

{kind=link}
{kind=link}
{kind=link}
基于高密度遗传图谱的玉米籽粒性状QTL定位
[秦伟伟 , 李永祥, 李春辉, 陈林, 吴迅, 白娜, 石云素, 宋燕春, 张登峰, 王天宇
, 李永祥, 李春辉, 陈林, 吴迅, 白娜, 石云素, 宋燕春, 张登峰, 王天宇* , 黎裕* ]
, 李永祥, 李春辉, 陈林, 吴迅, 白娜, 石云素, 宋燕春, 张登峰, 王天宇, 黎裕]
|
|


第一作者联系方式: E-mail: 13811531756@139.com
籽粒大小及百粒重是决定玉米产量的重要因素。为解析籽粒性状遗传基础, 本研究以玉米自交系黄早四(HZS)和Mo17为亲本, 构建包含130个重组自交系(recombination inbred line, RIL)的RIL群体。基于GBS (genotyping- by-sequencing)技术获得的高密度多态性SNP (single nucleotide polymorphism)位点, 构建了包含1262个Bin标记的高密度遗传图谱。采用完备区间作图法, 对5个环境条件下的粒长、粒宽、百粒重、粒长/粒宽4个性状分别进行QTL (quantitative trait locus)定位, 共检测到30个QTL。利用5个环境性状均值, 共检测到11个QTL。其中粒长主效QTL qklen1、粒长/粒宽主效QTL qklw1在3个单环境条件下均被检测到, 且定位在第1染色体相邻区域, 物理位置分别为210~212 Mb、207~208 Mb, 表型贡献率分别为22.60%和26.79%, 被认为是控制玉米籽粒形状的主效位点。针对第1染色体207~212 Mb区间, 采用成组法 t检验, 对黄早四(受体)和Mo17 (供体)构建的BC3F1回交群体进行单标记分析。结果表明, 在BC3F1群体中 qklen1和 qklw1同样具有显著的遗传效应。本研究结果不仅为分子标记辅助选择籽粒性状提供了实用标记, 而且为主效基因的进一步精细定位和候选基因挖掘奠定了基础。
Kernel size and weight are major determinants of grain yield. For understanding molecular mechanisms of kernel related traits, a recombinant inbred line (RIL) mapping population including 130 families was developed from the cross of two maize elite inbreds, Huangzaosi (HZS) and Mo17. By using the approach of GBS (genotyping-by-sequencing), the high-density genetic map with 1262 bin markers was constructed. QTLs for kernel related traits were identified by stepwise regression (RSTEP-LRT) using Windows QTL ICI-Mapping software under five environments. In total, 30 QTLs were detected under single environment and 11 QTLs were detected under the joint environment. The kernel length major QTL qklen1and the length/width major QTL qklw1 were found in the adjoining regions with a strong genetic effect under three environments. QTL qklen1was located in a region of 210-212 Mb on chromosome 1 with explained 22.60% of phenotypic variance, and qklw1 was located in a region of 207-208 Mb on chromosome 1 with explained 26.79% of phenotypic variance. In addition, for further verification, another introgression population of BC3F1 was developed with Mo17 as the donor parent and HZS as the recurrent parent. The result of the single marker analysis suggested that qklen1 and qklw1 also had a significantly genetic effect, which is similar to that in the RIL population. The present study provides a good basis for studying genetic mechanism and molecular marker assisted selection for the improvement of kernel related traits in maize.
