
某些位点的优势等位变异就是优异等位变异[21]。与株高关联的稳定位点wsnp_Ex_c6065_10623213, 其优势等位变异为碱基C, 携带该等位变异的材料在4个环境中的株高及均值都极显著低于携带碱基A的材料; BobWhite_c48009_52对株高的增效作用最大, 该位点的碱基G相对于A为优异等位变异, 携带该优异等位变异的材料平均降低株高12.9 cm; Excalibur_c1208_72对株高的增效作用最小, 携带其优异等位变异C的材料较携带A的材料平均降低株高5.9 cm。此外, 与穗下节间长关联的优异等位变异为BS00039422_51-C和IAAV1698-A, 分别降低穗下节间长5.9 cm和6.6 cm。与倒二节间长关联的优异等位变异为RFL_Contig5495_464-C, 平均降低倒二节间长3.8 cm (表6)。
| 表6 相对稳定的关联位点等位变异的表型效应 Table 6 Phenotypic effect of alleles for the relatively stable loci |
全基因组关联分析(GWAS)是以自然群体为研究对象, 以长期重组后保留下来的基因(位点)间连锁不平衡为基础, 利用分布于全基因组的SNP将目标性状表型的多样性与基因(标记)的多态性结合起来分析, 可直接鉴定出与表型变异密切相关且具有特定功能的SNP或基因位点。目前, GWAS已应用于水稻[14]、玉米[15]等农作物及模式植物拟南芥[22]主要性状位点的鉴定与挖掘。由于小麦全基因组精密物理图谱尚在构建之中, 因此小麦上利用与遗传作图群体共有标记的遗传图谱信息, 整合构建自然群体标记相应的遗传图谱, 为进行GWAS提供标记位点的SNP水平的遗传信息, 具有重要应用价值。Zanke等[23]利用358个欧洲冬麦品种组成的自然群体, 构建了包括732个SSR标和7934个SNP标记的遗传图谱, 定位到控制小麦抽穗期的72个SSR和432个SNP位点, 并通过和水稻、两穗短柄草同源序列的比对, 发现在水稻上控制光周期的基因Hd6和小麦位于5BL上控制抽穗期的显著位点具有高度同源性。Zhang等[24]利用在中国黄淮麦区收集的96个小麦材料, 构建了包括443个SSR和738个DArT标记的遗传图谱, 在不同的水肥条件下定位到37个控制小麦粒重的显著关联位点。本研究根据6个已知遗传群体SNP标记的位点信息, 整合了自然群体24 355个SNP标记相应的复合遗传图谱, 为株高相关性状GWAS显著关联位点提供遗传信息, 下一步可以利用这些信息对关联SNP位点序列进行同源比对、基因注释等生物信息学分析, 为小麦株高候选基因的筛选和预测提供重要参考。
目前, 已命名了25个与株高相关的Rht (reduced height)基因。其中, Rht1(Rht-B1b)、Rht2 (Rht- D1b)、Rht8、Rht9和Rht10等12个基因被定位于4B、4D、2D、4D和7B等染色体上, 另外13个矮秆基因尚未定位[25, 26]。本文通过GWAS得到的38个株高/标记的显著SNP位点, 分布在1B、2A、2B、3A、3B、3D、4A、4B、4D、5A和6D染色体上。在4B染色体检测到的标记Ra_c2142_821 (114.87 cM), 同时与株高和倒三节间长关联, 遗传贡献率较大。刘宾等[5]也在4B染色体检测到控制株高的Qph4B在整个生育期频繁表达; Bö rner等[27]在相似的位置发现一个QTL, 并确定为矮秆基因Rht1的位点。在4D染色体上发现的控制株高的标记Kukri_rep_c68594_530与Zhang等[28]定位的株高QTL位置相似, Cadalen等[3]、Huang等[29]和王竹林等[30]也证实Rht-D1b位于4D染色体的相邻区间, 可能为矮秆基因Rht2。2A染色体上的标记BS00039422_51和IAAV1698遗传距离相近, 同时控制株高、穗下节间长和倒二节间长, 且遗传贡献率较大。魏添梅等[31]、张国华等[32]也在相似位置发现了控制株高的QTL。此外, 本研究还检测到一些新位点, 如3B染色体上与株高和倒二节间长关联的wsnp_BE498786B_Ta_2_1和BobWhite_c48009_52, 是否是未被定位的株高基因有待于进一步证实。
本研究利用全基因组SNP标记对小麦株高相关性状进行GWAS, 采用混合线性模型(MLM), 将群体结构分析所得各个体的Q值和亲缘关系作为协变量纳入回归分析, 并采用较高的阈值(P≤ 0.0001), 可有效消除由群体分层和亲缘关系引起的伪关联, 增加关联结果的可信度。不同环境下检测出的位点存在不一致的现象, 可能是环境影响的结果, 这是多基因控制的数量性状的突出特点。有些关联的位点在多个环境均能被检测到, 这些位点可视为相对稳定的位点。SNP标记在小麦基因组中广泛存在, 遗传稳定, 部分基因内部的SNP直接影响蛋白质的结构或基因表达的水平, 本身可能是候选基因的功能位点, 因此它们比SSR与功能基因的联系更为紧密[33]。本研究通过关联分析筛选到11个与株高相关性状稳定关联的SNP位点, 如4个环境下同时检测到的与株高显著关联的标记wsnp_Ra_c27787_37299352和BobWhite_c14066_403, 3个环境下同时检测到的与穗下节间长显著关联的标记BS00039422_51和IAAV1698等, 可用于开发CAPS标记, 应用于小麦株高分子标记辅助育种, 并通过对SNP位点序列的同源比对、基因注释等生物信息学分析, 有效发掘株高相关基因。
利用关联分析发掘作物的优异基因是目前植物基因组学研究的重要方法之一。本研究解析了与小麦株高稳定关联位点内等位变异的表型效应, 在性状的表型与基因型之间用特定的等位变异建立起联系, 发掘了与株高关联的优异等位变异, 如降低株高的等位变异BobWhite_c48009_52, 降低穗下节间长的等位变异BS00039422_51-C。在小麦育种工作中, 可以利用携带这些优异等位变异的材料做亲本, 并借助分子标记辅助选择将多个目标基因导入或聚合, 提高育种选择效率。
根据来自6个DH群体的遗传图谱信息, 构建了本研究群体SNP标记的复合遗传图谱。利用24 355个SNP标记对205份小麦品种(系)进行株高相关性状全基因组关联分析, 检测到38个与株高相关性状显著关联(P< 0.0001)的SNP, 其中稳定位点11个, 可用于开发株高CAPS标记。部分稳定位点的优异等位变异有较好的降低株高(BobWhite_c48009_52)和穗下节间长(BS00039422_51-C和IAAV1698-A)的作用, 可用于小麦株高改良育种。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|