玉米作为重要粮、经、饲兼用作物, 据预测全球需求量到2050年将增加一倍[1]。因此, 高产仍是玉米育种的主要目标。玉米产量性状是最复杂的性状之一[2]。将产量性状分解为若干构成因子, 不仅可提高产量相关基因挖掘效率[3], 而且有利于阐述影响产量的遗传基础[4, 5]。玉米的产量构成因子可分为百粒重、穗粒数、单位面积有效穗数。其中, 百粒重具有较高的遗传力, 是玉米育种过程中的重要选择性状。百粒重主要取决于籽粒的大小和形状[6]。在作物的选择驯化过程中, 粒形和粒重也被认为是产量的重要构成因子[7]。此外, 玉米籽粒性状(粒重、粒长、粒宽、粒厚)与产量均表现出较高的相关性[8], 且相对于产量而言, 具有受环境影响小的特点[9]。因此, 将籽粒相关性状作为研究目标是解析玉米产量遗传基础的重要途径。
玉米基因组学的迅速发展[10]和新型标记的开发, 推动了玉米籽粒相关数量性状位点(QTL)的挖掘进程。如Liu等[11]利用F2:3群体及256个SSR标记定位到28个籽粒相关QTL; Zhang等[12]利用永久性F2群体及217个SSR标记定位到54个籽粒相关QTL; Nikolic等[13]利用F3群体及234个分子标记定位到18个籽粒相关QTL。但是, 往往由于构建遗传连锁图谱时用到的标记数量较少, 使遗传图谱分辨率较低, 定位到的QTL置信区间大都在10~20 cM之间, 导致其结果难以用于分子标记辅助育种, 也限制了玉米产量遗传基础的深度解析。因此, 有必要利用高密度分子标记构建遗传连锁图谱, 提高籽粒相关性状QTL定位的精确度。
近年来, 随着单核苷酸多态性(SNP)标记的开发成本和使用成本大幅降低, 利用SNP标记鉴定遗传作图群体中个体的基因型并构建高密度遗传图谱得以实现。如Colasuonno等[14]利用5019个SNP标记构建的高密度遗传连锁图谱对硬粒小麦黄色素含量进行QTL定位, 标记间平均距离为0.36 cM。Guo等[15]利用玉米永久性F2群体整合18 840个SNP标记, 构建了含3184个区块(Bin)的高密度遗传连锁图谱, 标记间平均距离缩小到0.84 cM, 并对杂种优势进行遗传解析。Zou等[16]利用高粱RIL群体, 通过构建的含3418个Bin的高密度遗传连锁图谱, 标记间平均距离为0.50 cM, 对花期和株型相关性状进行了QTL定位。这些研究在很大程度上缩小了QTL置信区间, 对开发紧密连锁的分子标记和下一步的QTL精细定位提供了更多的遗传信息。本研究在不同环境条件下对玉米粒长、粒宽、百粒重和粒长/粒宽进行了表型鉴定, 并借助GBS (genotyping-by- sequencing)技术获得的SNP标记构建高密度遗传图谱, 进行玉米籽粒性状的QTL定位, 旨在利用高密度遗传图谱检测玉米籽粒性状QTL, 从而为分子标记辅助育种和玉米产量相关性状的主效QTL精细定位及候选基因挖掘提供新的信息和科学依据。
以玉米自交系黄早四和Mo17为亲本配制组合, 从F2代开始采用“ 单粒传法” 连续自交至F8代, 构建成一套包含130个重组自交系(recombination inbred line, RIL)群体。另外, 以玉米自交系黄早四为受体亲本, 以Mo17为供体亲本, 从BC1开始每个世代取混合花粉与黄早四连续回交3代, 获得含152个单株的BC3F1高代回交群体。
对RIL群体2009年在北京、新疆、河南及2010年在新疆、河南共5个环境条件下进行表型鉴定。在每个环境下均采用随机区组设计, 单行区, 2次重复, 行长3.0 m, 每行11株, 行距0.6 m。植株成熟后, 从每个小区收获5株果穗用于籽粒相关性状的考察。所考察的性状包括粒长、粒宽和百粒重。粒长和粒宽为每个果穗中间部分连续籽粒的10籽长和10粒宽, 单位为cm。百粒重为每个小区果穗脱粒后, 3次随机取样100粒的平均值, 单位为g。粒长/粒宽为二级性状, 即粒长与粒宽的比值。
2012年在北京对BC3F1群体进行表型鉴定。对所有材料均采用单粒播种, 行长3.0 m, 每行定苗11株, 行距0.6 m, 株距25 cm。玉米拔节期单株编号挂牌, 待植株完全成熟后, 按编号单株收获。取果穗中部籽粒, 测量粒长、粒宽和百粒重, 每个果穗重复测量3次。
全部田间试验中的施肥、灌溉、防虫和除草同当地大田管理一致。
对RIL群体, 在植株长到六叶时混合取每个家系的幼嫩叶片, 采用CTAB法[17]提取基因组DNA。利用简化基因组测序(genotyping-by-sequencing, GBS)技术[18]鉴定家系基因型。
对BC3F1群体, 在玉米拔节期取亲本和回交群体单株新鲜叶片, 利用CTAB法提取DNA[17]。参考中国农业大学赖锦盛教授课题组提供的黄早四和Mo17重测序结果, 针对RIL群体中定位到的QTL qklen1和qklw1的基因组区段, 利用Primer5.0设计开发InDel标记。通过PCR产物扩增, 8.0%非变性聚丙烯酰胺凝胶电泳, 0.1% AgNO3染色, 甲醛和NaOH显色, 进行BC3F1群体基因型鉴定。
黄早四籽粒属于硬粒型, 籽粒宽、饱满; 而Mo17籽粒属于马齿型, 籽粒长、扁平, 在5种不同环境下亲本间籽粒性状的表型具有明显的差异(表1)。因此, 利用这两个自交系构建的群体后代存在较丰富的表型变异(表2)。例如, 在RIL群体中, 家系粒长均值为9.03 cm, 变异范围为7.67~10.26 cm。粒长、粒宽、粒长/粒宽、百粒重等4个籽粒相关性状的表型频率均呈正态分布或近似正态分布(图1), 可作为数量性状进行QTL作图。此外, 对这4个籽粒相关性状多环境联合方差分析(表3)显示, 粒长、粒宽、粒长/粒宽、百粒重的遗传力分别为0.861、0.860、0.869、0.855。
| 表1 5个不同环境下亲本间籽粒性状的表型差异 Table 1 Difference of the kernel related traits between parents under five environments |
| 表2 RIL群体和BC3F1群体中籽粒性状的表现 Table 2 Performance of kernel related traits in RIL and BC3F1 populations |
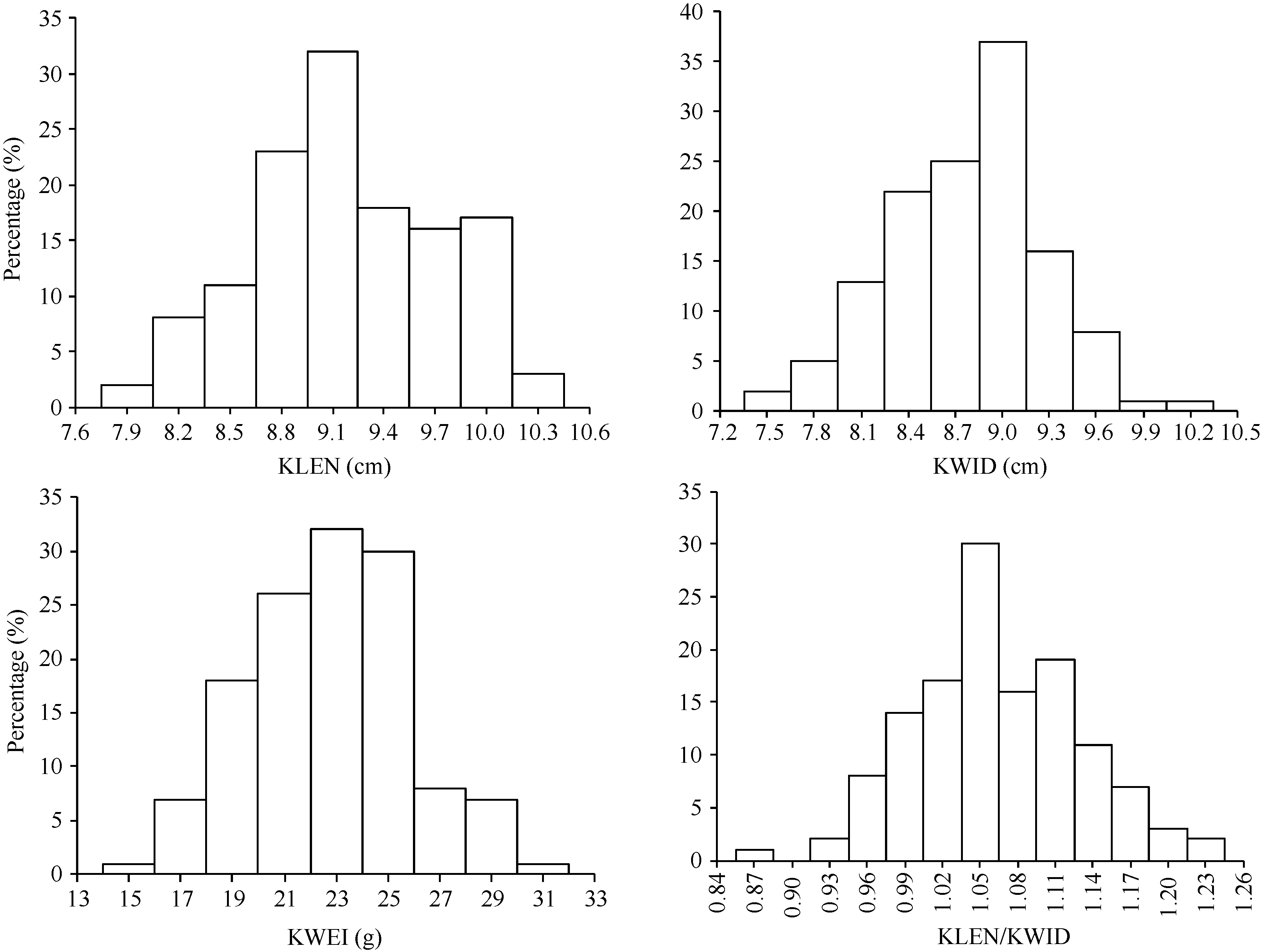 | 图1 RIL群体中粒长、粒宽、百粒重和粒长/粒宽表型频率分布Fig. 1 Frequency distributions of the KLEN, KWID, KWEI, KLEN/WEID in the RIL populationKLEN: 10-kernel length; KWID: 10-kernel width; KWEI: 100-kernel weight; KLEN/KWID: kernel length/kernel width. |
| 表3 籽粒相关性状在5个环境下的联合方差分析 Table 3 Analysis of variance (ANOVA) for kernel-related traits under five environments |
利用GBS技术, 检测RIL群体的基因型, 共得到78 507个高质量SNP。基于这些SNP, 构建RIL群体的重组区块图谱共得到1262个Bin标记。利用这些Bin构建的遗传连锁图谱总长度为1524.53 cM, 标记间最大距离为24.14 cM, 最小距离0.38 cM, 标记间的平均距离为1.21 cM。
在单环境作图中, 共检测到4个籽粒相关性状30个QTL, 分布在第1、第2、第4、第6、第7、第8、第10染色体上, 单个QTL的表型贡献率变化范围在6.28%~27.63%之间(表4)。利用多环境平均值进行QTL定位时, 共检测到所有性状11个QTL, 分布在第1、第2、第4、第6、第9、第10染色体上, 单个QTL解释的表型变异在6.01%~26.79%之间(表5)。单个环境和多个环境检测到的QTL在遗传图谱上的位置如图2所示。
2.3.1 粒长QTL 在单环境条件下, 共定位到7个粒长QTL, 分布在第1、第4、第7、第10染色体上。利用多环境平均值进行QTL定位时, 在第1染色体物理位置210~212 Mb处检测到一个粒长主效QTL并命名为qklen1, 其LOD值为7.15, 表型贡献率达22.60% (表5)。该位点可在2009北京、2009新疆、2010新疆各单环境下被检测到, 相应表型贡献率分别为23.35%、21.00%、10.63% (表4)。
| 表4 单环境下籽粒相关性状QTL分析 Table 4 QTL analysis of kernel related traits under single environment |
| 表5 多环境条件下籽粒相关性状表型平均值QTL定位分析 Table 5 QTL analysis of kernel related traits under joint-environments using phenotypic means |
2.3.2 粒宽QTL 在单环境条件下, 共定位到6个粒宽QTL, 分布在第6、第10染色体上。其中在第10染色体物理位置147 Mb处存在一粒宽QTL, 在2009北京、2009新疆、2010河南各单环境条件下均能被检测到(表4)。以多环境平均值进行QTL定位时, 在第4、第6、第9和第10染色体上各检测到1个粒宽QTL, 分别命名为qkwid4、qkwid6、qkwid9、qkwid10, 其表型贡献率在6.01%~15.91%之间(表5)。
2.3.3 百粒重QTL 在各个单环境下共检测到7个粒重QTL, 分布在第1、第4、第7、第8、第10染色体上。以多环境平均值进行QTL定位时, 共检测到4个QTL。其中qkwei10表型贡献率为9.31%, LOD值为3.63, 物理位置位于第10染色体147 Mb左右(表5)。同时, 该位点在2009北京、2009新疆2个单环境下均能检测到(表4)。
2.3.4 粒长/粒宽QTL 在各个单环境下共检测到10个粒长/粒宽QTL, 分布在第1、第2、第6、第8染色体上。以多环境平均值进行QTL定位时, 共检测到2个QTL, 分布在第1、第2染色体上。其中, qklw1物理位置在第1染色体207~208 Mb处, 其LOD值为9.20, 表型贡献率可达26.79% (表5)。同时该主效位点在2009北京、2009河南、2010新疆3个单环境下均能检测到(表4)。
RIL群体的籽粒相关性状QTL定位结果表明, qklen1和qklw1不仅是两个环境钝感的主效QTL, 而且其物理位置在第1染色体上相邻(图3)。因此, 这两个QTL被认为是精细定位的重要候选区段。针对该区间, 利用5个InDel标记(表6), 采用成组法t检验, 对BC3F1群体粒长、粒长/粒宽进行单标记分析。其中, 针对粒长的单标记分析结果表明, 主效区段内5个InDel标记对应的P值均达到极显著水平(P< 0.01)。另外, 在粒长/粒宽的单标记分析中, 标记ML162、ML178和ML93对应的P值同样达到极显著水平(表6)。以上表明, 主效位点qklen1和qklw1在BC3F1群体中同样具有显著的遗传效应。
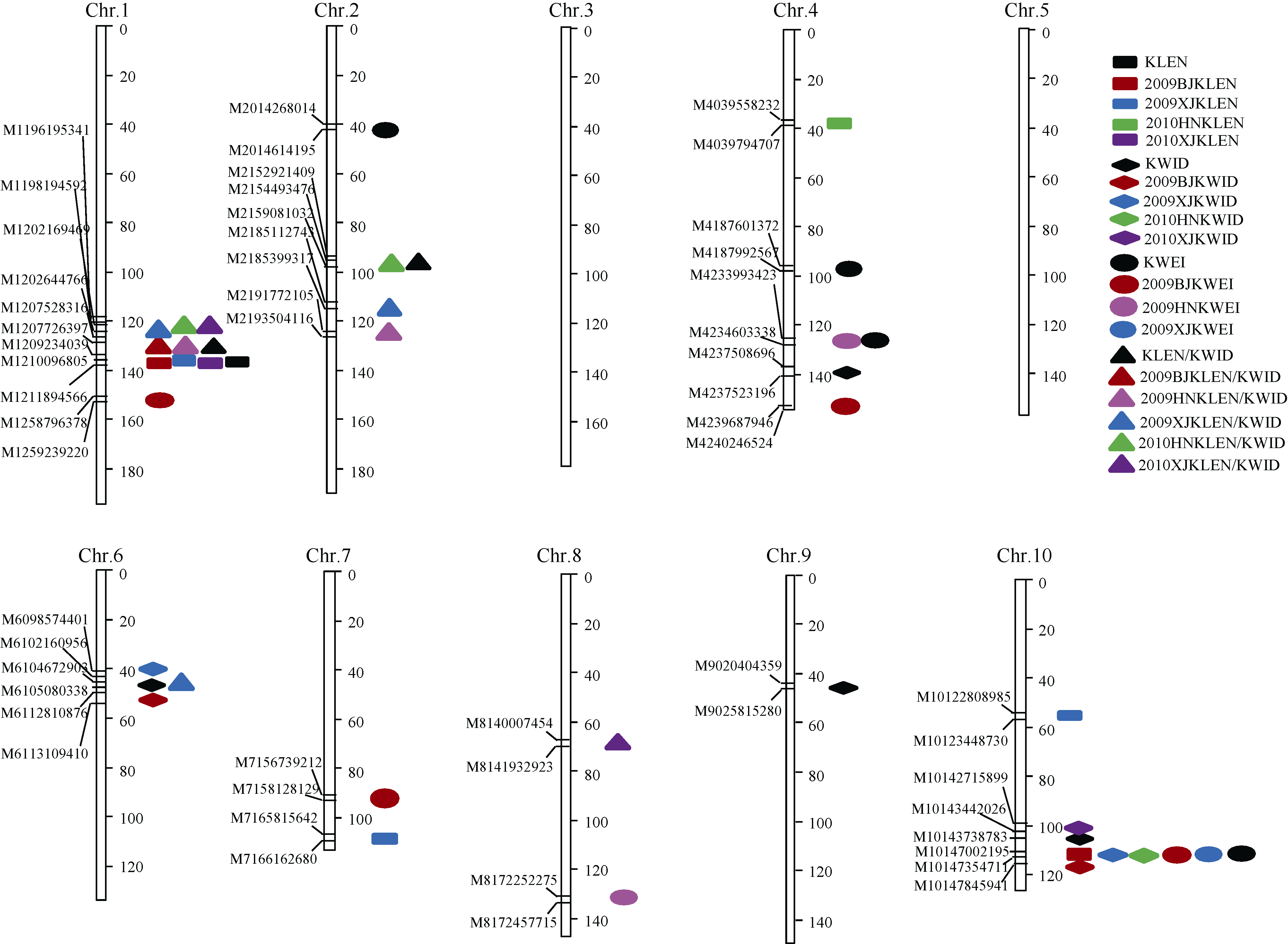 | 图2 籽粒相关性状的QTL在遗传图谱上的分布彩色图案为单个环境监测到的QTL, 黑色图案为多环境联合定位到的QTL; 2009BJKLEN: 2009年北京粒长; 2009XJKLEN: 2009年新疆粒长; 2010HNKLEN: 2010年河南粒长; 2010XJKLEN: 2010年新疆粒长; 2009BJKWID: 2009年北京粒宽; 2009XJKWID: 2009年新疆粒宽; 2010HNKWID: 2010年河南粒宽; 2010XJKWID: 2010年新疆粒宽; 2009BJKWEI: 2009年北京粒重; 2009HNKWEI: 2009年河南粒重; 2009XJKWEI: 2009年新疆粒重; 2009BJKLEN/KWID: 2009年北京粒长/粒宽; 2009HNKLEN/KWID: 2009年河南粒长/粒宽; 2009XJKLEN/KWID: 2009年新疆粒长/粒宽; 2010HNKLEN/KWID: 2010年河南粒长/粒宽; 2010XJKLEN/KWID: 2010年新疆粒长/粒宽。Fig. 2 Distributions of identified QTL for kernel related traits on genetic linkage mapsThe color symbols stand for QTLs detected in a single environment, and the black symbols stand for QTLs detected across five environments; 2009BJKLEN: 10-kernel length in Beijing in 2009; 2009XJKLEN: 10-kernel length in Xinjiang in 2009; 2010HNKLEN: 10-kernel length in Henan in 2010; 2010XJKLEN: 10-kernel length in Xinjiang in 2010; 2009BJKWID: 10-kernel width in Beijing in 2009; 2009XJKWID 10-kernel width in Xinjiang in 2009; 2010HNKWID: 10-kernel width in Henan in 2010; 2010XJKWID: 10-kernel width in Xinjiang in 2010; 2009BJKWEI: 100-kernel weight in Beijing in 2009; 2009HNKWEI: 100-kernel weight in Henan in 2009; 2009XJKWEI: 100-kernel weight in Xinjiang in 2009; 2009BJKLEN/KWID: kernel length/kernel width in Beijing in 2009; 2009HNKLEN/KWID: kernel length/kernel width in Henan in 2009; 2009XJKLEN/KWID: kernel length/kernel width in Xinjiang in 2009; 2010HNKLEN/KWID: kernel length/kernel width in Henan in 2010; 2010XJKLEN/KWID: kernel length/kernel width in Xinjiang in 2010; KLEN: 10-kernel length; KWID: 10-kernel width; KWEI: 100-kernel weight; KLEN/KWID: kernel length/kernel width. |
| 表6 InDel标记引物及相应P值 Table 6 Primers of InDel markers and the corresponding P-value |
 | 图3 粒长和粒长/粒宽相关主效QTL在第1染色体上的分布Fig. 3 Distributions of KLEN and KLEN/KWID related major QTL on chromosome 1KLEN: 10-kernel length; KLEN/KWID: kernel length/kernel width. |
玉米是世界上最重要的粮食作物之一, 高产仍是其育种的首要目标[23]。而籽粒产量性状遗传基础复杂, 是由相互关联的多基因控制的一系列生理生化过程的最终体现[24]。产量性状的复杂性导致直接解析其遗传基础的一定难度。但是, 与产量显著相关的玉米籽粒性状, 例如, 粒长、粒宽、粒长/粒宽和百粒重等, 遗传力较高, 表型评价更加准确, 可作为产量性状的重要构成因子。在此条件下, 加强产量构成因子的遗传基础的解析和功能分子标记开发对于玉米高产育种具有重要意义。
本研究利用亲本黄早四和Mo17构建的RIL群体, 借助高密度遗传图谱, 对多环境下的籽粒相关性状进行QTL定位分析。结果显示, 一些QTL在不同环境下定位在基因组的同一区域, 如粒长主效QTL qklen1、粒长/粒宽QTL qklw1在3个不同环境条件下都被检测到。这说明这些QTL在不同的环境条件下都能稳定地表达, 是进一步精细定位和挖掘主效基因的候选项区段。另外, 不同性状QTL被定位于基因组中同一位点或相近区域, Tuberosa等[25]认为“ 一因多效” 很可能是导致该现象的主要原因; 控制不同性状的基因存在紧密连锁关系也可引起这种富集现象的发生。
通过遗传连锁作图, 本文构建了高密度遗传连锁图谱, 图谱共包含1262个Bin标记, 长度为1524.53 cM, 标记间的平均距离仅为1.21 cM。与前人研究相比, QTL作图精度显著提高。Li等[26]利用179个分子标记对本文中的同一RIL群体的籽粒性状进行QTL分析, 在多环境联合条件定位到的Mqkwei4、Mqklen1、Mqkwid6、Mqkwid10分别和本研究中定位到的qkwei4-2、qklen1、qkwid6、qkwid10位于染色体同一区域。而本研究中定位到的qkwid4、qkwid9、qkwei2、qkwei4-1、qkwei10在Li等[26]研究中并未检测到, 分析其原因, 在低密度遗传图谱中, 较大的区间标记将导致部分重组无法被有效检测, 这将引起表型与基因型关联之间出现偏差, 进而导致一些主效QTL检测的缺失。另外, Li等[26]在RIL群体中定位到的一些QTL置信区间一般为10~30 cM, 而本文检测到的QTL位点置信区间一般为0.8~3.0 cM, 作图精度比低密度图谱提高了10倍左右。例如, 本文中粒长QTL位点qklen1置信区间为1.2 cM, 对应在第1染色体物理位置210~212 Mb处。Li等[26]也在此区域定位到一粒长主效QTL, 但置信区间达到了36.0 cM。进一步显示了高密度遗传图谱在数量性状遗传定位中的巨大优势。
本研究发现, 在第1染色体上临近的2个主效QTL位点, qklen1 (210~212 Mb)和qklw1 (207~208 Mb), 在多环境下都能稳定表达。Li等[26]、Austin等[27]、张向歌等[28]、张伟强等[29]也在相同或邻近该区域内检测到粒长QTL。另外, 在第10染色体定位到的粒宽QTL qkwid10 (143.4~143.7 Mb)和粒重QTLqkwei10 (147.0~147.3 Mb)同样在多环境下稳定表达。Peng等[9]利用F2:3群体在联合环境下将粒宽、粒重QTL定位在第10染色体物理位置143~147 Mb (bnlg1677~umc2172)之间。以上表明, 这两处区域遗传效应稳定, 是控制籽粒性状的重要位点, 可作为精细定位的重要候选区段。利用黄早四和Mo17构建的BC3F1群体对qklen1和qklw1遗传效应验证表明, 在BC3F1群体中qklen1和qklw1同样具有显著的遗传效应。由于BC3F1群体在207~212 Mb区段内发生重组交换的个体较少, 且2个QTL紧密连锁, 所以还不能判断该区段具有“ 一因多效” 的遗传效应, 还是两个独立基因在起作用, 需进一步自交利用大规模分离群体精细定位来验证。
控制不同性状的QTL可定位在相同或邻近的染色体区域; 部分主效位点可在多个环境下被检测到。其中, 粒长主效位点(qklen1)和粒长/粒宽主效位点(qklw1)在3个环境下均能被检测到, 且定位于染色体的相邻位置。qklen1和qklw1在BC3F1群体中同样具有显著的遗传学效应。以上结果为玉米籽粒性状主效QTL的精细定位和图位克隆提供了重要的研究基础。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|